
In Tuber Biocontrol of Potato Late Blight by a Collection of Phenazine-1-Carboxylic Acid-Producing Pseudomonas spp.

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions, and Preparation of the Inocula
2.2. Pathogen Strain, Growth Conditions, and Preparation of the Inoculum
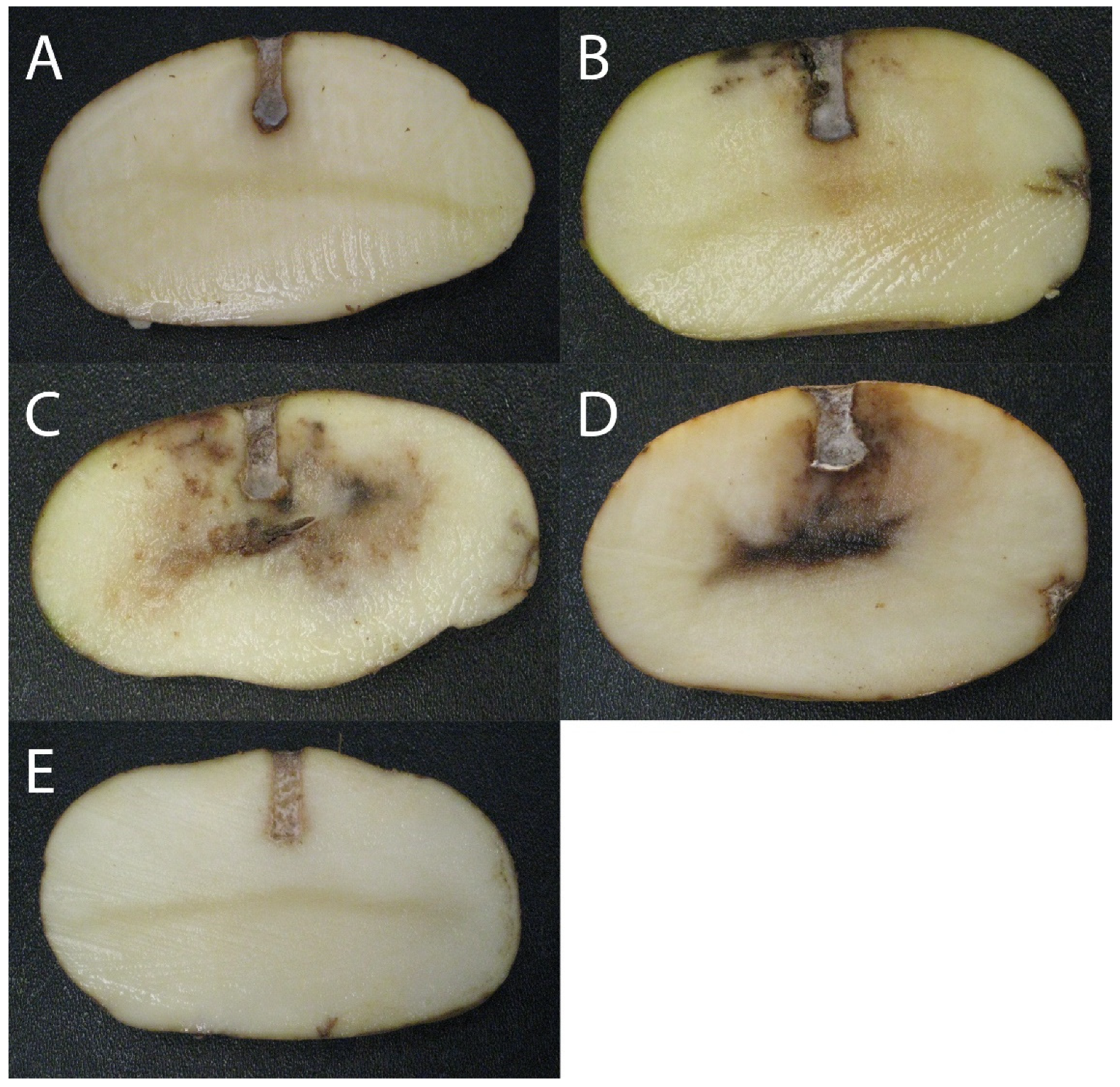
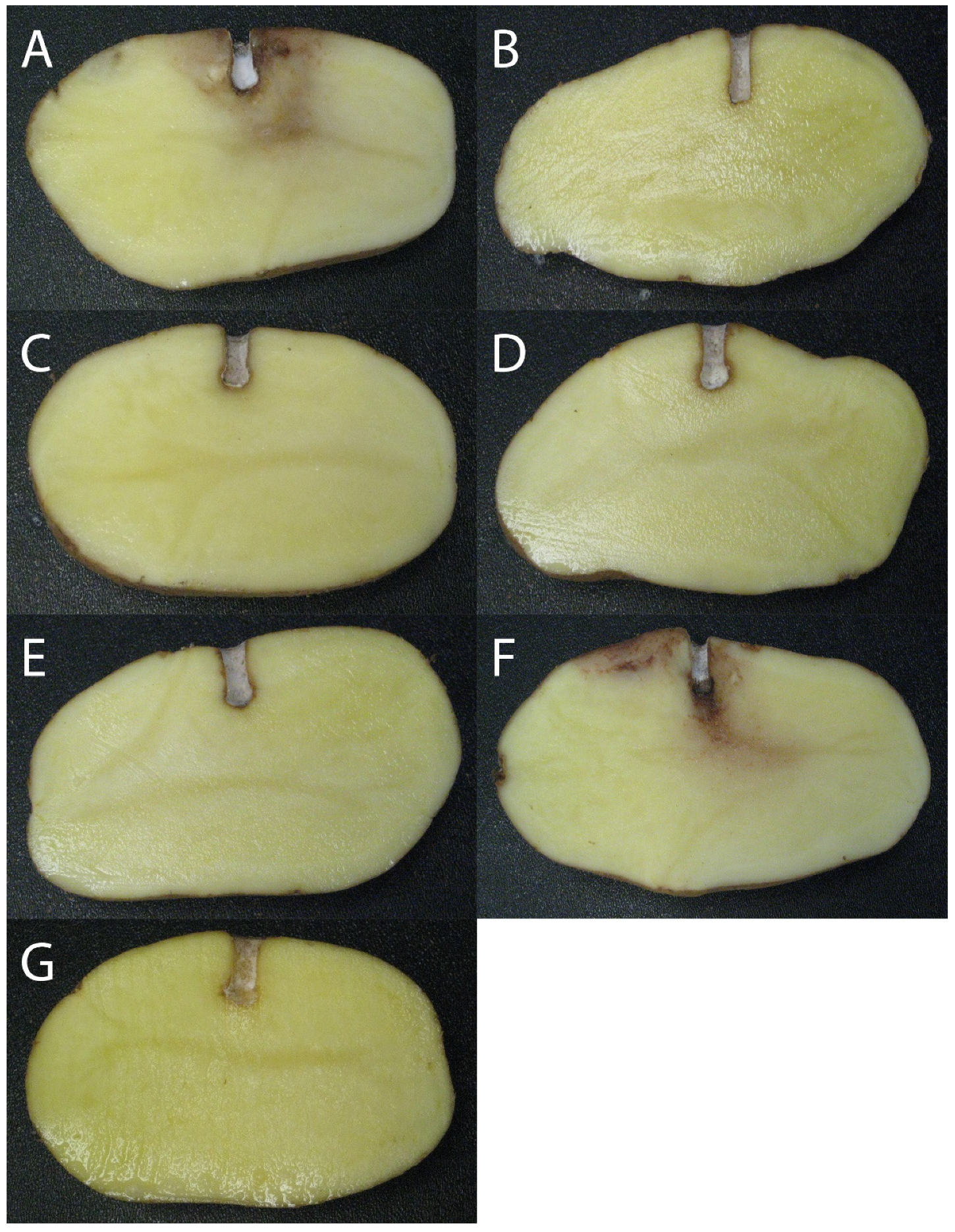
2.3. Potato Tuber Antagonistic Assays with Pseudomonas spp.
2.4. PCA Extractions
2.5. PCA Quantification Using HPLC Analyses
2.6. Potato Tuber Confrontational Assays with Exogenous Applied PCA
2.7. Statistical Analyses
2.8. Identification of Other Genetic Determinants of Biocontrol Interest
3. Results
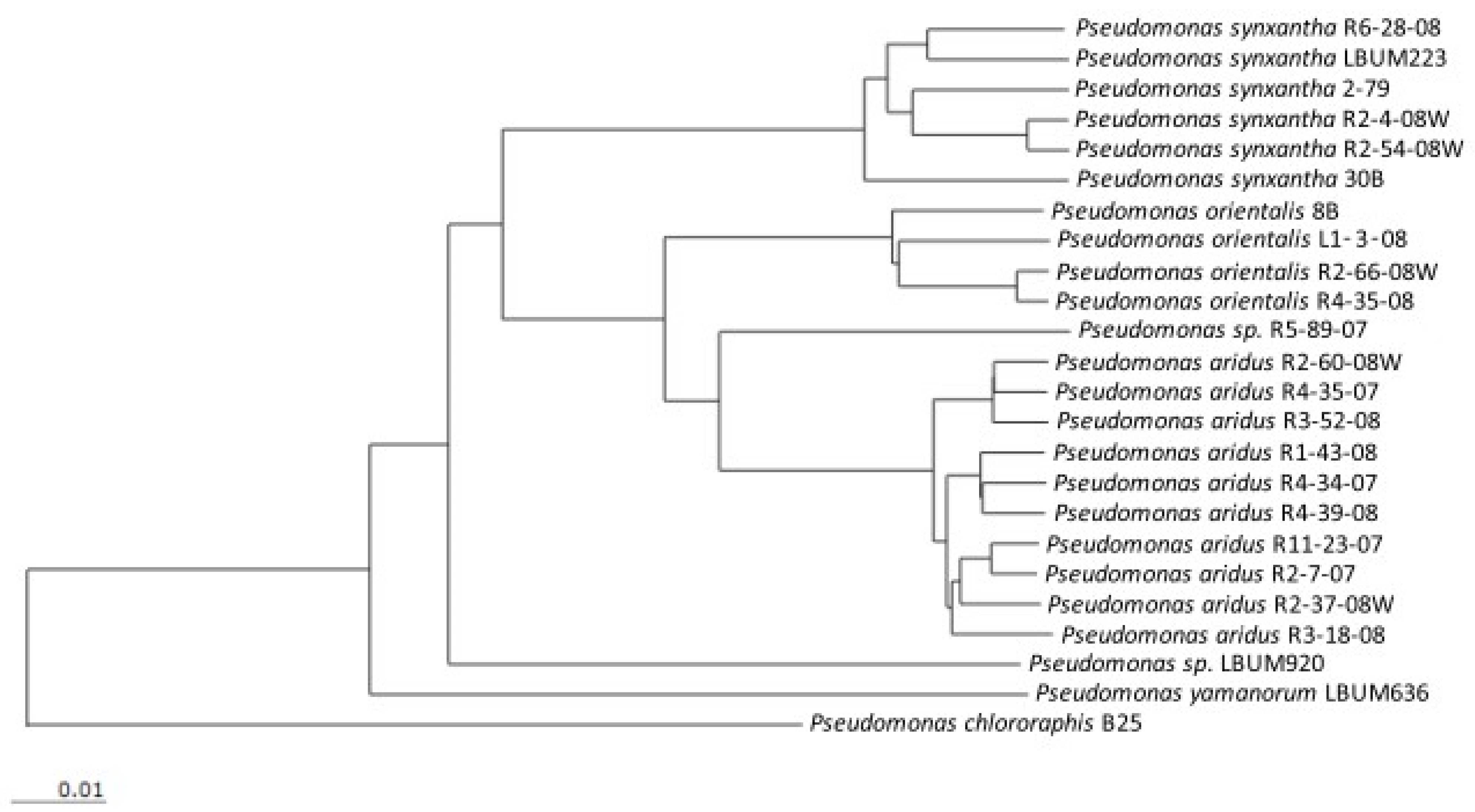
3.1. Potato Tuber Antagonistic Assays
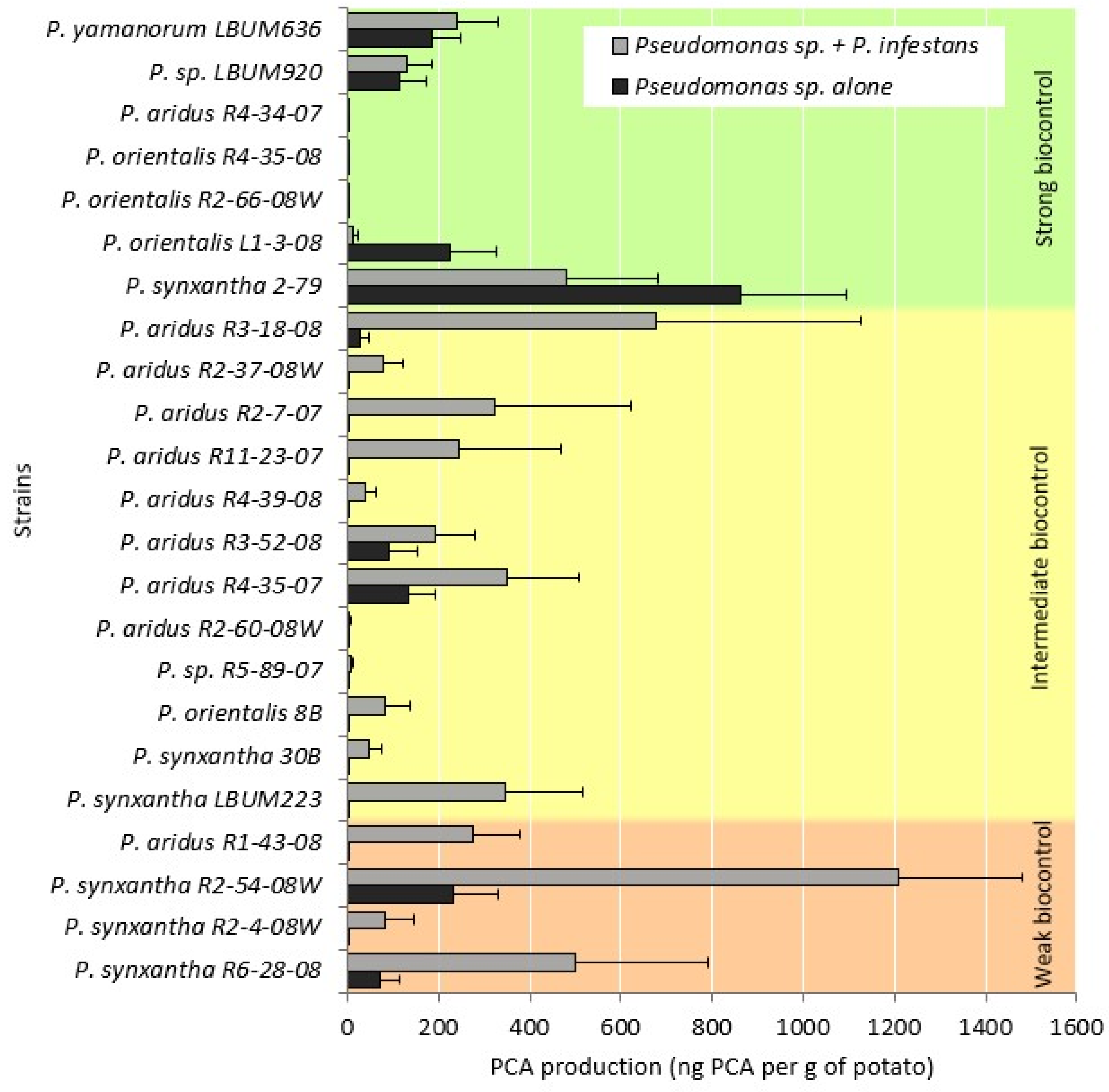
3.2. PCA Quantification Using HPLC
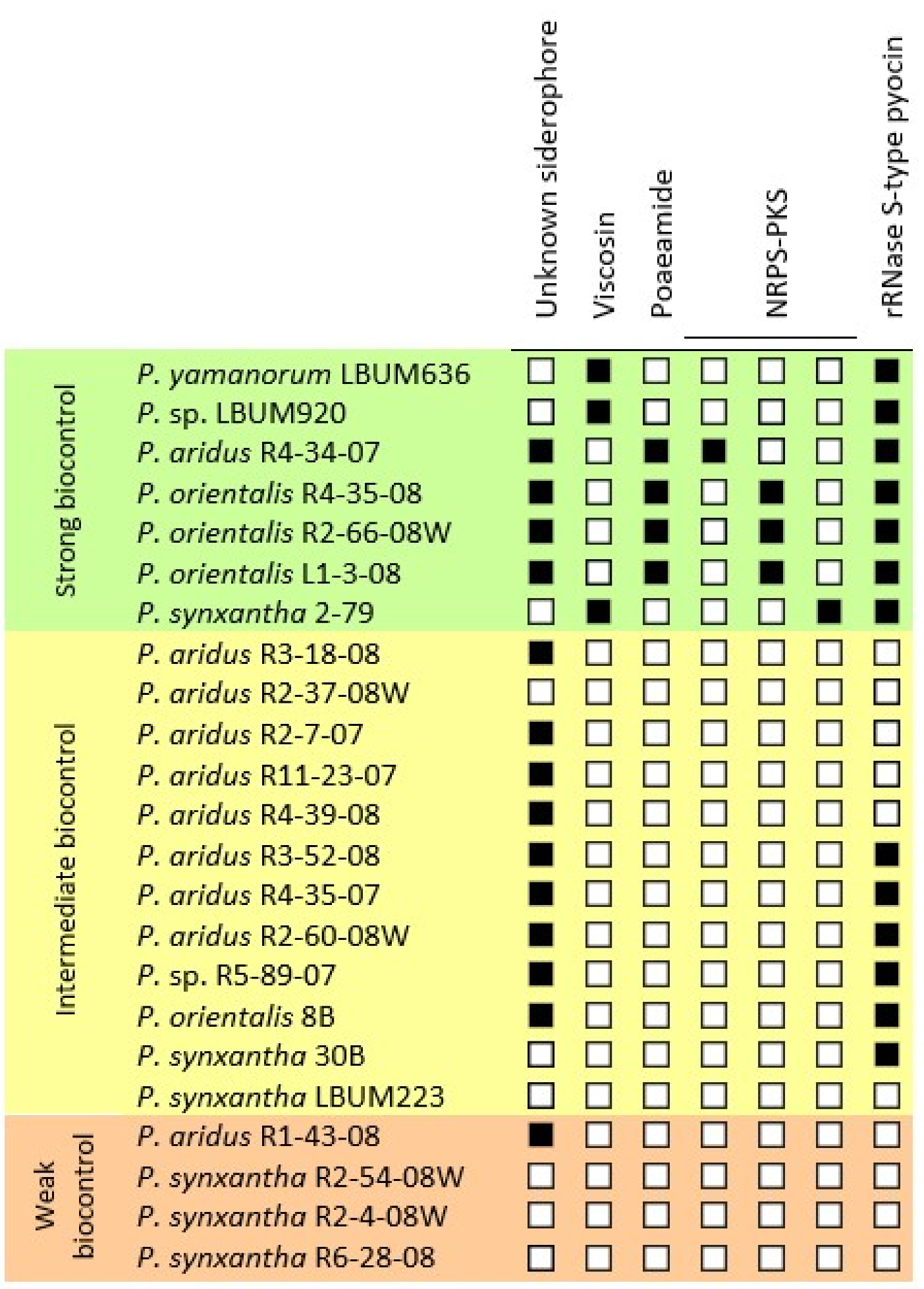
3.3. Identification of Other Genetic Determinants of Biocontrol Interest
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leesutthiphonchai, W.; Vu, A.L.; Ah-Fong, A.M.V.; Judelson, H.S. How does Phytophthora infestans evade control efforts? Modern insight into the late blight disease. Phytopathology 2018, 108, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Late Blight of Tomato and Potato. Available online: https://vtechworks.lib.vt.edu/handle/10919/87900 (accessed on 10 November 2021).
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five Reasons to consider Phytophthora infestans a reemerging pathogen. Phytopathology 2015, 105, 966–981. [Google Scholar] [CrossRef] [Green Version]
- González-Tobón, J.; Childers, R.; Olave, C.; Regnier, M.; Rodríguez-Jaramillo, A.; Fry, W.; Restrepo, S.; Danies, G. Is the phenomenon of mefenoxam-acquired resistance in Phytophthora infestans universal? Plant Dis. 2020, 104, 211–221. [Google Scholar] [CrossRef]
- Characterization and Tracking of Strains of the Potato Blight Pathogen in Canada–Final Progress Report March 2018. Available online: https://hortcouncil.ca/resources/publications/#late-blight (accessed on 10 November 2021).
- Peters, R.D.; Al-Mughrabi, K.L.; Kalischulk, M.L.; Dobinson, K.F.; Conn, K.L.; Alker, H.; Islam, M.R.; Daayf, F.; Lynn, J.; Bizimungu, B.; et al. Characterization of Phytophthora infestans population diversity in Canada reveals increased migration and genotype recombination. Can. J. Plant Pathol. 2014, 36, 73–82. [Google Scholar] [CrossRef]
- Siddiqui, Z.A. PGPR: Prospective Biocontrol Agents of Plant Pathogens. In PGPR: Biocontrol and Biofertilization; Siddiqui, Z.A., Ed.; Springer: Berlin, Germany, 2006; pp. 111–142. [Google Scholar]
- Kloepper, J.W.; Rodríguez-Kábana, R.; Zehnder, G.W.; Murphy, J.F.; Sikora, E.; Fernández, C.F. Plant root-bacterial interactions in biological control of soilborne diseases and potential extension to systemic and foliar disease. Australas. Plant Pathol. 1999, 28, 21–26. [Google Scholar] [CrossRef]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biessy, A.; Filion, M. Phenazines in plant-beneficial Pseudomonas spp.: Biosynthesis, regulation, function and genomics. Environ. Microbiol. 2018, 20, 3905–3917. [Google Scholar] [CrossRef] [Green Version]
- Chin-A-Woeng, T.F.C.; Bloemberg, G.V.; Lugtenberg, B.J. Phenazines and their role in biocontrol by Pseudomonas bacteria. New Phytol. 2003, 157, 503–523. [Google Scholar] [CrossRef] [Green Version]
- Mavrodi, D.V.; Blankenfeldt, W.; Thomashow, L.S. Phenazine compounds in fluorescent Pseudomonas spp. biosynthesis and regulation. Annu. Rev. Phytopathol. 2006, 44, 417–445. [Google Scholar] [CrossRef]
- Xu, S.; Pan, X.; Luo, J.; Wu, J.; Zhou, Z.; Liang, X.; He, Y.; Zhou, M. Effects of phenazine-1-carboxylic acid on the biology of the plant-pathogenic bacterium Xanthomonas oryzae pv. oryzae. Pestic. Biochem. Physiol. 2015, 117, 39–46. [Google Scholar] [CrossRef]
- Mazzola, M.; Cook, R.J.; Thomashow, L.S.; Weller, D.M.; Pierson, L.S., III. Contribution of phenazine antibiotic biosynthesis to the ecological competence of fluorescent Pseudomonads in soil habitats. Appl. Environ. Microbiol. 1992, 58, 2616–2624. [Google Scholar] [CrossRef] [Green Version]
- Delaney, S.M.; Mavrodi, D.V.; Bonsall, R.F.; Thomashow, L.S. phzO, a gene for biosynthesis of 2-hydroxylated phenazine compounds in Pseudomonas aureofaciens 30–84. J. Bacteriol. 2001, 183, 318–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomashow, L.S.; Weller, D.M. Role of a phenazine antibiotic from Pseudomonas fluorescens in biological control of Gaeumannomyces graminis var. tritici. J. Bacteriol. 1988, 170, 3499–3508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Goa, Q.; Hamada, M.S.; Dawood, D.H.; Zheng, J.; Chen, Y.; Ma, Z. Potential of Pseudomonas chlororaphis subsp. aurantiaca Strain Pcho10 as a Biocontrol Agent Against Furasium graminearum. Phytopathology 2014, 104, 1289–1297. [Google Scholar]
- Arseneault, T.; Goyer, C.; Filion, M. Phenazine Production by Pseudomonas sp. LBUM223 Contributes to the biological control of potato common scab. Phytopathology 2013, 103, 995–1000. [Google Scholar] [CrossRef] [Green Version]
- Arseneault, T.; Goyer, C.; Filion, M. Pseudomonas fluorescens LBUM223 Increases potato yield and reduces common scab symptoms in the field. Phytopathology 2015, 105, 1311–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, C.K.; Arseneault, T.; Novinscak, A.; Filion, M. Phenazine-1-carboxylic acid production by Pseudomonas fluorescens LBUM636 alters Phytophthora infestans growth and late blight development. Phytopathology 2017, 107, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Roquigny, R.; Novinscak, A.; Arseneault, T.; Joly, D.L.; Filion, M. Transcriptome alteration in Phytophthora infestans in response to phenazine-1-carboxylic acid production by Pseudomonas fluorescens strain LBUM223. BMC Genom. 2018, 19, 474. [Google Scholar] [CrossRef]
- De Vrieze, M.; Gloor, R.; Massada Codina, J.; Torriani, S.; Gindro, K.; L’Haridon, F.; Bailly, A.; Weisskopf, L. Biocontrol activity of three Pseudomonas on a newly assembled collection of Phytophthora infestans isolates. Phytopathology 2019, 109, 1555–1565. [Google Scholar] [CrossRef] [Green Version]
- Biessy, A.; Novinscak, A.; Blom, J.; Léger, G.; Thomashow, L.S.; Cazorla, F.M.; Josic, D.; Filion, M. Diversity of phytobeneficial traits revealed by whole-genome analysis of worldwide-isolated phenazine-producing Pseudomonas spp. Environ. Microbiol. 2019, 21, 437–455. [Google Scholar]
- Biessy, A.; Novinscak, A.; St-Onge, R.; Léger, G.; Zboralski, A.; Filion, M. Inhibition of three potato pathogens by phenazine-producing Pseudomonas spp. is associated with multiple biocontrol-related traits. mSphere 2021, 6, e00427-21. [Google Scholar] [CrossRef]
- Blom, J.; Kreis, J.; Spänig, S.; Juhre, T.; Bertelli, C.; Ernst, C.; Goesmann, A. EDGAR 2.0: An enhanced software platform for comparative gene content analyses. Nucleic Acids Res. 2016, 44, W22–W28. [Google Scholar] [CrossRef] [Green Version]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar]
- Parejko, J.A.; Mavrodi, D.V.; Mavrodi, O.V.; Weller, D.M.; Thomashow, L.S. Population Structure and Diversity of Phenazine-1-Carboxylic Acid Producing Fluorescent Pseudomonas spp. from Dryland Cereal Fields of Central Washington State (USA). Microb. Ecol. 2012, 64, 226–241. [Google Scholar] [CrossRef] [PubMed]
- Roquigny, R.; Arseneault, T.; Gadkar, V.J.; Novinscak, A.; Joly, D.L.; Filion, M. Complete genome sequence of biocontrol strain Pseudomonas fluorescens LBUM223. Genome Announc. 2015, 3, e00443-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, D.M.; Cook, R.J. Suppression of Take-All of Wheat by Seed Treatments with Fluorescent Pseudomonads. Phytopathology 1983, 73, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Shirzad, A.; Fallahzadeh-Mamaghani, V.; Pazhouhandeh, M. Antagonistic Potential of Fluorescent Pseudomonads and Control of Crown and Root Rot of Cucumber Caused by Phythophtora drechsleri. Plant Pathol. J. 2012, 28, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mavrodi, D.V.; Peever, T.L.; Mavrodi, O.V.; Parejko, J.A.; Raaijmakers, J.M.; Lemanceau, P.; Mazurier, S.; Heide, L.; Blankenfeldt, W.; Weller, D.M.; et al. Diversity and Evolution of the Phenazine Biosynthesis Pathway. Appl. Environ. Microbiol. 2010, 76, 866–879. [Google Scholar] [CrossRef] [Green Version]
- Morrison, C.K.; Novinscak, A.; Gadkar, V.J.; Joly, D.L.; Filion, M. Complete genome sequence of Pseudomonas fluorescens LBUM636, a strain with biocontrol capabilities against late blight of potato. Genome Announc. 2016, 4, e00446-16. [Google Scholar] [CrossRef] [Green Version]
- Gamboa, S.; Perez, W.; Andrade-Piedra, J.L.; Forbes, G. Laboratory Manual for Phytophthora Infestans Work at CIP; International Potato Center: Lima, Peru, 2019; p. 11. [Google Scholar]
- Brazinskiene, V.; Asakaviciute, R.; Razukas, A.; Ivanauskas, L. Quantification of biologically active compounds in the tubers of potato varieties of different maturity. Zemdirb. Agric. 2017, 104, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; de Los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. antiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.-Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M. InterPro in 2017—beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef]
- De Vrieze, M.; Varadarajan, A.R.; Schneeberger, K.; Bailly, A.; Rohr, R.P.; Ahrens, C.H.; Weisskopf, L. Linking Comparative Genomics of Nine Potato-Associated Pseudomonas Isolates with Their Differing Biocontrol Potential Against Late Blight. Front. Microbiol. 2020, 11, 857. [Google Scholar] [CrossRef] [PubMed]
- Zboralski, A.; Biessy, A.; Savoie, M.-C.; Novinscak, A.; Filion, M. Metabolic and genomic traits of phytobeneficial phenazine-producing Pseudomonas spp. are linked to rhizosphere colonization in Arabidopsis thaliana and Solanum tuberosum. Appl. Environ. Microbiol. 2020, 86, e02443-19. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, D.J.; O’Gara, F. Traits of fluorescent Pseudomonas spp. involved in suppression of plant root pathogens. Microbiol. Rev. 1992, 56, 662–676. [Google Scholar] [CrossRef] [PubMed]
- Loper, J.E.; Buyer, J.S. Siderophores in Microbial Interactions on Plant Surfaces. Mol. Plant-Microbe Interact. 1991, 4, 5–13. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; de Brujin, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [Green Version]
- Thrane, C.; Nielsen, T.H.; Nielsen, M.N.; Sørensen, J.; Olsson, S. Viscosinamide-producing Pseudomonas fluorescens DR54 exerts a biocontrol effect on Pythium ultimum in sugar beet rhizosphere. FEMS Microbiol. Ecol. 2000, 33, 139–146. [Google Scholar] [CrossRef]
- Zachow, C.; Jahanshah, G.; de Bruijn, I.; Song, C.; Ianni, F.; Pataj, Z.; Gerhardt, H.; Pianet, I.; Lämmerhofer, M.; Berg, G.; et al. The Novel Lipopeptide Poaeamide of the Endophyte Pseudomonas poae RE*1-1-14 Is Involved in Pathogen Suppression and Root Colonization. Mol. Plant-Microbe Interact. 2015, 28, 800–810. [Google Scholar] [CrossRef] [Green Version]
- Gross, H.; Loper, J.E. Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep. 2019, 26, 1408–1446. [Google Scholar] [CrossRef] [PubMed]
- Ghequire, M.G.; De Mot, R. Ribosomally encoded antibacterial proteins and peptides from Pseudomonas. FEMS Microbiol. Rev. 2014, 38, 523–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W., II; Lim, C.K.; Shaffer, B.T.; Elbourne, L.D.H.; Stockwell, V.O.; Hartney, S.L.; Breakwell, K.; et al. Comparative genomics of plant-associated Pseudomonas spp.: Insights into diversity and inheritance of traits involved in multitrophic interactions. PLoS Genet 2012, 8, e1002784. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin | Genome Accession Number | Reference or Source |
|---|---|---|---|
| P. synxantha R6-28-08 | Wheat, USA | CP027756 | Parejko et al. [27] |
| P. synxantha LBUM223 | Strawberry, Canada | CP011117 | Roquigny et al. [28] |
| P. synxantha 2-79 | Wheat, USA | CP027755 | Weller & Cook [29] |
| P. synxantha R2-4-08W | Wheat, USA | CP027757 | Parejko et al. [27] |
| P. synxantha R2-54-08W | Wheat, USA | CP027758 | Parejko et al. [27] |
| P. synxantha 30B | Wheat, Iran | CP027754 | Shirzad et al. [30] |
| P. orientalis 8B | Wheat, Iran | CP027723 | Shirzad et al. [30] |
| P. orientalis L1-3-08 | Wheat, USA | CP027724 | Parejko et al. [27] |
| P. orientalis R2-66-08W | Wheat, USA | CP027725 | Parejko et al. [27] |
| P. orientalis R4-35-08 | Wheat, USA | CP027726 | Parejko et al. [27] |
| P. sp. R5-89-07 | Wheat, USA | CP027727 | Parejko et al. [27] |
| P. aridus R2-60-08W | Wheat, USA | CP027731 | Parejko et al. [27] |
| P. aridus R4-35-07 | Wheat, USA | CP027732 | Mavrodi et al. [31] |
| P. aridus R3-52-08 | Wheat, USA | CP027730 | Parejko et al. [27] |
| P. aridus R1-43-08 | Wheat, USA | CP027734 | Parejko et al. [27] |
| P. aridus R4-34-07 | Wheat, USA | CP027760 | Mavrodi et al. [31] |
| P. aridus R4-39-08 | Wheat, USA | CP027733 | Parejko et al. [27] |
| P. aridus R11-23-07 | Wheat, USA | CP027761 | Mavrodi et al. [31] |
| P. aridus R2-7-07 | Wheat, USA | CP027759 | Mavrodi et al. [31] |
| P. aridus R2-37-08W | Wheat, USA | CP027728 | Parejko et al. [27] |
| P. aridus R3-18-08 | Wheat, USA | CP027729 | Parejko et al. [27] |
| P. sp. LBUM920 | Spruce, Canada | CP027762 | Richard Hamelin |
| P. yamanorum LBUM636 | Strawberry, Canada | CP012400 | Morrison et al. [32] |
| Biocontrol Activity | ||
|---|---|---|
| Weak | Intermediate | Strong |
| P. aridus R1-43-08 P. synxantha R2-54-08W P. synxantha R2-4-08W P. synxantha R6-28-08 | P. aridus R3-18-08 P. aridus R2-37-08W P. aridus R2-7-07 P. aridus R11-23-07 P. aridus R4-39-08 P. aridus R3-S2-08 P. aridus R2-60-08W P. sp. R5-89-07 P. orientalis 8B P. synxantha 30B P. synxantha LBUM223 | P. yamanorum LBUM636 P. sp. LBUM920 P. aridus R4-34-07 P. orientalis R4-35-08 P. orientalis R2-66-08W P. orientalis L1-3-08 P. synxantha 2-79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Léger, G.; Novinscak, A.; Biessy, A.; Lamarre, S.; Filion, M. In Tuber Biocontrol of Potato Late Blight by a Collection of Phenazine-1-Carboxylic Acid-Producing Pseudomonas spp. Microorganisms 2021, 9, 2525. https://doi.org/10.3390/microorganisms9122525
Léger G, Novinscak A, Biessy A, Lamarre S, Filion M. In Tuber Biocontrol of Potato Late Blight by a Collection of Phenazine-1-Carboxylic Acid-Producing Pseudomonas spp. Microorganisms. 2021; 9(12):2525. https://doi.org/10.3390/microorganisms9122525
Chicago/Turabian StyleLéger, Geneviève, Amy Novinscak, Adrien Biessy, Simon Lamarre, and Martin Filion. 2021. "In Tuber Biocontrol of Potato Late Blight by a Collection of Phenazine-1-Carboxylic Acid-Producing Pseudomonas spp." Microorganisms 9, no. 12: 2525. https://doi.org/10.3390/microorganisms9122525
APA StyleLéger, G., Novinscak, A., Biessy, A., Lamarre, S., & Filion, M. (2021). In Tuber Biocontrol of Potato Late Blight by a Collection of Phenazine-1-Carboxylic Acid-Producing Pseudomonas spp. Microorganisms, 9(12), 2525. https://doi.org/10.3390/microorganisms9122525