
New Insights into the Relationships between Bacterial Dynamics and Water Quality of Aquaculture Systems Supplemented with Carbon Source and Biofilm Substratum

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiments and Reagents
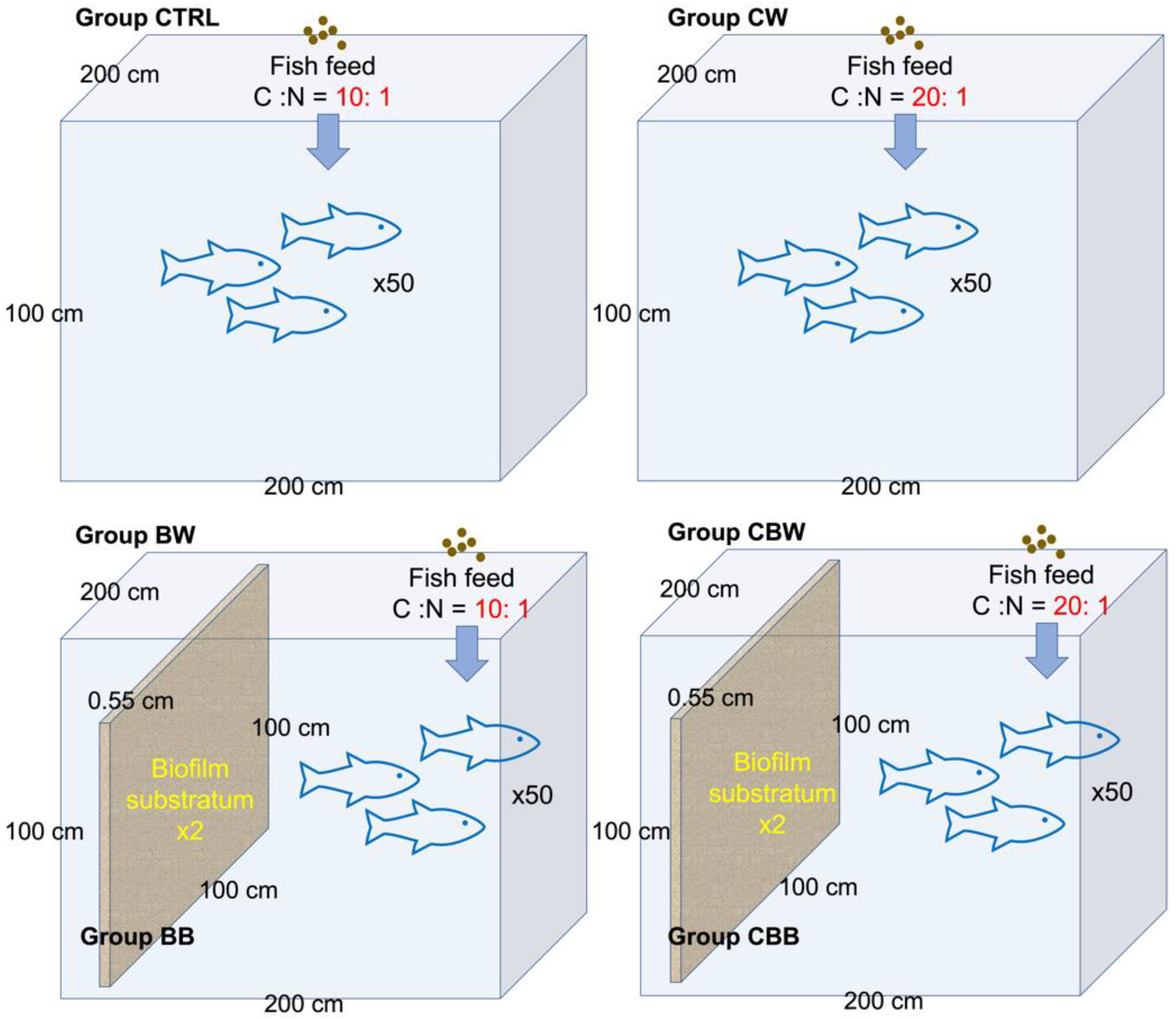
2.1.1. Mesocosm Systems Set-Up and Sample Collections
2.1.2. Water Quality Monitoring and Fish Growth
2.1.3. DNA Extraction and 16S rRNA Amplicon Sequencing
2.2. Data Processing and Statistical Analysis
2.2.1. Measurement of Fish Growth and Water Quality
2.2.2. 16S rRNA Amplicon Data Processing and Taxonomic Classification
2.2.3. Diversity Analysis
2.2.4. Relative Abundance of the Bacterial Community
2.2.5. Bacterial Distinction
2.2.6. Genera Co-Occurrence Network
3. Results
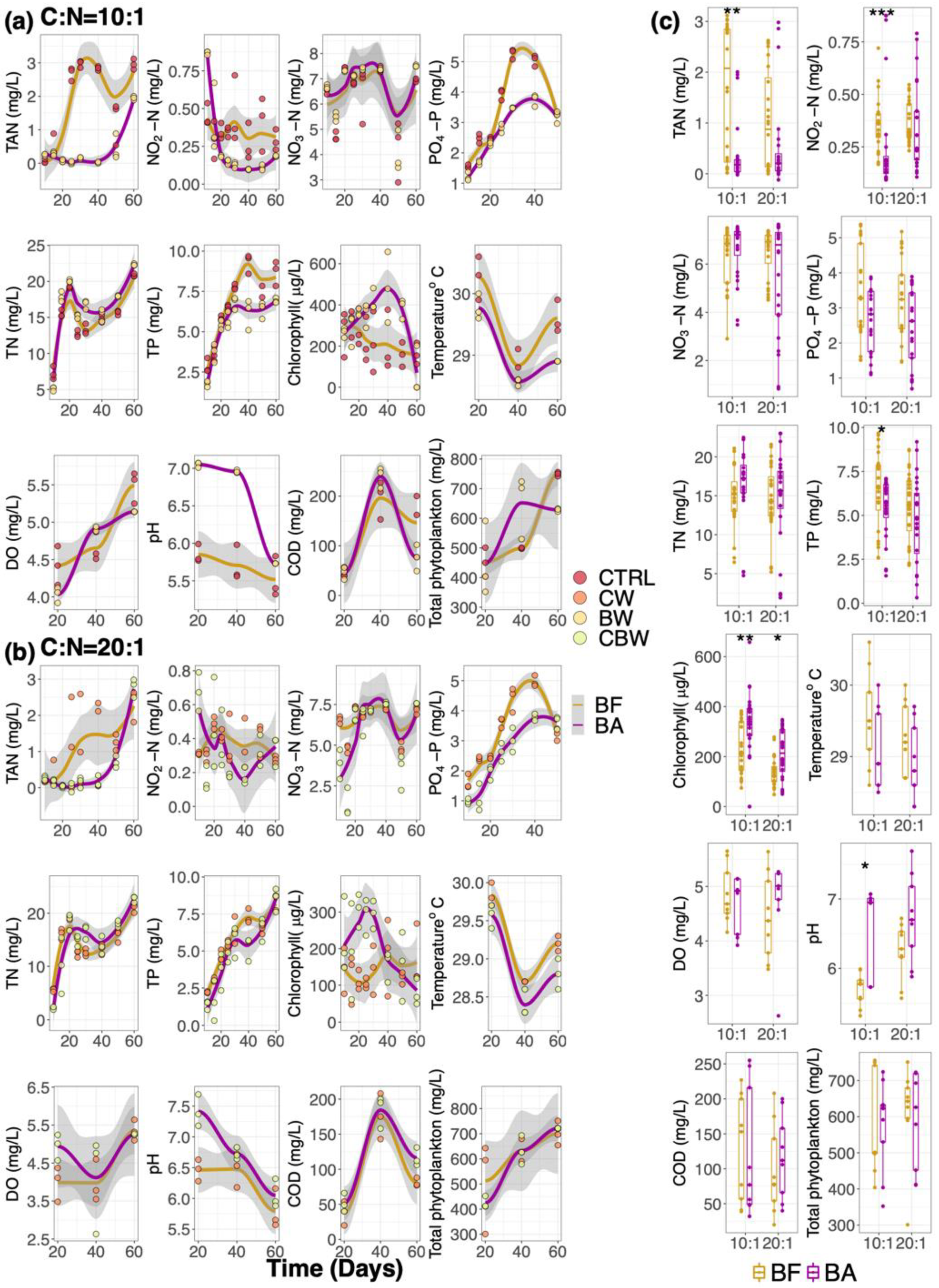
3.1. Fish Growth and Water Quality
3.1.1. Fish Weight and WGR
3.1.2. Water Quality Parameters
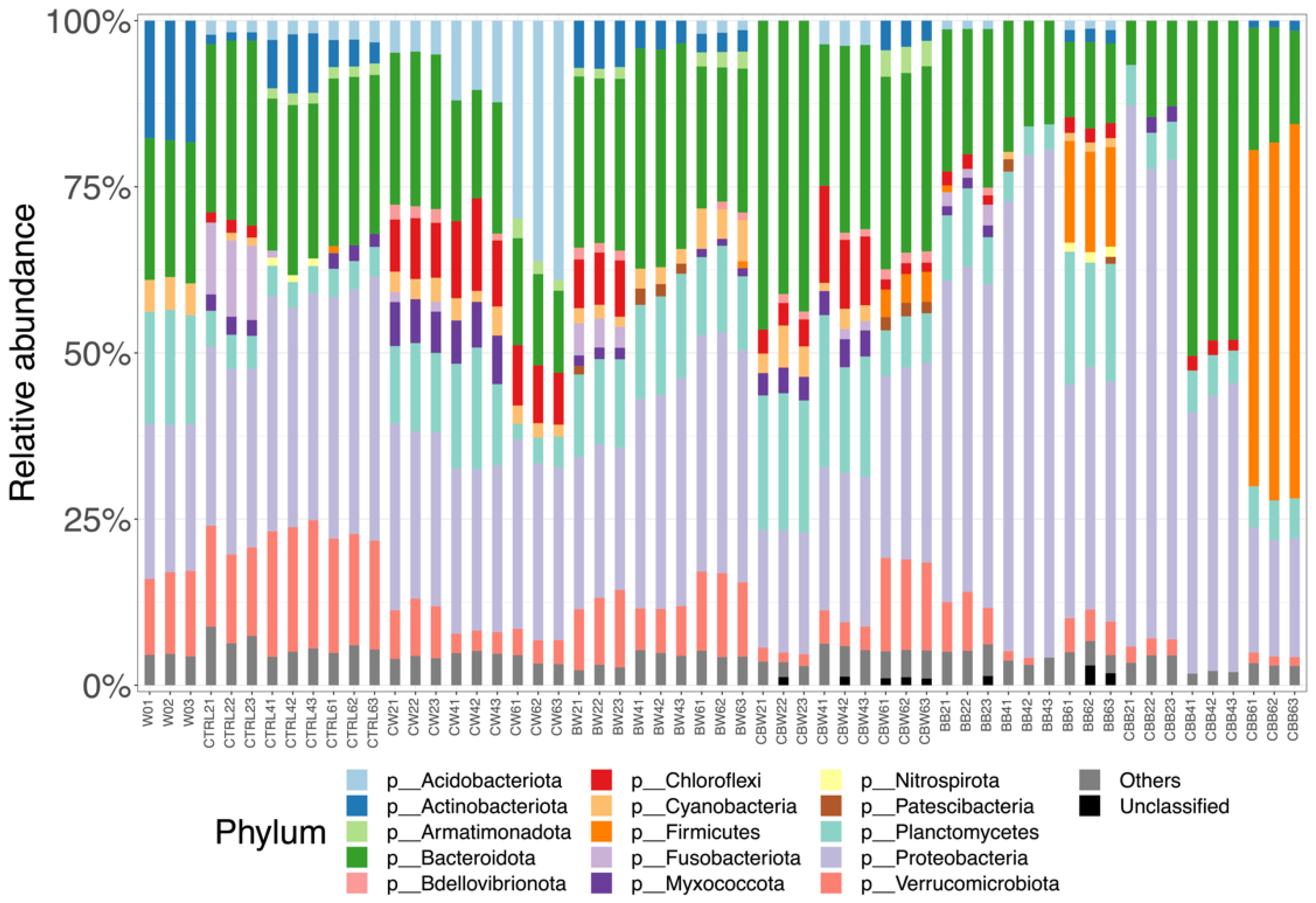
3.2. Bacterial Community in Water and Biofilm-Substrata Samples
3.2.1. 16S rRNA Data
3.2.2. Alpha Diversity and Beta Diversity Analysis
3.2.3. Bacterial Composition at Phylum Level
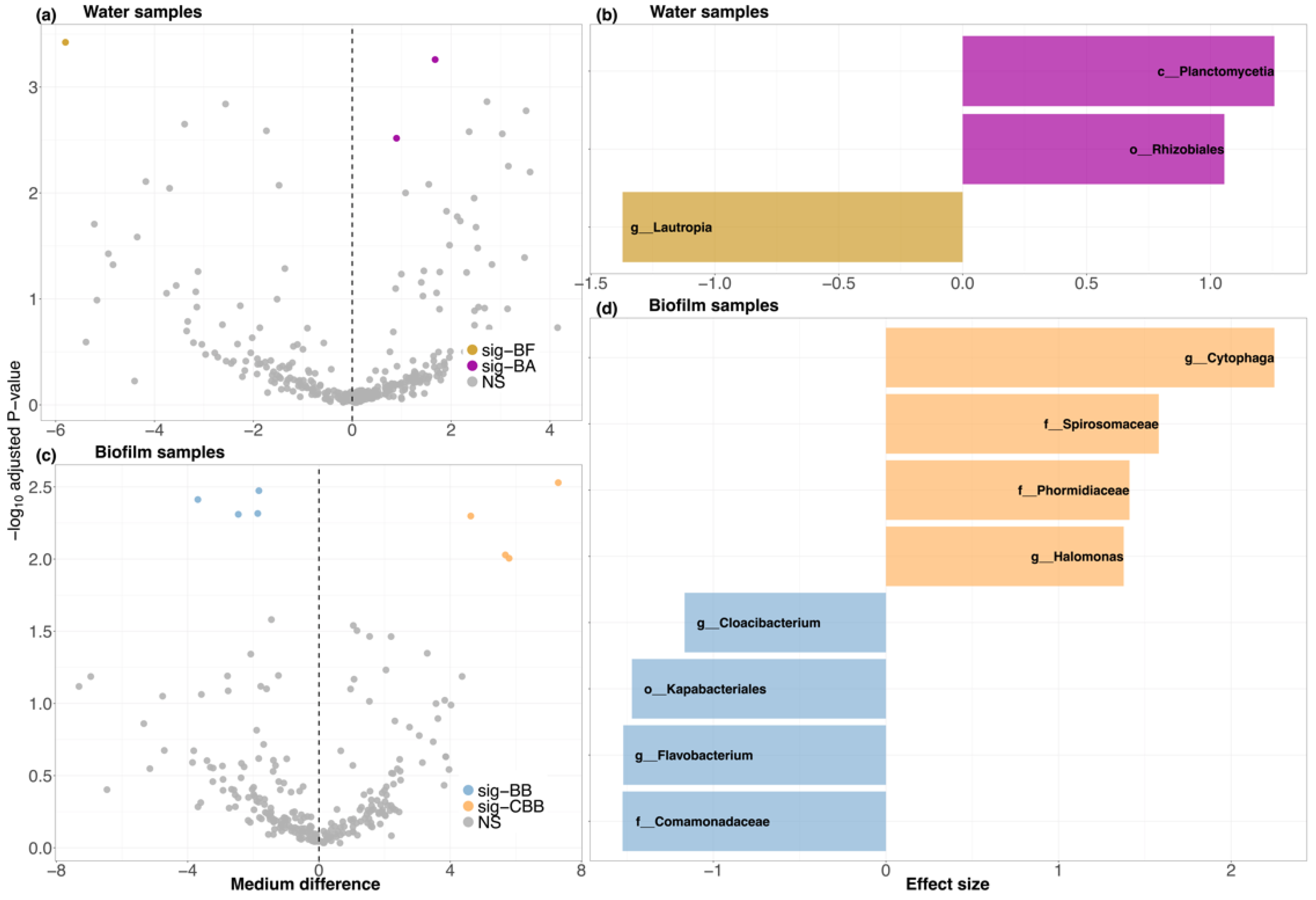
3.2.4. Bacterial Distinction
3.2.5. Genera Co-Occurrence Network
4. Discussion
4.1. Biofilm Substratum Shifted the Predominance of Bacteria in Mesocosm Aquaculture Systems
4.2. Biofilm Substratum Addition Controlled Ammonia Nitrogen and Benefited Fish Growth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018-Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Ahmed, N.; Thompson, S. The blue dimensions of aquaculture: A global synthesis. Sci. Total Environ. 2019, 652, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Wu, X.; Li, L.; Lv, Z.; Zhang, Z.; Hao, A.; Iseri, Y.; Kuba, T.; Zhang, X.; Wu, W.M.; et al. Pollution control and in situ bioremediation for lake aquaculture using an ecological dam. J. Clean. Prod. 2018, 172, 2256–2265. [Google Scholar] [CrossRef]
- Bosma, R.H.; Verdegem, M.C.J. Sustainable aquaculture in ponds: Principles, practices and limits. Livest. Sci. 2011, 139, 58–68. [Google Scholar] [CrossRef]
- Abdou, K.; Ben Rais Lasram, F.; Romdhane, M.S.; Le Loc’h, F.; Aubin, J. Rearing performances and environmental assessment of sea cage farming in Tunisia using life cycle assessment (LCA) combined with PCA and HCPC. Int. J. Life Cycle Assess. 2018, 23, 1049–1062. [Google Scholar] [CrossRef]
- McMahon, K.D.; Read, E.K. Microbial Contributions to Phosphorus Cycling in Eutrophic Lakes and Wastewater. Annu. Rev. Microbiol. 2013, 67, 199–219. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Wang, X.; Olsen, L.M.; Reitan, K.I.; Olsen, Y. Discharge of nutrient wastes from salmon farms: Environmental effects, and potential for integrated multi-trophic aquaculture. Aquac. Environ. Interact. 2012, 2, 267–283. [Google Scholar] [CrossRef] [Green Version]
- Woodhouse, J.N.; Kinsela, A.S.; Collins, R.N.; Bowling, L.C.; Honeyman, G.L.; Holliday, J.K.; Neilan, B.A. Microbial communities reflect temporal changes in cyanobacterial composition in a shallow ephemeral freshwater lake. ISME J. 2016, 10, 1337–1351. [Google Scholar] [CrossRef]
- Berga, M.; Székely, A.J.; Langenheder, S. Effects of disturbance intensity and frequency on bacterial community composition and function. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Pollock, F.J.; Lamb, J.B.; Van De Water, J.A.J.M.; Smith, H.A.; Schaffelke, B.; Willis, B.L.; Bourne, D.G. Reduced diversity and stability of coral-associated bacterial communities and suppressed immune function precedes disease onset in corals. R. Soc. Open Sci. 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacon, A.G.J. Trends in Global Aquaculture and Aquafeed Production: 2000–2017. Rev. Fish. Sci. Aquac. 2019, 2016, 1–14. [Google Scholar] [CrossRef]
- Wang, G.; Yu, E.; Xie, J.; Yu, D.; Li, Z.; Luo, W.; Qiu, L.; Zheng, Z. Effect of C/N ratio on water quality in zero-water exchange tanks and the biofloc supplementation in feed on the growth performance of crucian carp, Carassius auratus. Aquaculture 2015, 443, 98–104. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Yang, L.; Chen, H.; Zhang, S.; Zhao, H.; Zhang, N. Allelopathic control of cyanobacterial blooms by periphyton biofilms. Environ. Microbiol. 2011, 13, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Y.; Feng, L.J.; Zhu, L.; Xu, J.; Ding, W.; Qi, H.Y. Biofilm formation and microbial community analysis of the simulated river bioreactor for contaminated source water remediation. Environ. Sci. Pollut. Res. 2012, 19, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Janssen, S.; Mcdonald, D.; Gonzalez, A.; Navas-molina, J.A.; Jiang, L.; Xu, Z. Phylogenetic Placement of Exact Amplicon Sequences. MSystems 2018, 3, e00021-18. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; Desantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Chao, A. Estimating the Population Size for Capture-Recapture Data with Unequal Catchability. Biometrics 1987, 43, 783. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley Stats Ref: Statistics Reference Online; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 1–15. ISBN 9781118445112. [Google Scholar]
- Fernandes, A.D.; Reid, J.N.S.; Macklaim, J.M.; McMurrough, T.A.; Edgell, D.R.; Gloor, G.B. Unifying the analysis of high-throughput sequencing datasets: Characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2014, 2, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doncheva, N.T.; Assenov, Y.; Domingues, F.S.; Albrecht, M. Topological analysis and interactive visualization of biological networks and protein structures. Nat. Protoc. 2012, 7, 670–685. [Google Scholar] [CrossRef]
- Rawat, S.R.; Männistö, M.K.; Bromberg, Y.; Häggblom, M.M. Comparative genomic and physiological analysis provides insights into the role of Acidobacteria in organic carbon utilization in Arctic tundra soils. FEMS Microbiol. Ecol. 2012, 82, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Gómez, B.; Richter, M.; Schüler, M.; Pinhassi, J.; Acinas, S.G.; González, J.M.; Pedrós-Alió, C. Ecology of marine bacteroidetes: A comparative genomics approach. ISME J. 2013, 7, 1026–1037. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Xu, J.; Xu, X.; Wang, Q.; Kong, Q.; Xu, F.; Du, Y. Organ-specific responses to total ammonia nitrogen stress on juvenile grass carp (Ctenopharyngodon idellus). Environ. Sci. Pollut. Res. 2019, 26, 10826–10834. [Google Scholar] [CrossRef]
- Tuson, H.H.; Weibel, D.B. Bacteria–surface interactions. Soft Matter 2013, 9, 4368. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.N.; Méndez–García, C.; Geier, R.R.; Iakiviak, M.; Chang, J.; Cann, I.; Mackie, R.I. Metabolic networks for nitrogen utilization in Prevotella ruminicola 23. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kartal, B.; Kuenen, J.G.; Van Loosdrecht, M.C.M. Sewage treatment with anammox. Science 2010, 328, 702–703. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mesocosm Systems | Water Samples | Biofilm Substrata Samples | Carbon Source Level (C:N) | Biofilm Substrata | WInitial | WFianl | WGR (%) | FCR |
|---|---|---|---|---|---|---|---|---|
| 1 | CTRL | 10:1 | Free | 120.1 ± 1.8 | 187.1 ± 27.7 | 33.86 ± 11.65 | 3.35 ± 0.08 | |
| 2 | CW | 20:1 | Free | 120.4 ± 0.9 | 130.3 ± 8.3 | 7.82 ± 4.92 *** | 2.87 ± 0.17 *** | |
| 3 | BW | BB | 10:1 | Added | 120.6 ± 1.1 | 221.7 ± 31.2 | 43.93 ± 8.32 *** | 3.10 ± 0.11 *** |
| 4 | CBW | CBB | 20:1 | Added | 120.2 ± 1.4 | 252.5 ± 41.9 | 52.04 ± 7.81 *** | 2.21 ± 0.14 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, E.-M.; Li, Z.-C.; Li, Z.-F.; Wang, G.-J.; Xia, Y.; Zhang, K.; Tian, J.-J.; Gong, W.-B.; Xie, J. New Insights into the Relationships between Bacterial Dynamics and Water Quality of Aquaculture Systems Supplemented with Carbon Source and Biofilm Substratum. Microorganisms 2021, 9, 2168. https://doi.org/10.3390/microorganisms9102168
Yu E-M, Li Z-C, Li Z-F, Wang G-J, Xia Y, Zhang K, Tian J-J, Gong W-B, Xie J. New Insights into the Relationships between Bacterial Dynamics and Water Quality of Aquaculture Systems Supplemented with Carbon Source and Biofilm Substratum. Microorganisms. 2021; 9(10):2168. https://doi.org/10.3390/microorganisms9102168
Chicago/Turabian StyleYu, Er-Meng, Zhen-Chi Li, Zhi-Fei Li, Guang-Jun Wang, Yun Xia, Kai Zhang, Jing-Jing Tian, Wang-Bao Gong, and Jun Xie. 2021. "New Insights into the Relationships between Bacterial Dynamics and Water Quality of Aquaculture Systems Supplemented with Carbon Source and Biofilm Substratum" Microorganisms 9, no. 10: 2168. https://doi.org/10.3390/microorganisms9102168
APA StyleYu, E. -M., Li, Z. -C., Li, Z. -F., Wang, G. -J., Xia, Y., Zhang, K., Tian, J. -J., Gong, W. -B., & Xie, J. (2021). New Insights into the Relationships between Bacterial Dynamics and Water Quality of Aquaculture Systems Supplemented with Carbon Source and Biofilm Substratum. Microorganisms, 9(10), 2168. https://doi.org/10.3390/microorganisms9102168