
Transmission of Escherichia coli from Manure to Root Zones of Field-Grown Lettuce and Leek Plants


Abstract
:1. Introduction
2. Materials and methods
2.1. E. coli Strain 0611 Cultivation, CFU Recovery and DNA Extraction
2.2. Soil Treatments and Experimental Field Design
2.3. Recovery of Escherichia coli 0611 from Manure, Soil and Root Samples
2.4. Rhizosphere Soil Bacterial Community Analysis
2.5. Screening for Strain 0611-Specific Reads in Manure, Bulk and Rhizosphere Soil Metagenomes
2.6. Statistical Analyses
3. Results
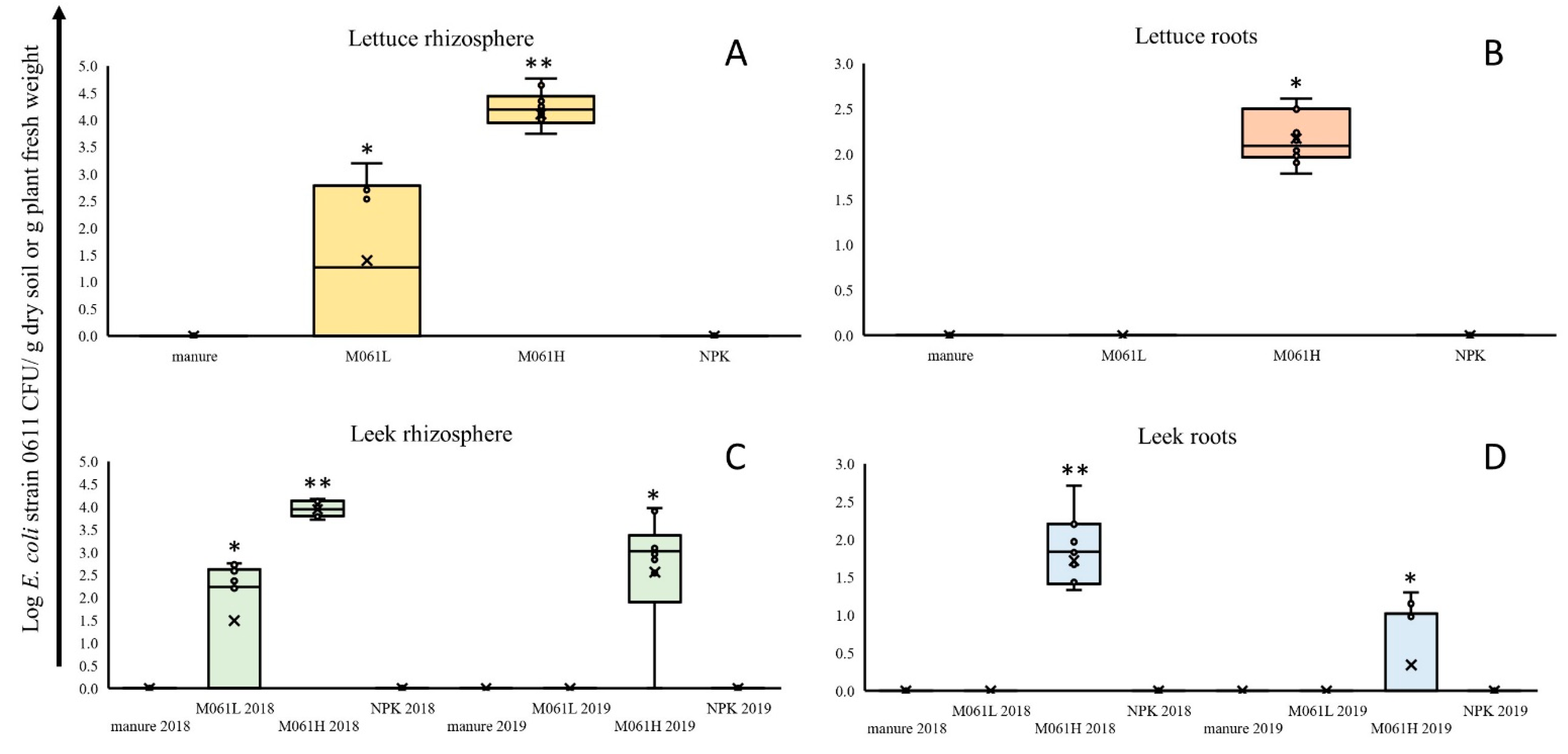
3.1. Lettuce and Leek Plant Growth in the Field and Colonization of Field-Grown Plants by Strain 0611
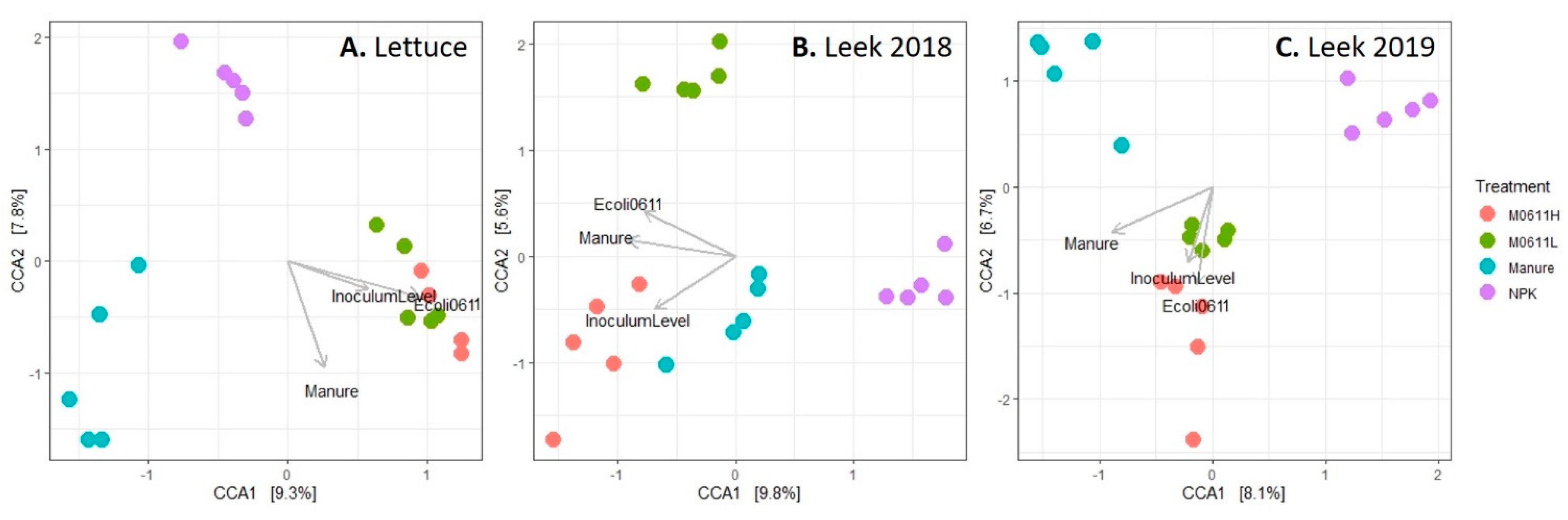
3.2. Impact of Plant and Soil Treatments on the Bacterial Community Composition in Bulk and Rhizosphere Soils
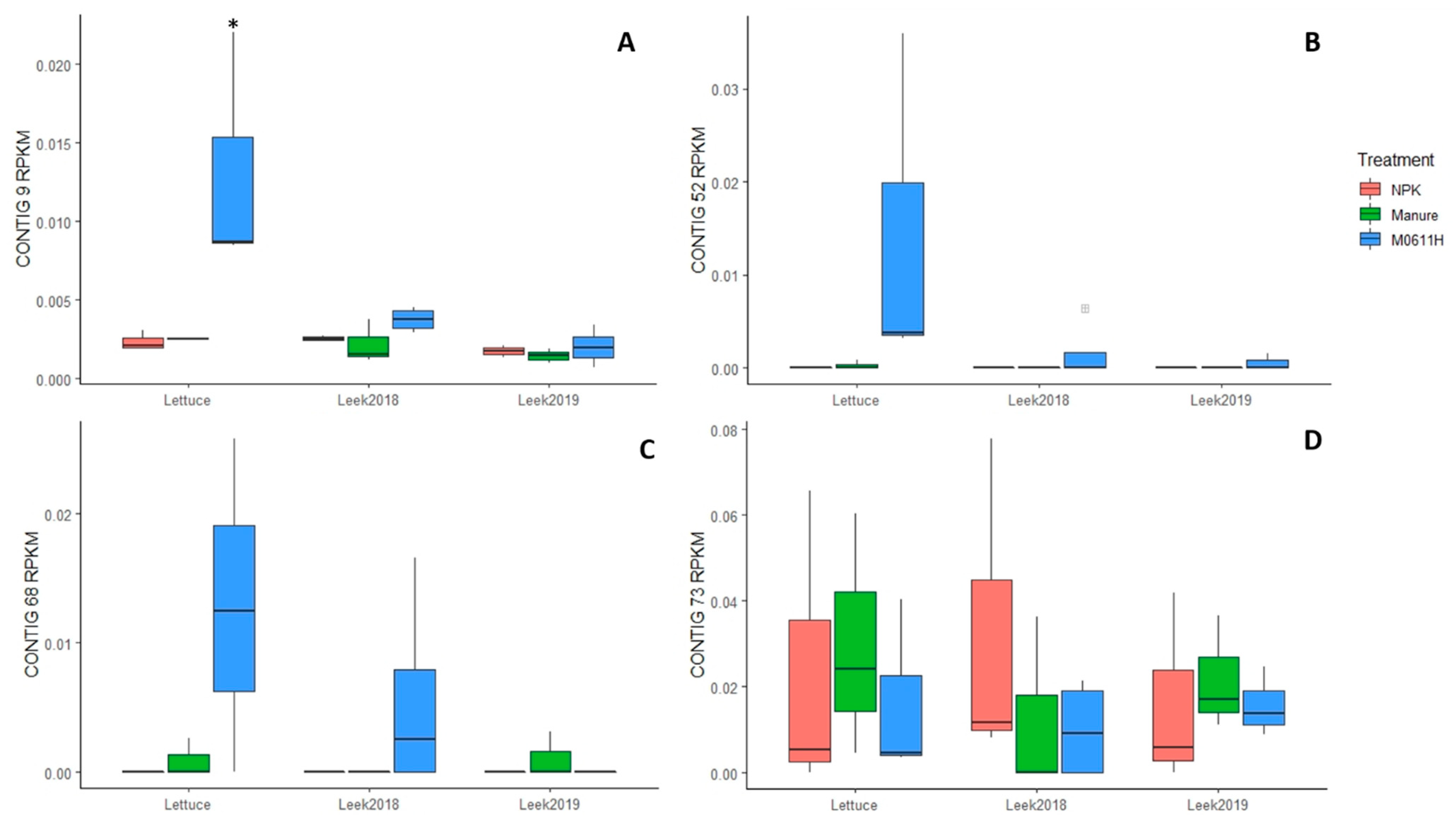
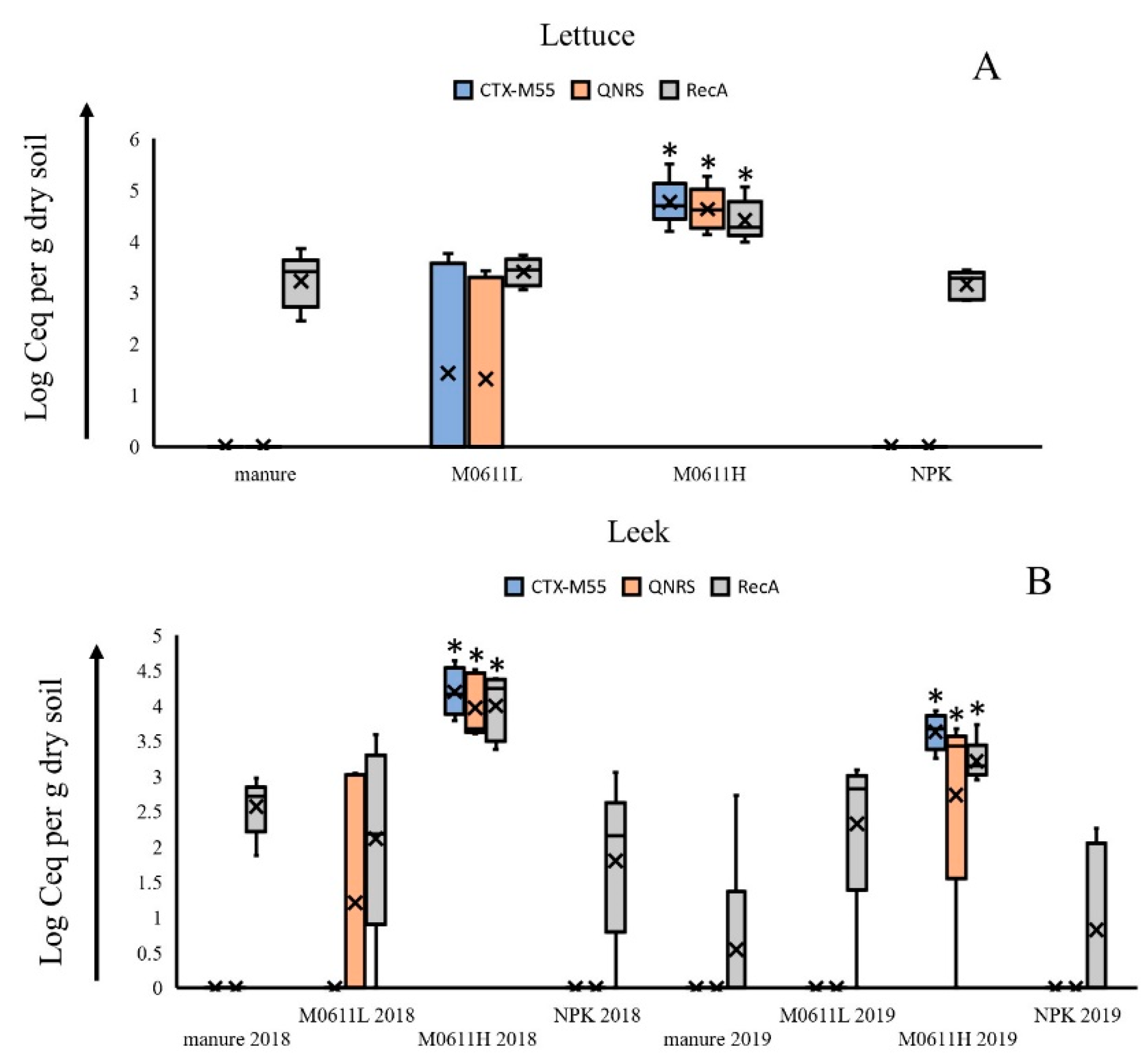
3.3. Rhizosphere Soil Metagenome Screening on Presence of Strain 0611-Specific Reads
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wijnands, L.M.; Delfgou-van Asch, E.H.; Beerepoot-Mensink, M.E.; van der Meij-Florijn, A.; Fitz-James, I.; van Leusden, F.M.; Pielaat, A. Prevalence and concentration of bacterial pathogens in raw produce and minimally processed packaged salads produced in and for the Netherlands. J. Food Prot. 2014, 77, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Beutin, L.; Martin, A. Outbreak of Shiga toxin–producing Escherichia coli (STEC) O104:H4 infection in Germany causes a paradigm shift with regard to human pathogenicity of STEC strains. J. Food Prot. 2012, 75, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Karch, H.; Denamur, E.; Dobrindt, U.; Finlay, B.B.; Hengge, R.; Johannes, L.; Ron, E.Z.; Tønjum, T.; Sansonetti, P.J.; Vicente, M. The enemy within us: Lessons from the 2011 European Escherichia coli O104:H4 outbreak. EMBO Mol. Med. 2012, 4, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Mellmann, A.; Harmsen, D.; Cummings, C.; Zentz, E.B.; Leopold, S.R.; Rico, A. Prospective genomic characterization of the German Enterohemorrhagic Escherichia coli O104:H4 outbreak by rapid next generation sequencing technology. PLoS ONE 2011, 6, e22751. [Google Scholar] [CrossRef]
- Habteselassie, M.Y.; Bischoff, M.; Applegate, B.; Reuhs, B.; Turca, R.F. Understanding the role of agricultural practices in the potential colonization and contamination by Escherichia coli in the rhizospheres of fresh produce. J. Food Prot. 2010, 73, 2001–2009. [Google Scholar] [CrossRef]
- Semenov, A.; van Overbeek, L.; van Bruggen, A. Percolation and survival of Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium in soil amended with contaminated dairy manure or slurry. Appl. Environ. Microbiol. 2009, 75, 3206–3215. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.P.; Avery, L.M.; Killham, K.; Jones, D.L. Survival of Escherichia coli O157:H7 in the rhizosphere of maize grown in waste-amended soil. J. Appl. Microbiol. 2007, 102, 319–326. [Google Scholar] [CrossRef]
- Belias, A.M.; Sbodio, A.; Truchado, P.; Weller, D.; Pinzon, J.; Skots, M.; Allende, A.; Munther, D.; Suslow, T.; Wiedmann, M.; et al. Effect of weather on the die-off of Escherichia coli and attenuated Salmonella enterica serovar Typhimurium on preharvest leafy greens following irrigation with contaminated water. Appl. Environ. Microbiol. 2020, 86, e00899-20. [Google Scholar] [CrossRef] [PubMed]
- Petersen, F.; Hubbart, J.A. Physical factors impacting the survival and occurrence of Escherichia coli in secondary habitats. Water 2020, 12, 1796. [Google Scholar] [CrossRef]
- Chen, Z.; Biswas, S.; Aminabadi, P.; Stackhouse, J.; Jay-Russell, M.; Pandey, P. Prevalence of Escherichia coli O157 and Salmonella spp. in solid bovine manure in California using real-time quantitative PCR. Lett. Appl. Microbiol. 2019, 69, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Van Overbeek, L.S.; van Doorn, J.; Wichers, J.; van Amerongen, A.; van Roermund, H.J.W.; Willemsen, P.T.J. The arable ecosystem as battleground for emergence of new pathogens. Front. Microbiol. 2014, 5, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Bruggen, A.H.C.; Goss, E.M.; Havelaar, A.; van Diepeningen, A.D.; Finckh, M.R.; Morris, J.G., Jr. One Health—Cycling of diverse microbial communities as a connecting force for soil, plant, animal, human and ecosystem health. Sci. Total Environ. 2019, 664, 927–937. [Google Scholar] [CrossRef]
- Hora, R.; Warriner, K.; Shelp, B.J.; Griffiths, M.W. Internalization of Escherichia coli O157:H7 following biological and mechanical disruption of growing spinach plants. J. Food Prot. 2005, 68, 2506–2509. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.M.; Crozier, L.; Marshall, J.; Merget, B.; Holmes, A.; Holden, N.J. Differences in internalization and growth of Escherichia coli O157:H7 within the apoplast of edible plants, spinach and lettuce, compared with the model species Nicotiana benthamiana. Microb. Biotechnol. 2016, 10, 555–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Overbeek, L.S.; Lombaers-van der Plas, C.; van der Zouwen, P. The role of pea (Pisum sativum) seeds in transmission of entero-aggregative Escherichia coli to growing plants. Microorganisms 2020, 8, 1271. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, I.; Cottyn, B.; Uyttendaele, M.; Vlaemynck, G.; Maes, M. Long-term survival of Escherichia coli O157:H7 and Salmonella enterica on butterhead lettuce seeds, and their subsequent survival and growth on the seedlings. Int. J. Food Microbiol. 2013, 161, 214–219. [Google Scholar] [CrossRef]
- Banach, J.L.; van Bokhorst-van de Veen, H.; van Overbeek, L.S.; van der Zouwen, P.S.; van der Fels-Klerx, I. The efficacy of chemical sanitizers on the reduction of Salmonella Typhimurium and Escherichia coli affected by bacterial cell history and water quality. Food Control 2017, 81, 137–146. [Google Scholar] [CrossRef]
- Doan, H.K.; Antequera-Gómez, M.L.; Parikh, A.N.; Leveau, J.H.J. Leaf surface topography contributes to the ability of Escherichia coli on leafy greens to resist removal by washing, escape disinfection with chlorine, and disperse through splash. Front. Microbiol. 2020, 11, 1485. [Google Scholar] [CrossRef]
- Eißenberger, K.; Moench, D.; Drissner, D.; Weiss, A.; Schmidt, H. Adherence factors of enterohemorrhagic Escherichia coli O157:H7 strain Sakai influence its uptake into the roots of Valerianella locusta grown in soil. Food Microbiol. 2018, 76, 245–256. [Google Scholar] [CrossRef]
- Ongeng, D.; Muyanja, C.; Ryckeboer, J.; Geeraerd, A.H.; Springael, D. Rhizosphere effect on survival of Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium in manure-amended soil during cabbage (Brassica oleracea) cultivation under tropical field conditions in Sub-Saharan Africa. Int. J. Food Microbiol. 2011, 149, 133–142. [Google Scholar] [CrossRef]
- Gekenidis, M.T.; Rigotti, S.; Hummerjohann, J.; Walsh, F.; Drissner, D. Long-term persistence of blaCTX-M-15 in soil and lettuce after introducing extended-spectrum β-lactamase (ESBL)-producing Escherichia coli via manure or water. Microorganisms. 2020, 8, 1646. [Google Scholar] [CrossRef] [PubMed]
- Mark Ibekwe, A.; Grieve, C.; Papiernik, S.; Yang, C.-H. Persistence of Escherichia coli O157:H7 on the rhizosphere and phyllosphere of lettuce. Lett. Appl. Microbiol. 2009, 49, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Van Overbeek, L.S.; Wichers, J.H.; van Amerongen, A.; van Roermund, H.J.W.; van der Zouwen, P.; Willemsen, P.T.J. Circulation of Shiga toxin-producing Escherichia coli phylogenetic group B1 strains between calve stable manure and pasture land with grazing heifers. Front. Microbiol. 2020, 11, 1355. [Google Scholar] [CrossRef]
- Jones, D. Potential health risks associated with the persistence of Escherichia coli O157 in agricultural environments. Soil Use and Management 1999, 15, 76–83. [Google Scholar] [CrossRef]
- Brennan, F.; O’flaherty, V.; Kramers, G.; Grant, J.; Richards, K. Long-term persistence and leaching of Escherichia coli in temperate maritime Soils. Appl. Environ. Microbiol. 2009, 76, 1449–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenov, A.V.; van Bruggen, A.H.; van Overbeek, L.; Termorshuizen, A.J.; Semenov, A.M. Influence of temperature fluctuations on Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium in cow manure. FEMS Microbiol. Ecol. 2007, 60, 419–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, J.; Wang, H.; Brookes, P.; Salles, J.; Xu, J. Soil pH and microbial diversity constrain the survival of E.coli in soil. Soil Biol. Biochem. 2018, 128, 139–149. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, H.; Wu, L.; Lou, J.; Wu, J.; Brookes, P.C.; Xu, J. Survival of Escherichia coli O157:H7 in soils from Jiangsu Province, China. PLoS ONE 2013, 8, e81178. [Google Scholar] [CrossRef] [PubMed]
- Van Overbeek, L.S.; Franz, E.; Semenov, A.V.; de Vos, O.J.; van Bruggen, A.H. The effect of the native bacterial community structure on the predictability of E. coli O157:H7 survival in manure-amended soil. Lett. Appl. Microbiol. 2010, 50, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Van Elsas, J.D.; Chiurazzi, M.; Mallon, C.A.; Elhottova, D.; Kristufek, V.; Salles, J.F. Microbial diversity determines the invasion of soil by a bacterial pathogen. Proc. Natl. Acad. Sci. USA 2012, 109, 1159–1164. [Google Scholar] [CrossRef] [Green Version]
- Veldman, K.; Kant, A.; Dierikx, C.; van Essen-Zandbergen, A.; Wit, B.; Mevius, D. Enterobacteriaceae resistant to third-generation cephalosporins and quinolones in fresh culinary herbs imported from Southeast Asia. Int. J. Food Microbiol. 2014, 177, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Herlemann, D.P.R.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.; McMurdie, P.; Holmes, S. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [Green Version]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge—Accurate paired shotgun read merging via overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.B.; Simpson, G.; Solymos, P.; Stevenes, M.H.H.; Wagner, H. Vegan: 2012 Community Ecology Package. R Package Version 2.0-2. Available online: http://CRAN.R-project.org/package=vegan (accessed on 10 September 2021).
- Fagan, P.K.; Hornitzky, M.A.; Bettelheim, K.A.; Djordjevic, S.P. Detection of shiga-like toxin (stx1 and stx2), intimin (eaeA), and enterohemorrhagic Escherichia coli (EHEC) hemolysin (EHEC hlyA) genes in animal feces by multiplex PCR. Appl. Environ. Microbiol. 1999, 65, 868–872. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.; Hur, H.G.; Sadowsky, M.J.; Byappanahalli, M.N.; Yan, T.; Ishii, S. Environmental Escherichia coli: Ecology and public health implications-a review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Van Overbeek, L.S.; Eberl, L.; Givskov, M.; Molin, S.; van Elsas, J.D. Survival of, and induced stress resistance in, carbon-starved Pseudomonas fluorescens cells residing in soil. Appl. Environ. Microbiol. 1995, 61, 4202–4208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerritsen, J.; Hornung, B.; Ritari, J.; Paulin, L.; Rijkers, G.; Schaap, P.; De Vos, W.; Smidt, H. A comparative and functional genomics analysis of the genus Romboutsia provides insight into adaptation to an intestinal lifestyle. BioRxiv 2019, 845511. [Google Scholar] [CrossRef] [Green Version]
- Rieke, E.L.; Soupir, M.L.; Moorman, T.B.; Yang, F.; Howe, A.C. Temporal dynamics of bacterial communities in soil and leachate water after swine manure application. Front. Microbiol. 2018, 9, 3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil/Plant Treatment | Description |
|---|---|
| Soil with manure (Manure) | Soil mixed with non-treated fresh cow manure (3.5% weight basis). |
| Soil with manure and strain 0611 high (M0611H) | Soil mixed with 3.5% fresh cow manure containing Log 7.15 strain 0611 CFU/g manure. Calculated density of strain 0611 in soil was Log 5.70 CFU/g dry soil, which corresponds to Log 8.07 CFU per plant. |
| Soil with manure and strain 0611 low (M0611L) | Soil mixed with 3.5% fresh cow manure containing Log 5.60 strain 0611 CFU/g manure. Calculated density of strain 0611 in soil was Log 4.15 CFU/g dry soil, which corresponds to Log 6.52 CFU per plant |
| Soil with NPK (NPK) | Soil mixed with NPK fertilizer to 0.4 mg per g dry soil, which corresponds to 100 mg per plant. |
| Lettuce | Four soil treatments, 30 plants per treatment. Plants were sampled at 39 days after planting (DAP). |
| Leek 2018 | Four soil treatments, 30 plants per treatment. Plants were sampled at 90 DAP. |
| Leek 2019 | Four soil treatments, 30 plants per treatment. Plants were sampled at 272 DAP. |
| Sample Type | % Escherichia-Shigella Amplicon Reads of Total Reads per Sample (Total Number of Samples with Reads; Total Number of Samples) |
|---|---|
| Manure (non-treated) | 0.069 (1; 1) |
| M0611H | 13.9 (1; 1) |
| M0611L | 0.15 (1; 1) |
| Soil with manure | 0.0029 (3; 3) |
| Soil with M0611H | 0.88 (3; 3) |
| Soil with M0611L | 0.014 (3; 3) |
| Soil with NPK | 0.0027 (1; 3) |
| Lettuce rhizosphere soil with M0611H | 0.034 (4; 5) |
| Leek 2018 rhizosphere soil with M0611H | 0.0064 (3; 5) |
| Contig | Resistance or Plasmid Type | Gene | Accession | Total Length (bp) | Identity (%) |
|---|---|---|---|---|---|
| 9 | Ampicillin, Ciprofloxacin Sulfonamide | blaCTX-M-55 qnrS1 sul2 | GQ456159, AB187515, AY034138 | 876 657 816 | 100 100 100 |
| 52 | Tetracycline | tet(A) | AF534183 | 1275 | 97.8 |
| 65 | Gentamicin | aac(3)-IId | EU022314 | 861 | 99.9 |
| 68 | plasmid | repA (IncY plasmid) | K02380 | 765 | 100 |
| 73 | Trimethoprim | dfrA14 | DQ388123 | 483 | 99.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Overbeek, L.; Duhamel, M.; Aanstoot, S.; van der Plas, C.L.; Nijhuis, E.; Poleij, L.; Russ, L.; van der Zouwen, P.; Andreo-Jimenez, B. Transmission of Escherichia coli from Manure to Root Zones of Field-Grown Lettuce and Leek Plants. Microorganisms 2021, 9, 2289. https://doi.org/10.3390/microorganisms9112289
van Overbeek L, Duhamel M, Aanstoot S, van der Plas CL, Nijhuis E, Poleij L, Russ L, van der Zouwen P, Andreo-Jimenez B. Transmission of Escherichia coli from Manure to Root Zones of Field-Grown Lettuce and Leek Plants. Microorganisms. 2021; 9(11):2289. https://doi.org/10.3390/microorganisms9112289
Chicago/Turabian Stylevan Overbeek, Leo, Marie Duhamel, Stefan Aanstoot, Carin Lombaers van der Plas, Els Nijhuis, Leo Poleij, Lina Russ, Patricia van der Zouwen, and Beatriz Andreo-Jimenez. 2021. "Transmission of Escherichia coli from Manure to Root Zones of Field-Grown Lettuce and Leek Plants" Microorganisms 9, no. 11: 2289. https://doi.org/10.3390/microorganisms9112289
APA Stylevan Overbeek, L., Duhamel, M., Aanstoot, S., van der Plas, C. L., Nijhuis, E., Poleij, L., Russ, L., van der Zouwen, P., & Andreo-Jimenez, B. (2021). Transmission of Escherichia coli from Manure to Root Zones of Field-Grown Lettuce and Leek Plants. Microorganisms, 9(11), 2289. https://doi.org/10.3390/microorganisms9112289