1. Introduction
Contamination of crop plants by human pathogenic bacteria can already occur at plant growth in the field or greenhouse. The transmission routes via which these pathogens are transmitted to crop plants are not always clear, but contamination can occur via (fresh) manure application to soil, upon irrigation, or via human handling with contaminated materials, equipment or machines. Alternative transmission routes of human pathogens to plants are less explored and one of these can be via starting materials such as seeds.
Seeds used for production of sprouts were reported to be contaminated with human pathogens such as
Escherichia coli, especially the enterohemorrhagic variants thereof (EHEC),
Salmonella enterica, Bacillus cereus, Listeria monocytogenes, Staphylococcus aureus and
Yersinia enterolitica [
1]. Human pathogens of different strains of
S. enterica,
E. coli O157:H7 and
E. coli O104:H4 were shown to attach to fenugreek, alfalfa, lettuce and tomato seeds within 5 h upon exposure [
2]. Furthermore, it was shown that
S. enterica and
E. coli O157:H7 strains could persist on butterhead lettuce seeds for two years and after that time, both pathogens were still able to proliferate on germinating seeds [
3]. From these cases it is clear that seeds can become contaminated with human pathogens and that contaminated seeds can pose serious risks for human health at primary production of freshly consumed vegetables and fruits.
The largest outbreak related to consumption of produce was in Hamburg and the surrounding area (Germany) in 2011 upon consumption of fenugreek sprouts. Most likely, it was the fenugreek seeds used for sprout production that were the source of the contaminating agent,
E. coli O104:H4, although the pathogen could never be traced back in the seed batches that were used for sprout production [
4]. However, the outbreak strain was exceptional as it did not belong to the more ‘classical’ EHEC outbreak strains such as
E. coli O157:H7, but instead to the group of entero-aggregative
E. coli [
5]. Commonly,
E. coli O104:H4 has a human reservoir and is endemic in central Africa, but has never been reported before to be carried over via plants to humans. Entero-aggregative
E. coli strains are dissimilar from the more common EHEC serotypes that carry the
eae gene on a plasmid of which the product, the outer membrane protein intimin, is responsible for causing the typical intimate attachment to epithelial cells.
Escherichia coli O104:H4 do not produce the intimin protein, but instead form a typical stacked brick-like cell structure on the intestinal cells and from there the bacterial cells release Shiga toxin into the human gut [
6]. The aggregative adherence fimbriae (AAF), expressed from a gene located on the pAA plasmid, are responsible for this typical stacked brick bacterial cell feature, enabling entero-aggregative
E. coli strains to colonize the intestinal mucosa layer. It is a question that still needs to be addressed whether these differences in bacterial attachment to human epithelial cells might also be responsible for eventual different behaviour of
E. coli strains on seeds and on, or inside, plants emerging from seeds.
Plants can be colonized by
E. coli and persistence near plants, on their outside surface and even internally, by different
E. coli strains has been reported over time [
7,
8,
9].
Escherichia coli strains show chemotactic responses to plants and can resist local plant immune responses in the form of oxidative stress [
10,
11] and it is able to protect itself against osmotic stress by conversion of plant derived choline to betaine [
12].
Escherichia coli strains were found at different locations in multiple plant species and most commonly
E. coli showed preference for the root zones and rhizosphere soils [
13,
14,
15]. However, soil management also played an important role in persistence of
E. coli in agricultural systems as the type of soil amendment on
E. coli persistence was stronger than presence or absence of maize roots [
16]. Damaging of seminal roots and root hairs, either mechanically, or caused by the phytopathogenic root-knot nematode
Meloidogyne hapla, did not result in significant higher root internalization by
E. coli, with respect to the undamaged control treatment [
17]. Most of the described studies in plants were undertaken with
E. coli O157:H7 and occasionally with
E. coli K12, but not so often with strains belonging to other
E. coli types, such as entero-aggregative
E. coli strains to which
E. coli O104:H4 belong to.
Escherichia coli is a complex species group and in the past this group was classified on the basis of their colonization behaviour in warm-blooded animals and on the clinical symptoms derived from oral ingestion of pathogenic strains. However, the
E. coli species complex consists of strains whose lifestyles diverge from pathogens to commensals, and their capacity to persist and colonize ecological niches outside the colon is remarkable [
18].
Escherichia coli genomes are flexible, allowing strains easily to adapt to local reigning circumstances [
19,
20,
21]. Therefore, it should be accounted for that
E. coli strains present in soil ecosystems are distinct from the ones in animals [
22,
23]. Human pathogens taxonomically related to
E. coli, such as
Salmonella enterica, also can colonize plants and even can communicate with plant cells [
7,
24,
25]. Plants must be recognized as ‘alternative’ habitats for food-borne enteric bacteria [
26] and Enterobacteriaceae, the taxonomical family to which
E. coli and
S. enterica belong to, are common inhabitants of plants. Particular groups of endophytes and phytopathogens also belong to this group [
21,
27] and therefore Enterobacteriaceae must be considered as an important component of the plant microbiome. Microbiomes of plants will form an important barrier against invasions of human pathogens in plant production systems and Enterobacteriaceae also play an important role in the defence of plants against invasive micro-organisms [
28,
29,
30]. Human pathogens invasive to plants may acquire genetic information from plant-indigenous Enterobacteriaceae making them better adapted to circumstances locally reigning in, and near plants [
21,
31].
In this study we raise the questions as to whether
E. coli O104:H4 is capable to colonize developing plants from seeds and if it would be possible that, via plant internalization,
E. coli cells could reach the embryonic tissue of developing seeds. If the last is possible, then it would mean that, in the worst case,
E. coli O104:H4 could persist in seeds for over many generations and even could be protected from common seed surface disinfection treatments because of the protection provided by the seed coat. Application to plant reproductive organs with the endophyte strain
Paraburkholderia phytofirmans PsJN lead to internal seed contamination resulting in dissemination of this strain to next generations of plants [
32]. Via experimentation, we explored the possibility if this could also be the case for
E. coli. To address this question, we followed the fate of strain 55989 and of two other
E. coli strains from artificially contaminated seeds to growing plants, and from developing flower tissue to mature seeds upon proximate introductions to the plant reproductive organs. We chose for Afila type (dwarf) pea,
Pisum sativum, as model plant because of its fast and robust growth under variable circumstances, but also because of the size and smooth surface structure of the seeds making them easy to handle under the applied experimental circumstances. Experiments were conducted under realistic growth circumstances in greenhouse and open field settings.
Escherichia coli strains differing in origin (plant versus human), adherence properties to epithelial cells (aggregative adherence fimbriae versus surface protein intimin), and virulence profile (possession of EHEC virulence genes or not) were used in our experimentation to relate eventual differences in seed and plant colonization to typical
E. coli properties.
2. Materials and Methods
2.1. Escherichia coli Strains
Entero-aggregative
E. coli O104:H4 strain 55989 was isolated in the period between 1996 and 1999 from faeces of a patient suffering from human immunodeficiency virus (HIV) and persistent diarrhoea from the Central African Republic [
33]. This strain was used in all experiments conducted in this study. Two other strains were used for comparison in pea plant colonization: extended spectrum beta lactamase
E. coli strain 0611 and Shiga toxin producing
E. coli O150:H2 strain N112 (
Table 1). Strain 0611 was obtained from morning glory leaves imported as culinary herb from Thailand [
34], whereas strain N112 was obtained from grass plants taken from a meadow land located in the neighbourhood of Nijkerk, the Netherlands [
23]. All
E. coli strains were grown overnight under shaking (180 RPM) at 37 °C in Luria Bertani broth (LB: Tryptone, 10 g; yeast extract, 5 g; NaCl, 10 g; dissolved in one L water and standard autoclaved for 20 min at 121 °C). Thus obtained late exponential cells were harvested by centrifugation and cell pellets were washed twice in Ringer solution (¼ strength Ringers, OXOID BR0052, Basingstoke, UK; one tablet dissolved in 0.5 L demineralized water followed by autoclaving). Prior to experimentation, optical densities at 600 nm (OD
600) of suspensions were measured and retrospectively validated by colony forming unit (CFU) enumeration onto Brilliance
E.coli/coliform selective agar (BECSA, OXOID CM1046, Basingstoke, UK) medium without further amendments for strain 55989, or onto BECSA amended with 1 µg per ml of cefotaxime for strain 0611 or onto CHROMagar
TM O157 (CHROMagar
TM, Paris, France) for strain N112.
2.2. Pea Seed Treatment with E. coli
The same seed batch of Afila type (dwarf) pea (Pisum sativum) were used throughout all experiments. Microbial analysis of seeds revealed a contaminating fungal load in the seed lot and, therefore, seeds were disinfected with 1% chlorine solution for 10 min. As a result of disinfection, the germination rate of the seeds increased from 84.8% to 93.2%. Disinfected seeds were, therefore, used in all further experiments.
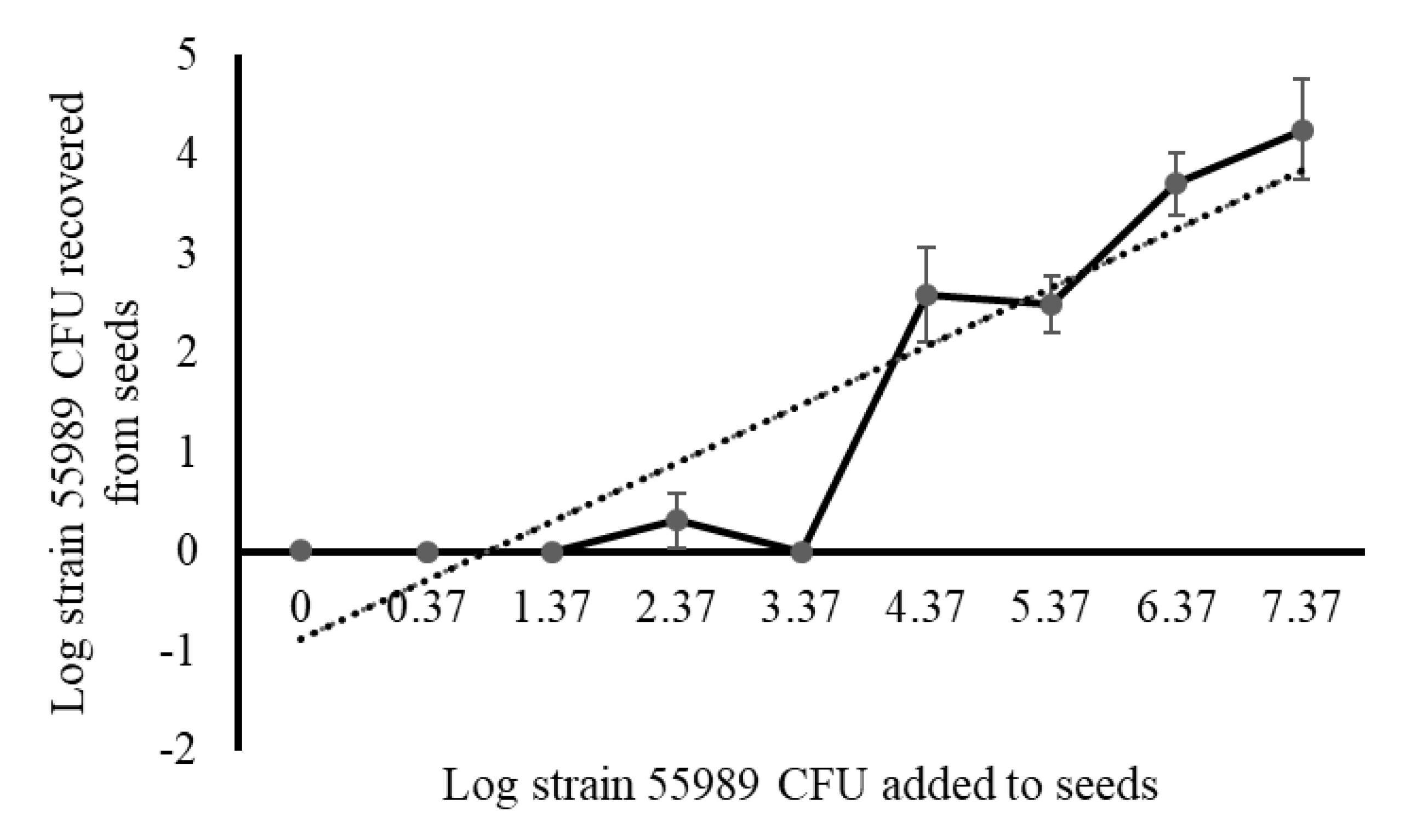
For setting up a standardized seed inoculation protocol, the minimal exposure time and cell density required for efficient attachment of
E. coli cells to the seed surface, performed with strain 55989 only, were established. For establishment of the minimal exposure time, seeds were exposed for 1, 5, 10, 30 and 60 min to exponentially-grown Log
10 (Log) 5.59
E. coli strain 55989 cells per mL and upon exposure, treated seeds were stored for 24 h at room temperature, after which seeds were rinsed twice in sterile Ringer solution to remove loosely attached strain 55989 cells from the seed surface. For establishment of the minimal effective density of strain 55989 cells for seed treatment, seeds were exposed for one h to a range of inocula, 10-fold increasing in densities, starting from Log 0.37 to up to Log 7.37 cells per mL, using sterile water as negative control. After exposure, treated seeds were stored for 24 h at room temperature, after which seeds were washed two times in sterile Ringers solution and 100 µL of washing solutions and smashed pea seed extracts (made by hammering one seed for 30 s in a Bioreba bag [Bioreba AG, Reinach, Switzerland] with three mL Ringer solution) were plated onto BECSA, and plates were incubated under standard conditions, i.e., for 16 h at 37 °C for colony formation. For inactivation of strain 55989 cells attaching to the pea seed surface, three different disinfection solutions; i.e., silver/copper (AMS), chlorine dioxide and hypochlorite, as previously described for lettuce leaf decontamination [
35], were applied. Therefore, 10 µL droplets containing Log 5.79 strain 55989 CFUs were placed onto seed surfaces and incubated for one h at room temperature. Upon incubation, droplets were removed with a pipet tip and treated seeds (
n = 5 per treatment) were incubated for one min in the different disinfecting solutions using sterile water as control. Upon incubation, liquids and smashed pea extracts were plated onto BECSA, and plates were incubated.
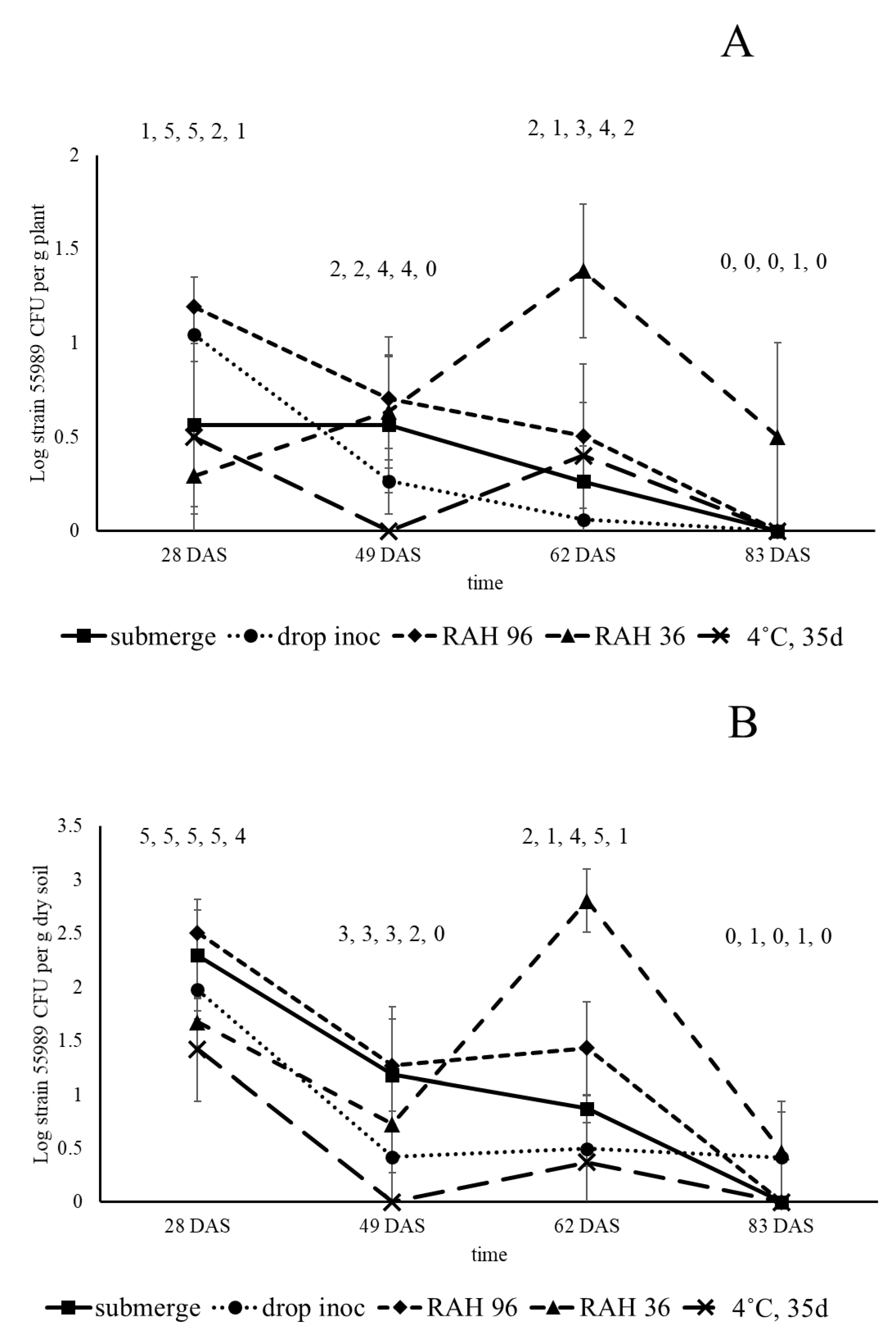
For experimentation in greenhouse and fields, two types of seed inoculation methods were applied: (1) local seed inoculation using 10 µL droplets with E. coli cells (drop inoculation) placed on the seed surface and (2) submergence of seeds in an E. coli cell suspension (seed submergence). For drop inoculation, a single droplet with between Log 7.2 and 7.4 cells per 10 µL were placed on the seed surface and treated seeds were kept in a laminar flow cabinet at room temperature for one h, after which droplets were removed using a sterile pipet tip. For submergence, seeds were incubated for one h in a laminar flow cabinet at room temperature in a washed E. coli cell suspension with densities between Log 9.2 and 9.4 cells per mL. Seeds from all treatments were collected, washed twice in Ringer solution and dried overnight (16 h) in a laminar flow cabinet. For the second field study, conducted in 2016 and executed with strain 55989 only, submerged seeds were washed, dried and stored in a desiccator for 48 h at constant relative air humidity (RAH) of 36% or 96%, or stored for 35 d in a fridge at 4 °C, at ambient RAH. Relative air humidities in the desiccators was maintained at 36% by using a saturated CaCl2 solution and at 96% by using demineralized water.
2.3. Comparison of Three Different E. coli Strains in Pea Plant Colonization
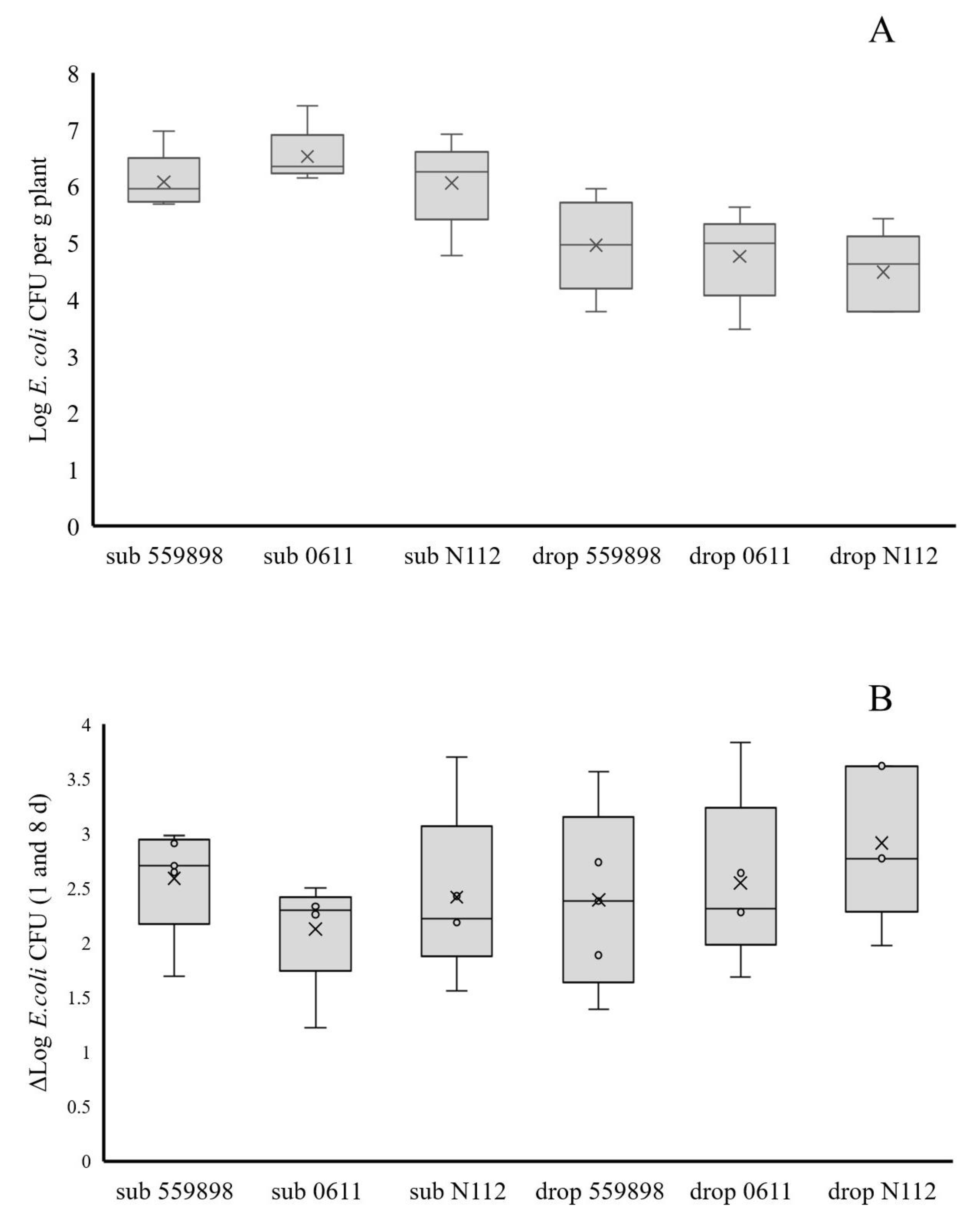
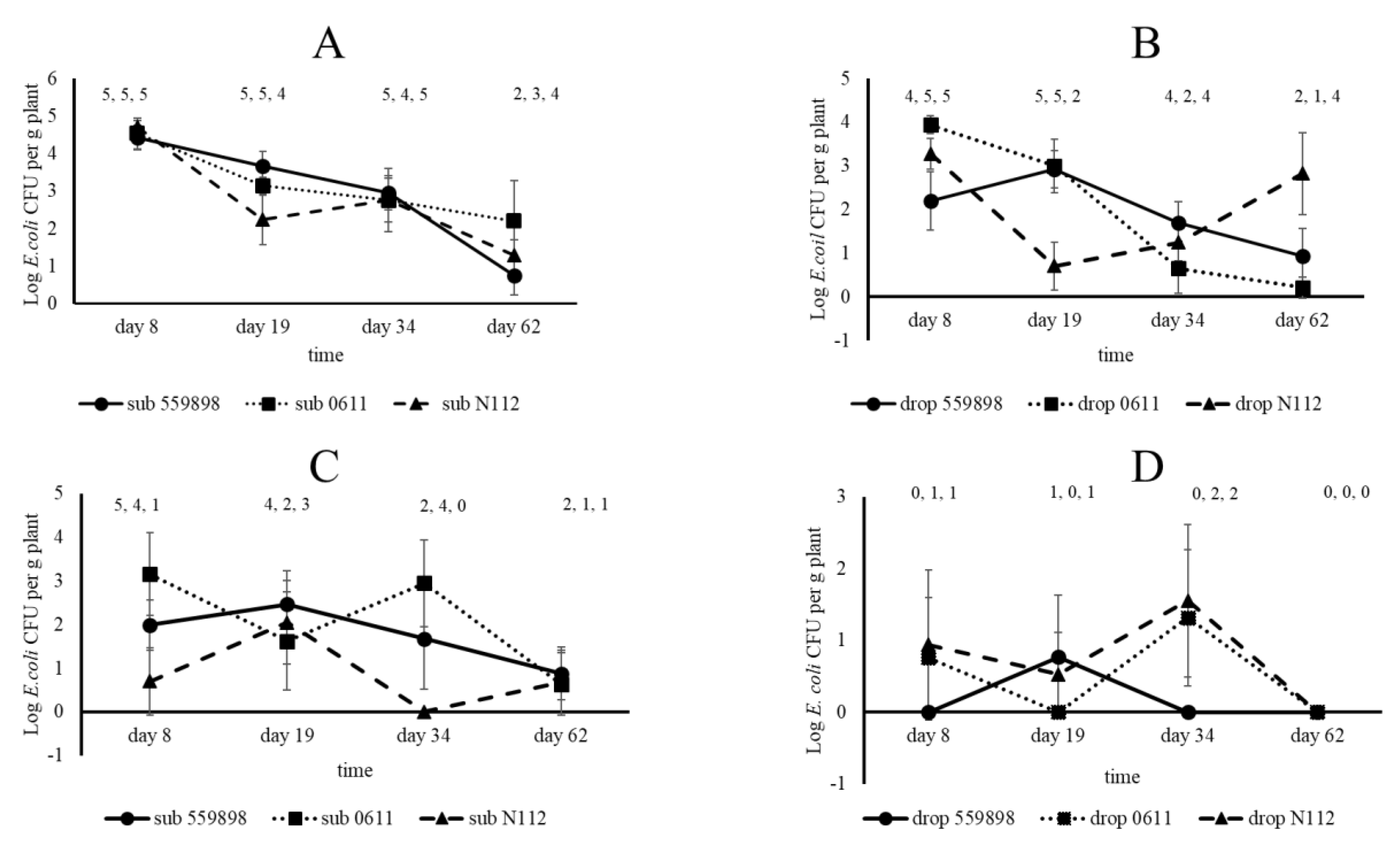
A plant colonization study with three E. coli strains (55989, 0611, N112) in developing pea plants was performed in the greenhouse. For that purpose, seeds were treated with the three strains separately or with water (control) via submergence and drop inoculation (n = 5 per treatment, three strains plus control and two inoculation types, leading to a total of eight treatments). A total of 160 treated, and 80 non-treated (see later) seeds were planted into standard potting soil and treated seeds were allowed to germinate in the dark for three d followed by subsequent plant growth at a day/night regime of 8 h darkness at 18 °C and 16 h light at 22 °C with a constant air humidity of 70%. Densities of the different E. coli strains, using water-treated seeds as controls, were determined by plating onto the agar medium respective for each strain and on BECSA for control seeds, for eventual presence of indigenous E. coli in seeds. Based on different growth stages after 8, 19, 34, 50, 62 and 80 d, plants of all treatments were analysed at different locations for the eventual presence of (inoculated) E. coli. On day 8, the primary roots, cotyledon and epicotyl were analysed, whereas at days 19, 34 and 62 roots, stem base (stem 0.5 cm above the soil until the first true leaf pair) and top stem (stem part between the fourth and fifth true leaf pair) were analysed. At 80 days after sowing (DAS) seeds were collected for later analysis on eventual presence of E. coli. Primary roots, cotyledons and epicotyls at day 8 were taken from plants and transferred to Bioreba bags containing one mL Ringer solution and extracts were made by hammering as described before for treated seeds. These extracts were then serial diluted 10-fold, and non-diluted and diluted samples were plated onto the agar medium respective for each strain and on BECSA for extracts from control plants, and all plates were incubated under standard conditions. Root and stem (stem base and top) samples, taken at 19, 34 and 62 days after sowing (DAS), were transferred to Bioreba bags with three mL Ringer solution after which extracts were made. Non-diluted and tenfold serial diluted samples were then plated onto the different agar media after which E. coli colonies were allowed to grow out.
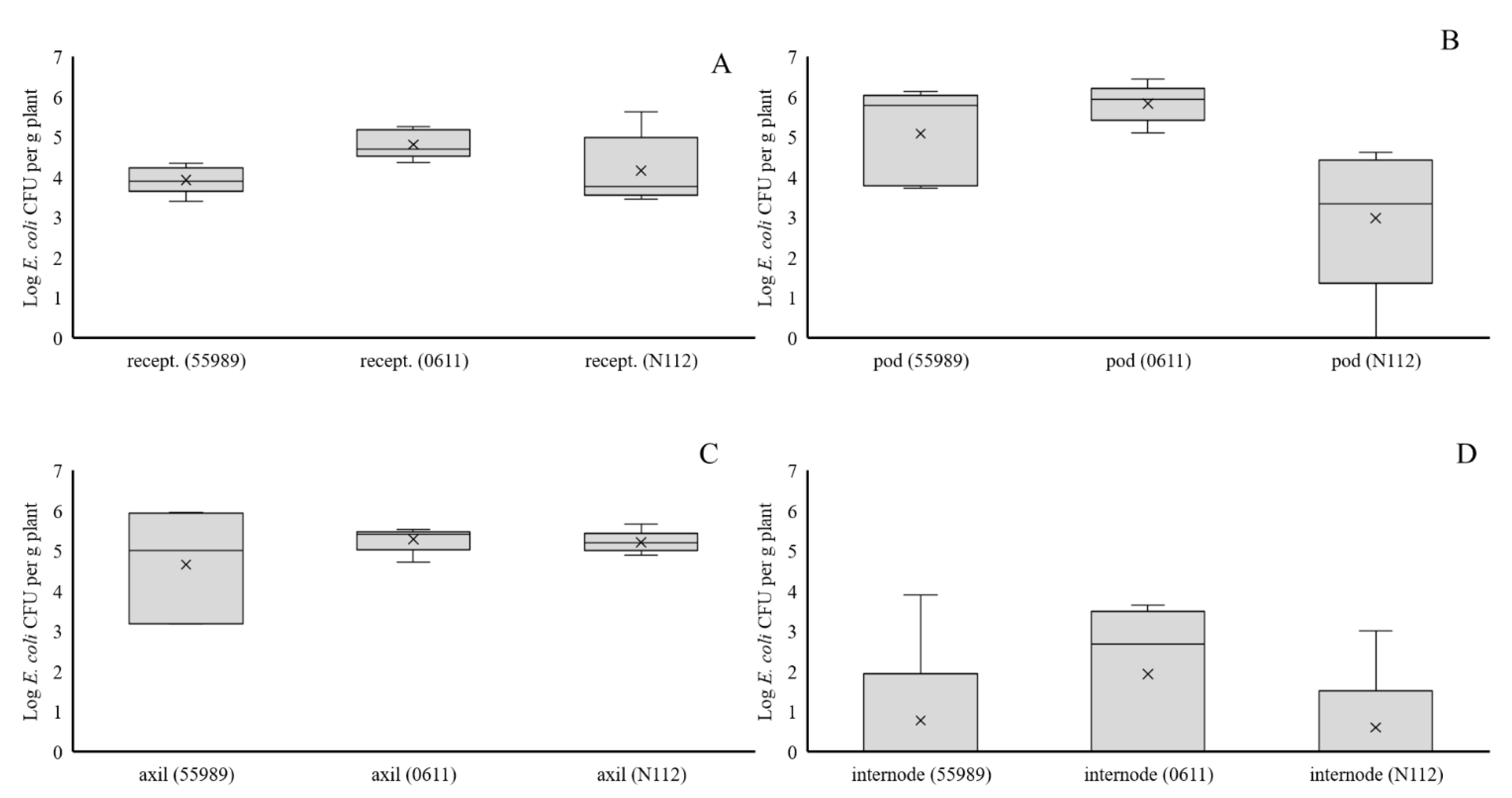
Non-treated plants (80 in total) at day 45 were inoculated with each of the three E. coli strains and water (as control) in the flower receptacle, or in the first leaf axil below the developing flower receptacle (n = 5, three strains plus control and two inoculation types leading to eight treatments) in separate plants to investigate whether local inoculation with E. coli could lead to (internal) seed infection. Therefore, wounds were created by punching with a sterile needle into the flower receptacle and leaf axil tissues, and five µL droplets, containing no (control) or between Log 6.4 and 6.6 E. coli CFUs, were placed on top of these wounds. Two hours after applications, the droplets were taken up by the plants and five and 35 d after inoculation, at 50 and 80 DAS respectively, treated plants were analysed for the local presence of E. coli at the inoculation sites and/or in developing pods and seeds. At 50 DAS, only developing pods were analysed for plants inoculated at flower receptacles, whereas for plants inoculated at the leaf axils, developing pods, leaf axil and internode tissue were sampled. At 80 DAS, developed pods and seeds were sampled for both treatments. Samples taken from all tissues, of between 0.3 and 2 g in weight, were further processed and plated on agar media for E. coli colony formation. Ripened seeds from plants of all 16 treatments, taken at 80 DAS (one seed per plant; n = 5 per treatment) were collected and stored at 4 °C for later analysis on presence of E. coli.
2.4. Colonization of Pea Plants by Strain 55989, Introduced onto Seeds, under Open Field Circumstances
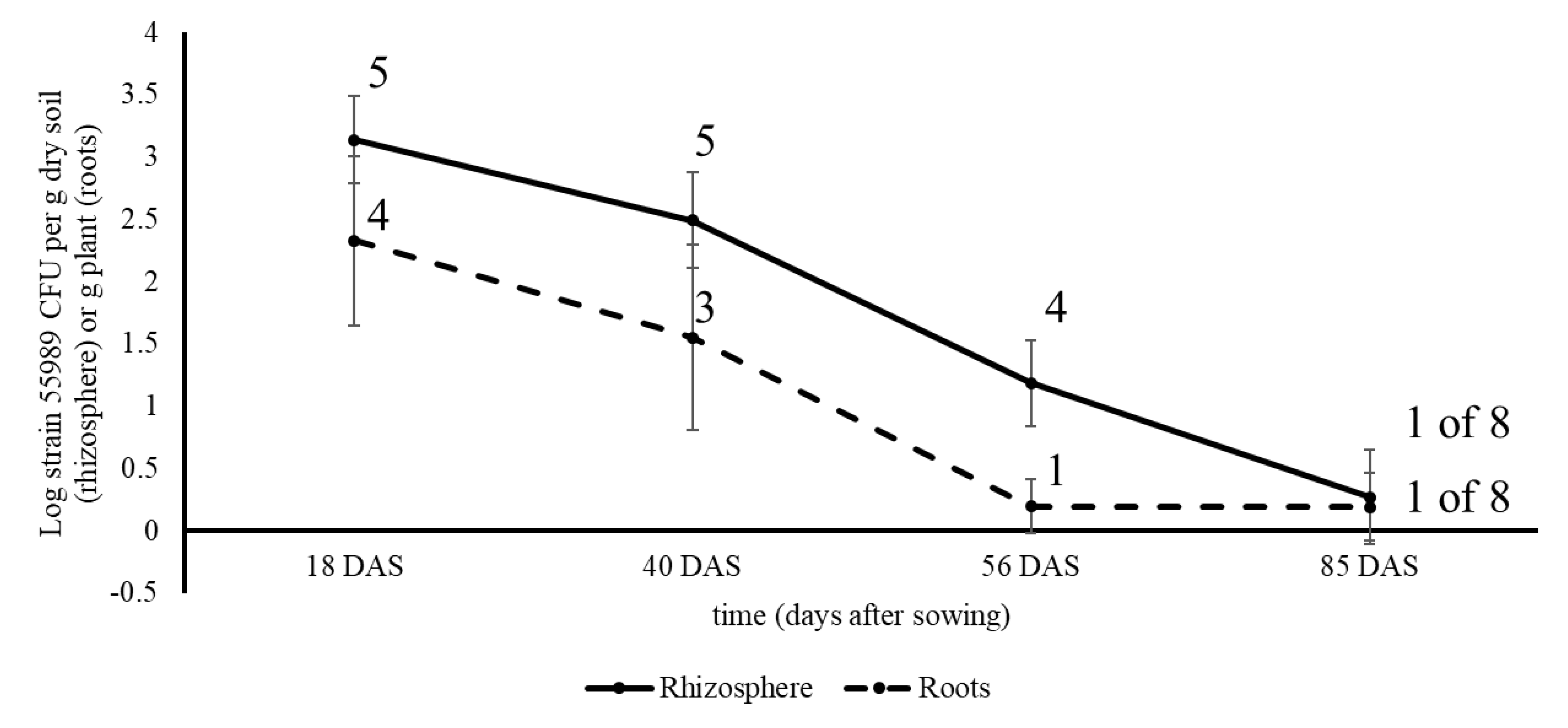
Field studies in two consecutive years were conducted with pea plants grown from water (control) and strain 55989-treated seeds in the periods between 28 May and 20 August 2015 and between 26 May and 17 August 2016. Both studies were performed in an agricultural field at the Unifarm experimental farm of Wageningen University and Research centre (GPS coordinates, 51°59′18.0″ N; 5°39′40.1″ E). The field was 4 m × 6 m in size and was covered by a net to avoid entrance of birds into the field. The soil at the Unifarm field location was a sandy soil consisting of 86% sand, 9% silt, 1% clay, organic matter content of 3.9% and pH of 5.9. In the first field experiment, one seed treatment was applied; i.e., seeds submerged in strain 55989 solution. Seeds submerged in water served as control and for each sampling, five plants per treatment and sample time were used. Plants were sampled for analysis in the presence of strain 55989 in rhizosphere soil, roots, stem base and seeds. For the second field experiment, five seed treatments were applied that are realistic under commercial seed treatment and storage circumstances: (1) submergence, (2) drop inoculation, (3) drop inoculation followed by 48 h storage at 96% RAH, (4) same as 3, but then stored at 36% RAH, and (5) submerged and stored for 35 d at 4 °C. Seeds submerged in water were used as control, leading to six treatments using five plants per treatment and sampling time. At 0 DAS (28 May 2016) seeds were sown into the soil and at 28, 49, 62 and 83 DAS, plants were sampled and analysed for presence of strain 55989 in rhizosphere soil, roots and seeds. Sampling times were chosen on the basis of the same plant developmental stages as in the first field experiment. At the first, second, third and fourth samplings in 2015 and 2016, plants were, respectively, in the seventh true leaf stage, in the flowering stage, at the end of flowering/beginning of the pod filling stage, and at the senescent stage with ripened seeds. Plant development was slower in the second than in the first year of field experimentation, due to lower temperatures in 2016, and therefore samples were taken at later time points after sowing in 2016 than in 2015. For each sampling, plants with roots were taken from the soil and soil loosely attached to roots was removed by shaking. Intact plants were separately packed in plastic bags and transported to the laboratory where all samples were processed on the same day. Soil firmly attached to roots was considered to be ‘rhizosphere soil’ and per plant between one and seven g mixed roots with soil were shaken in sterile 50 mL plastic tubes filled with one g gravel and 20 mL 0.1% sodium pyrophosphate solution. Tubes were vortexed for one min and soil suspensions were either plated non-diluted onto BECSA, or in 10-fold serial dilutions made in 0.1% sodium pyrophosphate solution. Then, roots were removed from the tubes, two times rinsed in sterile demineralized water and pat dried on sterile filter paper. Stem base parts (only taken in 2015 and between 0.2 g and maximally 3 g in weight) and rinsed roots (between 0.5 and 4 g) were transferred to Bioreba bags containing three mL Ringer solution, after which stem base and root parts were hammered and thus derived extracts were plated undiluted and tenfold serial diluted. Individual purple stained colonies, indicative for strain 55989, at the last samplings in 2015 and 2016 (respectively, at 85 and 83 DAS) were streaked to purity onto BECSA and single colonies were grown overnight in LB at 37 °C. Resulting dense cultures were either mixed with sterile glycerol to a final concentration of 20% for storage at –70 °C, or cells from one mL culture suspensions were pelleted by centrifugation after which DNA was extracted from resulting cell pellets for whole genome sequencing, using the Illumina MiSeq platform. Comparison of obtained isolates with strain 55989 and other
E. coli strains, as described in van Overbeek et al. [
23], was made on the basis of DNA sequences of 10 cellular household genes, i.e., adenylate kinase (
adk), classII fumarate hydratase (
fumC), glycerol kinase (
glpK), DNA gyrase subunit B (
gyrB), 3-isopropylmalate dehydrogenase (
icd), diaminopimelate decarboxylase (
lysA), malate/lactate/ureidoglycolate dehydrogenase (
mdh), methionine-tRNA ligase (
metG), adeylosuccinate synthetase (
purA), and DNA recombination/repair protein (
recA).
Seeds, five per treatment (two in field experiment 1 and six in field experiment 2) from different plants taken at the last samplings in both years, were collected and stored at 4 °C for later analysis for eventual presence of E. coli.
2.5. Analysis for Presence of E. coli in Surface-Sterilized Seeds
All collected seeds from greenhouse and both field experiments, coming to a total of 120 seeds, were surface disinfected using 1% chlorine solution as described before. Final wash water was plated onto the agar medium respective for each E. coli strain, using BECSA for water-treated controls. Disinfected seeds were allowed to germinate on sterile water-soaked filter paper in Petri dishes for 6 d in the dark at room temperature. Then, emerging plants were transferred to Bioreba bags containing 3 mL Ringers solution, hammered and resulting extracts were directly plated onto the agar medium respective for each strain, using BECSA for extracts from plantlets grown from water-treated seeds (control extracts). In parallel, these control extracts were also spiked with strain 55989 (to an estimated final density of Log 4 CFU per mL) and plated onto BECSA as a control for the establishment of eventual E. coli growth inhibition in pea seed extracts.
2.6. Data Processing and Statistics
E. coli densities on seeds, in root, pod or stem tissue, and in rhizosphere soil were expressed as Log10 (n + 1) values per seed, per g of plant or per g of dry soil, respectively. Prevalence was defined as the number of samples with detectable (introduced) E. coli CFUs. When absent in a sample, introduced E. coli was considered to be undetectable and the lowest limit of detection was calculated on the basis of lower than one single colony on duplicate plates that had received non-diluted extracts. These numbers are expressed as ‘0’ values. Geometric means were calculated from Log-converted values and used for statistical analysis of variance (ANOVA, Genstat 19th Ed. Hemel Hempstead, UK). Differences were considered to be different at levels of p = 0.05 and below.
4. Discussion
Entero-aggregative
E. coli strain 55989 was able to colonize the root compartment of pea plants from inoculated seeds. In the root and rhizosphere soil, the strain could persist until the senescent stage of plant growth, when seeds had ripened. Colonization of the above-soil parts of the plants by strain 55989 was negligible and, therefore, translocation of strain 55989 cells via the vascular tissue of the stems to developing seeds seems unlikely. The fact that even proximate introductions of cells of all three strains to developing flowers did not result in effective internal seed contamination led to the conclusion that there is no evidence for internal transmission of
E. coli via plants to developing seeds. The consequence thereof is that
E. coli O104:H4 will not be transmitted as internal seed contaminant to next generations of plants, at least not in pea plants. However, as external seed contaminant,
E. coli O104:H4 will be transmitted to next generations of plants and its ecological behaviour in plants is the same as for the two other
E. coli strains. This is an important aspect because it demonstrates that pathogenic
E. coli strains can contaminate crop production sites by making use of seeds as primary vectors for transmission to growing plants. The soil environment plays an important role in further transmission of these pathogens to edible parts of the plants. Our finding corroborate that of Habteselassie et al., [
14], where it was demonstrated that the rhizosphere played an important role in the early establishment and colonization of radish and lettuce plants by
E. coli O157:H7.
The two
E. coli phylotype B1 strains used in our study (O104:H4 strain 55989 and O150:H2 strain N112) were indistinguishable in their ecological behaviour in, and near developing pea plants, this in spite of their differences in pathotype. The behaviour of the two phylotype B1 strains on their turn were indistinguishable from the third one, the ESBL-producing
E. coli strain 0611 that originated from plants. Niche partition and adaptation to natural habitats has been hold responsible for diversification of different
E. coli clades in their ecological behaviour in many natural habitats such as in soils [
18,
19,
22,
36], but most likely this does not hold true for plant ecosystems. In plants and their surrounding areas (rhizosphere soil), plant-released nutrients are available for cell proliferation and like many bacteria naturally occurring in soils,
E. coli profit from available nutrients explaining the long-term persistence near pea roots and in rhizosphere soils as demonstrated in both field experiments. The three
E. coli strains declined over time in the pea rhizosphere which contrasted the ecological behaviour of the typical soil-borne bacterial species
Pseudomonas fluorescens that persisted at constant and higher levels in the pea rhizosphere [
37]. Although not a species fully adapted to soil environments,
E. coli responded here in an opportunistic fashion to nutrients made available by plants, either at seed emergence or at root exudation. The fact that
E. coli isolates were found in pea rhizosphere soil and roots that differed from the introduced strain 55989 indicate that
E. coli is intrinsically present in pea plants or their proximate surroundings. Therefore, these strains must be considered as indigenous inhabitants of the pea plant microbiome. This would also indicate that these
E. coli strains are adapted to life near or even inside plants. The presence of
E. coli in plants is still an unknown fact and would require further attention in eventual later studies. Altogether, this emphasizes the fact that plants must be considered as alternative ecosystems for
E. coli [
26].
The above soil compartments of the pea plants were less colonized than the roots by all three
E. coli strains, indicating that these parts of the plants are less favoured for colonization by
E. coli in general. Most likely nutrient availability was limited at the epicotyl from where later at plant growth the stem base developed. Possibly a part of the
E. coli populations colonized the internal tissue of the stem base, but because no surface sterilization attempts were undertaken in our study, no information can be provided on exact numbers of
E. coli cells internalized into plants. Internalization of spinach, lettuce and
Nicotiana benthamiana by
E. coli O157:H7 was reported in Wright et al. [
9] and internalization was indicated to play a critical role with respect to food safety and human health [
8]. However, food safety was not the principal motive for our research, but instead it was the ecological behaviour in, or on plants, in relation to colonization of developing seeds. From our study we can conclude that either via external, or internal colonization, even upon introduction shortly before flower formation, no internal seed colonization by any of the three studied
E. coli strains took place. From there, it can be concluded that
E. coli, under realistic growth circumstances, is not able to cross the barrier between maternal and embryonic tissue in pea plants. This in contrast to a typical endophyte,
Paraburkholderia phytofirmans PsJN, that was able to colonize internal seed compartments of maize, soy and pepper upon flower spraying [
32]. Most likely,
E. coli lack the appropriate cellular equipment to independently penetrate plant tissue like endophytes do. Penetration of
E. coli into seeds can be forced, e.g., by vacuum infiltration [
38], but these are circumstances that do not occur under realistic plant growth circumstances.
From seeds,
E. coli and
S. enterica strains were shown to colonize cotyledon and the primary roots of emerging alfalfa, fenugreek, lettuce and tomato plants [
38], in a fashion comparable to what was shown in our study with the three
E. coli strains in pea plants. In the study done by Liu and co-workers [
38], the pathogens were present under the seed coat, whereas in our study, the
E. coli strains were applied on the seed coat. It seems that no difference exist in colonization of plants either by
E. coli pathogens present inside or on the surface of seeds. In our study, the time needed for attachment to pea seeds was within one minute upon exposure and no differences in efficiency of attachment to the pea seed surface was observed between the three strains. In Cui et al. [
2], it was reported that differences in seed attachment between
E. coli O157:H7 and O104:H4 was present on different seed types (alfalfa, fenugreek, lettuce and tomato).
Escherichia coli O104:H4 strain ATCC BAA2326 (Hamburg 2011 outbreak strain) showed the lowest attachment potential in comparison with three
E. coli O157:H7 strains. Lowest attachment may be related to relative higher EPS production by
E. coli O104:H4 strains, favouring biofilm formation, but restricting adherence to solid surfaces [
2]. Also, higher cell hydrophobicity and rougher (more wrinkled) seed surface type often result in higher attachment of bacterial pathogens to seeds [
2]. So, in spite of the facts that: (i)
E. coli has a relatively lower surface hydrophobicity, (ii)
E. coli O104:H4 is less efficient in attachment to solid surfaces, and (iii) that the surface of pea seeds is rather smooth, we still observed long-term persistence to up to 35 days of
E. coli strain 55989 on the pea seed surface, and this strain was also shown to stay alive at low RAH of 35% for 48 h on the pea seed surface. In line with observations made by van der Linden et al. [
3], it must be concluded that
E. coli can persist over long periods in time on seed surfaces and these time periods and storage conditions are realistic in commercial seed production and logistics circumstances. It can therefore be concluded that presence of
E. coli on seeds, used for starting materials in fruit and vegetable production, pose a potential risk for food safety unless these seeds are appropriately disinfected.
It was demonstrated before that pea seeds can become contaminated with human pathogens as demonstrated at the
Campylobacter jejuni outbreak in Alaska in 2008 [
39]. In case human pathogens are present on seeds used as starting materials for cultivation of crop plants, then a risk on contamination of the edible parts of the plants may exist. From contaminated seeds, human pathogens can reach rhizosphere soil from where they can contaminate edible parts grown below the soil surface, as will be the case for radish and carrot plants, or the edible parts shortly above the soil surface, as will be the case for leafy greens. With regard to the consequences on long-term persistence of pathogenic
E. coli strains in soils of plant production systems, nothing is certain at the moment. Phytopathogens may create ports of entrances for human pathogens from where these pathogens further spread to the edible parts. However, no internalization occurred in spinach plants by
E. coli O157:H7 when two pathogens,
Pseudomonas syringae and
Meloidogyne hapla, were present in the production site [
17], indicating that plant pathogens do not play a dominant role in contamination of plants by human pathogenic
E. coli. However, gene fluxes in
E. coli are high [
18,
36] and auxiliary genomes of
E. coli are often prone to homologous recombination [
20]. Adaptation of pathogenic
E. coli strains to the plant environment might result from gene acquisition events taking place in plants and their surrounding areas [
18,
31]. On the longer run this may result in selection of new types of pathogenic
E. coli strains with improved capacities to colonize plants [
21,
31]. Therefore, longer persistence of human pathogens in plant environments might become a concern for fresh food production.
In summary, no evidence was provided on seed internalization via growing plants by the entero-aggregative E. coli strain 55989 and the other two tested E. coli strains. Internalization of E. coli into seeds, followed by transmission to next generations of crop plants, must be regarded as an unlikely event under realistic plant growth circumstances. However, E. coli strain 55989 could persist on the pea seed surface and was shown to survive under commercially applied seed storage conditions. All three E. coli strains showed stronger preference for the root-soil zones of growing pea plants than for the above soil plant compartments, in spite of their differences in clinical behaviour and origin. Therefore, these observations can be expanded to broader groups of (pathogenic) E. coli strains with the concept that contaminating E. coli strains will colonize root compartments of crop plants in production systems and from there are able to further spread to consumable parts of plants. Adequate seed disinfection protocols should avoid contamination of arable soils used for cultivation of freshly consumable crop plants.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}