
Gut Microbiota Profile and Changes in Body Weight in Elderly Subjects with Overweight/Obesity and Metabolic Syndrome

 , , ,
, , ,  , , ,
, , ,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Study Design
2.2. General Assessments, Anthropometric and Biochemical Measurements, Samples Collection
2.3. Microbial DNA Extraction, 16S Amplicon Sequencing and Data Processing
2.4. Statistical Analysis
3. Results
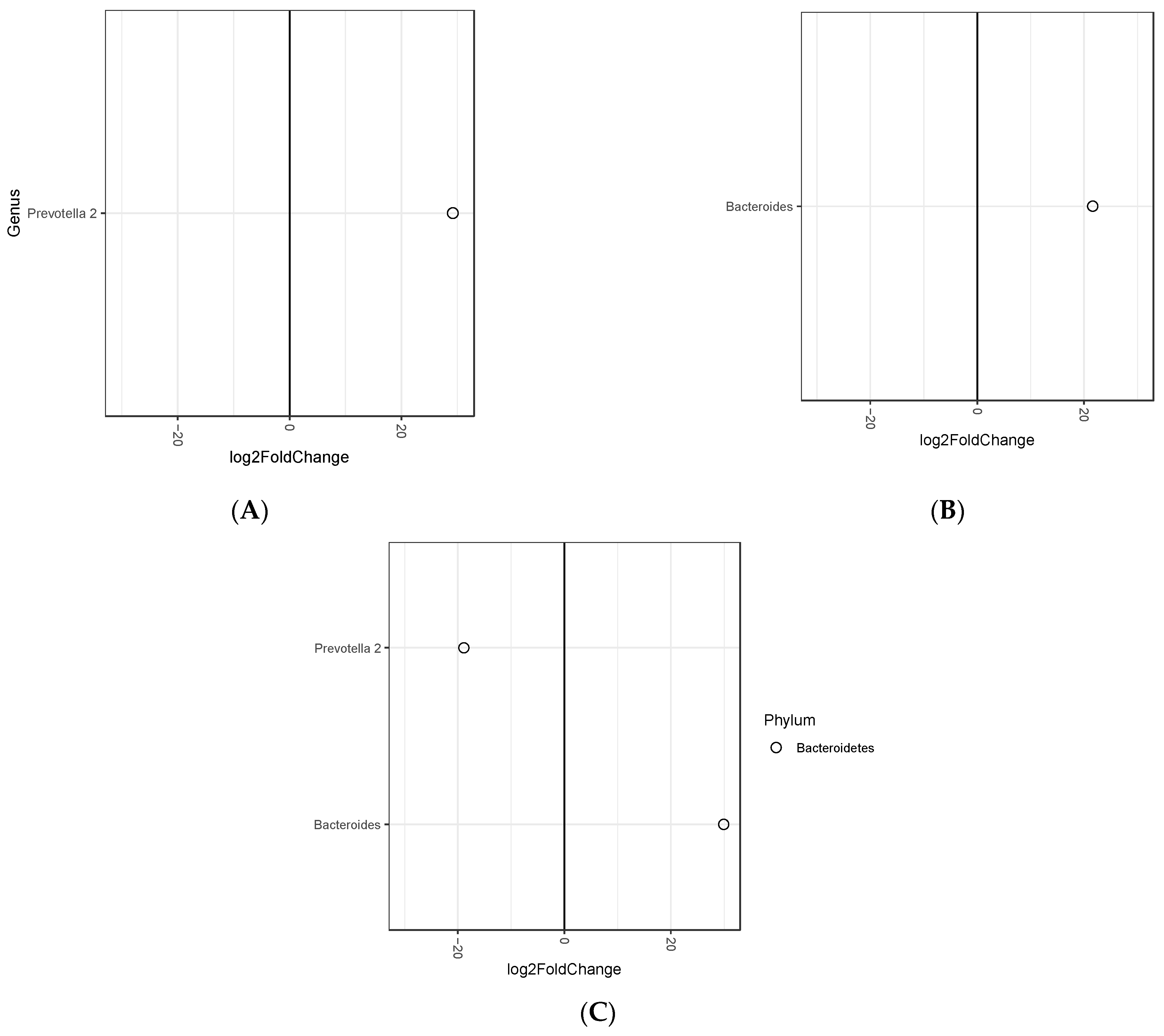
3.1. Association between Fecal Microbiota and Tertiles of Baseline Body Mass Index
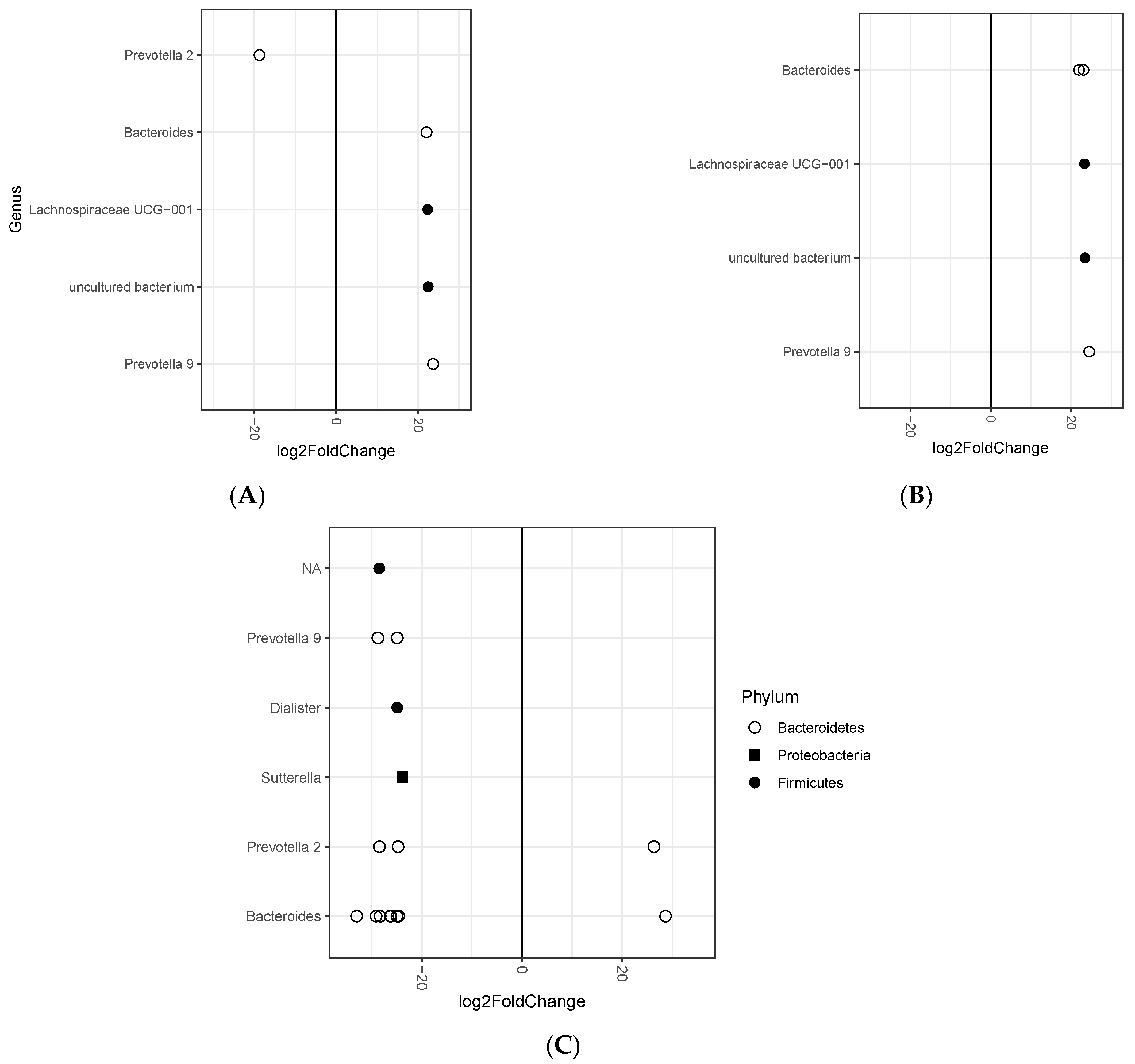
3.2. Association between Fecal Microbiota and Tertiles of Changes in Body Weight after 12-Month Intervention
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Beydoun, M.A. The obesity epidemic in the United States—Gender, age, socioeconomic, racial/ethnic, and geographic characteristics: A systematic review and meta-regression analysis. Epidemiol. Rev. 2007, 29, 6–28. [Google Scholar] [CrossRef]
- Ghosh, S.; Bouchard, C. Convergence between biological, behavioural and genetic determinants of obesity. Nat. Rev. Genet. 2017, 18, 731. [Google Scholar] [CrossRef] [PubMed]
- Martin-Rodriguez, E.; Guillen-Grima, F.; Martí, A.; Brugos-Larumbe, A. Comorbidity associated with obesity in a large population: The APNA study. Obes. Res. Clin. Pract. 2015, 9, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [Green Version]
- Thaiss, C.A.; Itav, S.; Rothschild, D.; Meijer, M.T.; Levy, M.; Moresi, C.; Dohnalová, L.; Braverman, S.; Rozin, S.; Malitsky, S.; et al. Persistent microbiome alterations modulate the rate of post-dieting weight regain. Nature 2016, 540, 544–551. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.; Roager, H.M.; Astrup, A.; Hjorth, M.F. Microbial enterotypes in personalized nutrition and obesity management. Am. J. Clin. Nutr. 2018, 108, 645–651. [Google Scholar] [CrossRef]
- Gardner, C.D.; Trepanowski, J.F.; Del Gobbo, L.C.; Hauser, M.E.; Rigdon, J.; Ioannidis, J.P.; Desai, M.; King, A.C. Effect of low-fat VS low-carbohydrate diet on 12-month weight loss in overweight adults and the association with genotype pattern or insulin secretion the DIETFITS randomized clinical trial. JAMA 2018, 319, 667–679. [Google Scholar] [CrossRef]
- Pedrogo, D.A.M.; Jensen, M.D.; Van Dyke, C.T.; Murray, J.A.; Woods, J.A.; Chen, J.; Kashyap, P.C.; Nehra, V. Gut Microbial Carbohydrate Metabolism Hinders Weight Loss in Overweight Adults Undergoing Lifestyle Intervention with a Volumetric Diet. Mayo Clin. Proc. 2018, 93, 1101–1110. [Google Scholar]
- Hjorth, M.F.; Roager, H.M.; Larsen, T.M.; Poulsen, S.K.; Licht, T.R.; Bahl, M.I.; Zohar, Y.; Astrup, A. Pre-treatment microbial Prevotella-to-Bacteroides ratio, determines body fat loss success during a 6-month randomized controlled diet intervention. Int. J. Obes. 2018, 42, 580–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.T.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the Metabolic Syndrome A Joint Interim Statement of the International Diabetes Federation Task Force on Epidemiology and Prevention for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mas-Lloret, J.; Obón-Santacana, M.; Ibáñez-Sanz, G.; Guinó, E.; Pato, M.L.; Rodriguez-Moranta, F.; Mata, A.; García-Rodríguez, A.; Moreno, V.; Pimenoff, V.N. Gut microbiome diversity detected by high-coverage 16S and shotgun sequencing of matched stool and colon biopsy samples. bioRxiv 2019, 742635. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods. 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, 590–596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.F.; Cotter, P.D.; Healy, S.; Marques, T.M.; O’sullivan, O.; Fouhy, F.; Clarke, S.F.; O’toole, P.W.; Quigley, E.M.; Stanton, C.; et al. Composition and energy harvesting capacity of the gut microbiota: Relationship to diet, obesity and time in mouse models. Gut 2010, 59, 1635–1642. [Google Scholar] [CrossRef]
- Duncan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human colonic microbiota associated with diet, obesity and weight loss. Int. J. Obes. 2008, 32, 1720–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.P.; Dhotre, D.P.; Chavan, S.G.; Sultan, A.; Jain, D.S.; Lanjekar, V.B.; Gangawani, J.; Shah, P.S.; Todkar, J.S.; Shah, S.; et al. Molecular analysis of gut microbiota in obesity among Indian individuals. J. Biosci. 2012, 37, 647–657. [Google Scholar] [CrossRef]
- John, G.K.; Mullin, G.E. The Gut Microbiome and Obesity. Curr. Oncol. Rep. 2016, 18, 1–7. [Google Scholar] [CrossRef]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of the gut microbiota composition between obese and non-obese individuals in a Japanese population, as analyzed by terminal restriction fragment length polymorphism and next-generation sequencing. BMC Gastroenterol. 2015, 15, 100. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef]
- Wexler, A.G.; Goodman, A.L. An insider’s perspective: Bacteroides as a window into the microbiome. Nat. Microbiol. 2017, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.D.; Wing, R.; Rittmann, B.E.; et al. Human gut microbiota in obesity and after gastric bypass. Proc. Natl. Acad. Sci. USA 2009, 106, 2365–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zupancic, M.L.; Cantarel, B.L.; Liu, Z.; Drabek, E.F.; Ryan, K.A.; Cirimotich, S.; Jones, C.; Knight, R.; Walters, W.A.; Knights, D.; et al. Analysis of the gut microbiota in the old order amish and its relation to the metabolic syndrome. PLoS ONE 2012, 7, e43052. [Google Scholar] [CrossRef]
- Hjorth, M.F.; Christensen, L.; Kjølbæk, L.; Larsen, L.H.; Roager, H.M.; Kiilerich, P.; Kristiansen, K.; Astrup, A. Pretreatment Prevotella-to-Bacteroides ratio and markers of glucose metabolism as prognostic markers for dietary weight loss maintenance. Eur. J. Clin. Nutr. 2020, 74, 338–347. [Google Scholar] [CrossRef]
- Christensen, L.; Vuholm, S.; Roager, H.M.; Nielsen, D.S.; Krych, L.; Kristensen, M.; Astrup, A.; Hjorth, M.F. Prevotella Abundance Predicts Weight Loss Success in Healthy, Overweight Adults Consuming a Whole-Grain Diet Ad Libitum: A Post Hoc Analysis of a 6-Wk Randomized Controlled. Trial. J. Nutr. 2019, 149, 2174–2181. [Google Scholar] [CrossRef] [PubMed]
- Damms-Machado, A.; Mitra, S.; Schollenberger, A.E.; Kramer, K.M.; Meile, T.; Königsrainer, A.; Huson, D.H.; Bischoff, S.C. Effects of surgical and dietary weight loss therapy for obesity on gut microbiota composition and nutrient absorption. BioMed Res Int. 2015, 2015, 806248. [Google Scholar] [CrossRef]
- Korpela, K.; Flint, H.J.; Johnstone, A.M.; Lappi, J.; Poutanen, K.; Dewulf, E.; Delzenne, N.; De Vos, W.M.; Salonen, A. Gut Microbiota Signatures Predict Host and Microbiota Responses to Dietary Interventions in Obese Individuals. PLoS ONE 2014, 9, e90702. [Google Scholar] [CrossRef] [Green Version]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The controversial role of human gut lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Biesiekierski, J.R.; Jalanka, J.; Staudacher, H.M. Can Gut Microbiota Composition Predict Response to Dietary Treatments? Nutrients 2019, 11, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fragiadakis, G.K.; Wastyk, H.C.; Robinson, J.L.; Sonnenburg, E.D.; Sonnenburg, J.L.; Gardner, C.D. Long-term dietary intervention reveals resilience of the gut microbiota despite changes in diet and weight. Am. J. Clin. Nutr. 2020, 111, 1127–1136. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Tertile Min–Max | T1 (n = 121) 25.9–31.5 | T2 (n = 122) 31.5–35.0 | T3 (n = 121) 35.0–40.3 | P Trend & |
|---|---|---|---|---|
| Sex, female | 58 (47.9) | 57 (46.7) | 73 (60.3) | 0.064 |
| Age, years | 64.9 ± 5.2 | 64.3 ± 4.8 | 65.0 ± 5.1 | 0.591 |
| Intervention group | 55 (45.5) | 63 (51.6) | 66 (54.5) | 0.352 |
| Body weight, kg | 79.4 ± 9.1 | 88.4 ± 10.4 ** | 96.9 ± 12.0 **† | <0.001 |
| BMI, kg/m2 | 29.4 ± 1.4 | 33.1 ± 1.0 ** | 37.3 ± 1.5 **† | <0.001 |
| Waist circumference, cm | 102.2 ± 7.1 | 109.7 ± 7.4 ** | 117.5 ± 8.1 **† | <0.001 |
| Smoking Current smoker Former smoker Never smoked | 20 (16.5) 48 (39.7) 52 (43.0) | 21 (17.2) 47 (38.5) 54 (44.3) | 15 (12.4) 39 (32.2) 67 (55.4) | 0.369 |
| Education Primary school Secondary school Academic or graduate | 64 (52.9) 37 (30.6) 20 (16.5) | 68 (55.7) 39 (32.0) 15 (12.3) | 64 (52.9) 41 (33.9) 16 (13.2) | 0.880 |
| Recruiting center | 0.093 | |||
| Reus | 45 (37.2) | 39 (32.0) | 55 (45.5) | |
| Malaga | 76 (62.8) | 83 (68.0) | 66 (54.5) | |
| Hypercholesterolemia | 77 (63.6) | 82 (67.2) | 75 (62.0) | 0.685 |
| Hypertension | 110 (90.9) | 117 (95.9) | 116 (95.9) | 0.159 |
| T2DM prevalence | 17 (14.0) | 33 (27.0) * | 35 (28.9) * | 0.012 |
| Insulin treatment | 2 (1.7) | 9 (7.4) | 10 (8.3) | 0.057 |
| Metformin treatment | 10 (8.3) | 29 (23.8) * | 26 (21.5) * | 0.003 |
| Other anti diabetic drugs use | 12 (9.9) | 27 (22.1) * | 28 (23.1) * | 0.013 |
| Glucose, mg/dL | 103.9 ± 19.8 | 112.4 ± 28.7 * | 112.9 ± 25.8 * | 0.007 |
| HbA1c, % | 5.7 [0.6] | 5.9 [0.6] ** | 5.9 [0.8] * | 0.001 |
| Triglycerides, mg/dL | 152 [100] | 147 [90] | 155.5 [78] | 0.291 |
| Total cholesterol, mg/dL | 204.8 ± 38.6 | 197.0 ± 37.2 | 203.0 ± 37.1 | 0.241 |
| HDL-cholesterol, mg/dL | 50.3 ± 12.9 | 48.0 ± 12.5 | 48.6 ± 11.9 | 0.316 |
| LDL-cholesterol, mg/dL | 122.6 ± 34.2 | 114.8 ± 33.1 | 118.8 ± 33.0 | 0.193 |
| SBP, mm Hg | 139.0 ± 18.2 | 140.2 ± 14.8 | 141.3 ± 17.6 | 0.589 |
| DBP, mm Hg | 78.8 ± 9.6 | 80.6 ± 9.6 | 77.9 ± 10.5 | 0.099 |
| Tertile Min—Max | T1 (n = 115) −24.2–−4.5 | T2 (n = 115) −4.5–−0.7 | T3 (n = 115) −0.72–11.6 | P Trend & |
|---|---|---|---|---|
| Sex, female | 54 (47.0) | 62 (53.9) | 57 (49.6) | 0.567 |
| Age, years | 64.4 ± 5.1 | 64.8 ± 4.8 | 64.8 ± 5.3 | 0.788 |
| Recruiting center | 0.178 | |||
| Reus | 45 (39.1) | 48 (41.7) | 35 (30.4) | |
| Malaga | 70 (60.9) | 67 (58.3) | 80 (69.6) | |
| Intervention group | 95 (82.6) | 63 (54.8) ** | 15 (13.0) ** | <0.001 |
| Hypercholesterolemia | 69 (60.0) | 72 (62.6) | 78 (68.7) | 0.455 |
| Hypertension | 105 (91.3) | 110 (95.7) | 109 (94.8) | 0.345 |
| Type 2 diabetes prevalence | 25 (21.7) | 35 (30.4) | 21 (18.3) | 0.081 |
| Insulin treatment | 4 (3.5) | 10 (8.7) | 6 (5.2) | 0.226 |
| Metformin treatment | 19 (16.5) | 26 (22.6) | 17 (18.3) | 0.268 |
| Other anti diabetic drugs use | 19 (16.5) | 28 (24.3) | 17 (14.8) | 0.139 |
| Body weight, kg | 89.2 ± 13.0 | 89.6 ± 14.1 | 86.1 ± 10.9 | 0.066 |
| Change in body weight, kg | −7.2 ± 3.4 | −2.3 ± 1.0 ** | 1.5 ± 1.7 **†† | <0.001 |
| BMI, kg/m2 | 33.3 ± 3.6 | 33.8 ± 3.6 | 32.5 (3.1) † | 0.018 |
| Change in BMI, kg/m2 | −2.6 ± 1.3 | −0.8 ± 0.5 ** | 0.6 ± 0.7 **†† | <0.001 |
| Waist circumference, cm | 110.4 ± 10.1 | 111.6 ± 10.2 | 107.6 ± 9.8 *† | 0.007 |
| Change in waist circumference, cm | −7.4 ± 4.7 | −2.4 ± 3.8 ** | 1.2 ± 3.8 **†† | <0.001 |
| Glucose, mg/dL | 107.9 ± 22.7 | 114.9 ± 30.5 * | 106.3 ± 21.3 † | 0.023 |
| Change in glucose, mg/dL | −7.8 ± 15.8 | −1.9 ± 21.1 * | 3.6 ± 21.4 **† | <0.001 |
| HbA1c, % | 5.9 [0.6] | 5.9 [0.9] | 5.7 [0.6] | 0.086 |
| Changes in HbA1c, % | −0.2 [0.4] | 0.0 [0.3] * | 0.1 [0.3] **† | <0.001 |
| Triglycerides, mg/dL | 137.0 [78.0] | 153.0 [98.0] | 162.0 [92.0] | 0.571 |
| Change in triglycerides, mg/dL | −19.0 [60.0] | −8.5 [60.2] | −4.5 [76.2] | 0.595 |
| Total cholesterol, mg/dL | 201.5 ± 31.1 | 195.5 ± 40.2 | 205.1 ± 40.9 | 0.169 |
| Change in total cholesterol, mg/dL | −1.6 ± 27.3 | −0.8 ± 31.7 | −4.9 ± 39.2 | 0.614 |
| HDL-cholesterol, mg/dL | 48.2 ± 13.3 | 48.0 ± 12.0 | 50.1 ± 12.2 | 0.387 |
| Change in HDL-cholesterol, mg/dL | 3.0 ± 6.7 | 3.0 ± 7.2 | 1.0 ± 8.2 | 0.065 |
| LDL-cholesterol, mg/dL | 120.4 ± 28.7 | 113.9 ± 34.5 | 120.7 ± 36.9 | 0.232 |
| Change in LDL-cholesterol, mg/dL | −1.2 ± 24.2 | −0.8 ± 27.1 | −5.2 ± 35.3 | 0.462 |
| SBP, mm Hg | 140.1 ± 15.6 | 141.7 ± 17.3 | 139.2 ± 17.3 | 0.531 |
| Change in SBP, mm Hg | −6.8 ± 13.1 | −4.1 ± 16.0 | −2.0 ± 16.6 | 0.058 |
| DBP, mm Hg | 79.9 ± 9.6 | 79.0 ± 10.2 | 79.0 ± 10.1 | 0.713 |
| Change in DBP, mm Hg | −3.6 ± 8.2 | −1.0 ± 8.3 * | −1.1 ± 8.3 * | 0.027 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atzeni, A.; Galié, S.; Muralidharan, J.; Babio, N.; Tinahones, F.J.; Vioque, J.; Corella, D.; Castañer, O.; Vidal, J.; Moreno-Indias, I.; et al. Gut Microbiota Profile and Changes in Body Weight in Elderly Subjects with Overweight/Obesity and Metabolic Syndrome. Microorganisms 2021, 9, 346. https://doi.org/10.3390/microorganisms9020346
Atzeni A, Galié S, Muralidharan J, Babio N, Tinahones FJ, Vioque J, Corella D, Castañer O, Vidal J, Moreno-Indias I, et al. Gut Microbiota Profile and Changes in Body Weight in Elderly Subjects with Overweight/Obesity and Metabolic Syndrome. Microorganisms. 2021; 9(2):346. https://doi.org/10.3390/microorganisms9020346
Chicago/Turabian StyleAtzeni, Alessandro, Serena Galié, Jananee Muralidharan, Nancy Babio, Francisco José Tinahones, Jesús Vioque, Dolores Corella, Olga Castañer, Josep Vidal, Isabel Moreno-Indias, and et al. 2021. "Gut Microbiota Profile and Changes in Body Weight in Elderly Subjects with Overweight/Obesity and Metabolic Syndrome" Microorganisms 9, no. 2: 346. https://doi.org/10.3390/microorganisms9020346
APA StyleAtzeni, A., Galié, S., Muralidharan, J., Babio, N., Tinahones, F. J., Vioque, J., Corella, D., Castañer, O., Vidal, J., Moreno-Indias, I., Torres-Collado, L., Fernández-Carrión, R., Fitó, M., Olbeyra, R., Martínez-González, M. A., Bulló, M., & Salas-Salvadó, J. (2021). Gut Microbiota Profile and Changes in Body Weight in Elderly Subjects with Overweight/Obesity and Metabolic Syndrome. Microorganisms, 9(2), 346. https://doi.org/10.3390/microorganisms9020346