
Cranberry Proanthocyanidins and Dietary Oligosaccharides Synergistically Modulate Lactobacillus plantarum Physiology


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Propagation and Growth Phenotype Assay
2.2. Bacterial Metabolic End-Product Quantification
2.3. PAC Degradation Product Identification
2.4. RNA-seq Transcriptome Library Preparation
2.5. Bioinformatic and Statistical Analysis of Transcriptome Data
2.6. Differential Gene Expression Analysis
2.7. Transcriptome Functional Annotation and Enrichment Analysis
3. Results
3.1. Cranberry PACs Increase Biomass Accumulation during L. plantarum Dietary Oligosaccharide Fermentation
3.2. Cranberry Polyphenols Alter Oligosaccharide Utilization Phenotypes of Lactobacilli Strains
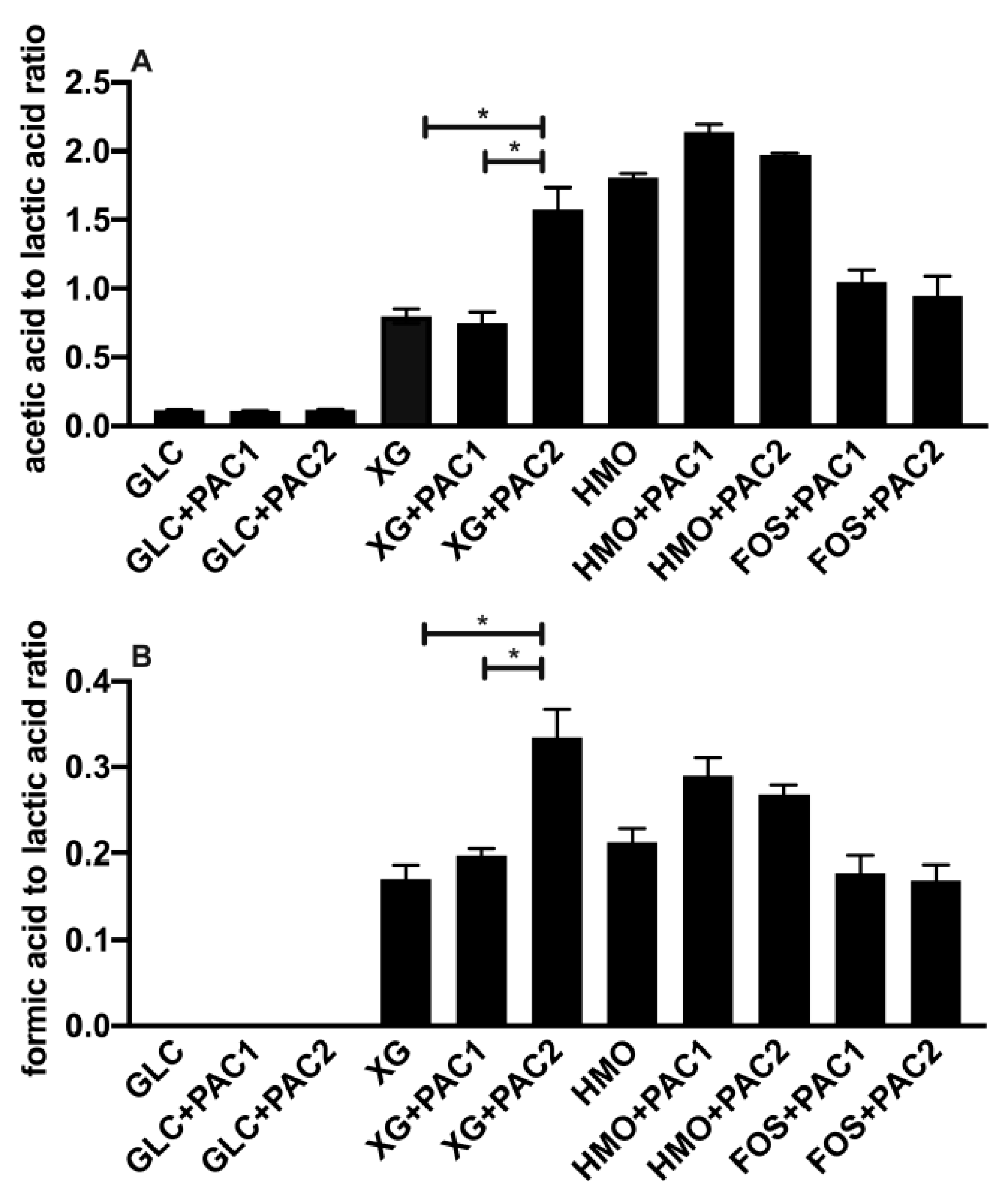
3.3. Cranberry PACs Alter Secreted Metabolic End-Products during Oligosaccharide Utilization
3.4. Biotransformation of Cranberry Phenolics Vary by Oligosaccharide Metabolism
3.5. PACs Generally Shift L. plantarum Transcriptomes in Concordance with Growth and Metabolic Phenotypes
3.6. Cranberry PACs Differentially Enrich Physiological Networks
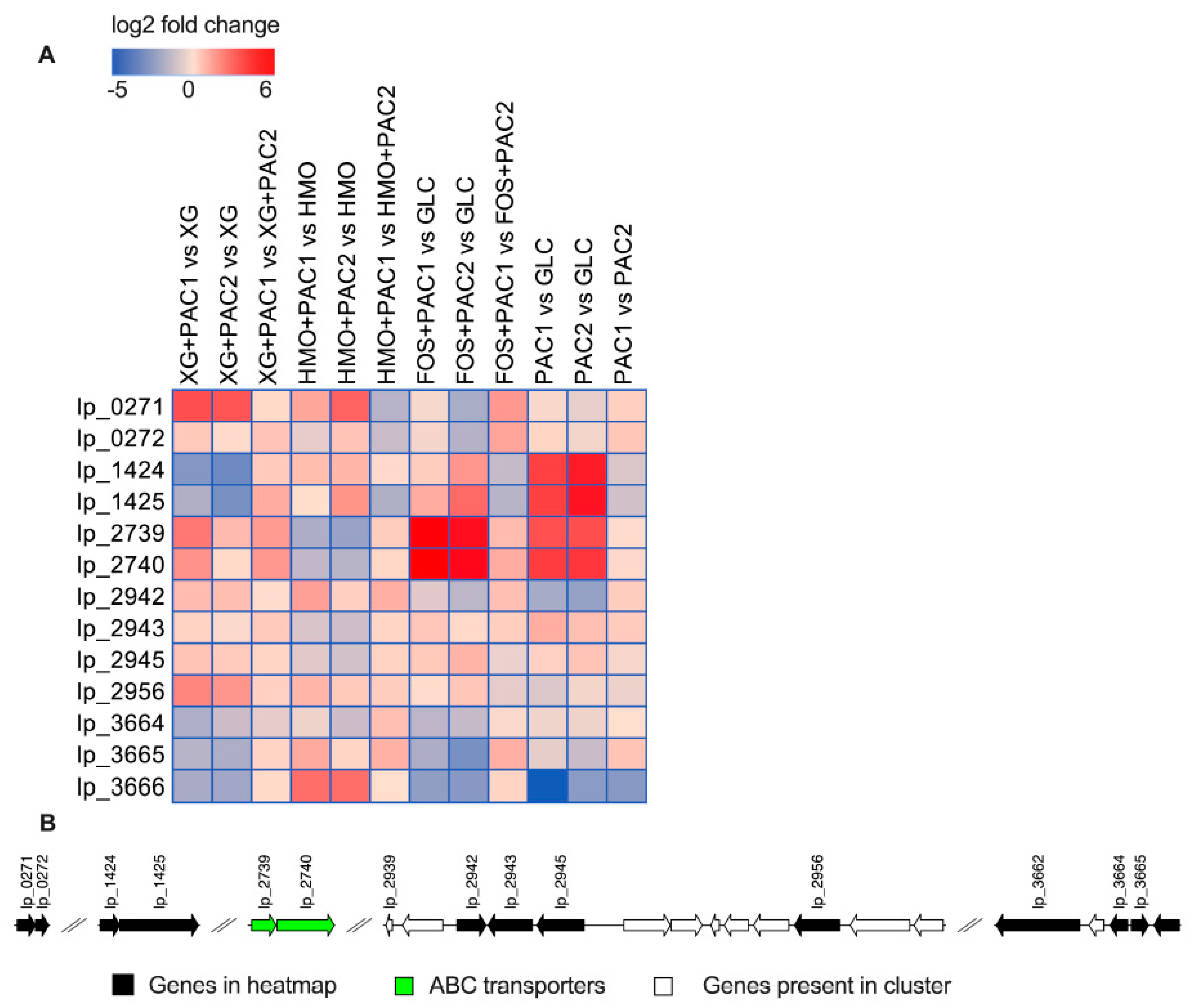
3.7. Polyphenol-Linked Genes Are Differentially Expressed during Synergistic Fermentations of Oligosaccharides with PACs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cerdá, B.; Tomás-Barberán, F.A.; Espín, J.C. Metabolism of antioxidant and chemopreventive ellagitannins from strawberries, raspberries, walnuts, and oak-aged wine in humans: Identification of biomarkers and individual variability. J. Agric. Food Chem. 2005, 53, 227–235. [Google Scholar] [CrossRef]
- Femia, A.P.; Caderni, G.; Vignali, F.; Salvadori, M.; Giannini, A.; Biggeri, A.; Gee, J.; Przybylska, K.; Cheynier, V.; Dolara, P. Effect of polyphenolic extracts from red wine and 4-OH-coumaric acid on 1,2-dimethylhydrazine-induced colon carcinogenesis in rats. Eur. J. Nutr. 2005, 44, 79–84. [Google Scholar] [CrossRef]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of Polyphenols on Gut Microbiota and Implications in Human Health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumberg, J.B.; Camesano, T.A.; Cassidy, A.; Kris-Etherton, P.; Howell, A.; Manach, C.; Ostertag, L.M.; Sies, H.; Skulas-Ray, A.; Vita, J.A. Cranberries and their bioactive constituents in human health. Adv. Nutr. 2013, 4, 618–632. [Google Scholar] [CrossRef] [Green Version]
- Ou, K.; Gu, L. Absorption and metabolism of proanthocyanidins. J. Funct. Foods 2014, 7, 43–53. [Google Scholar] [CrossRef]
- Novotny, J.A.; Baer, D.J.; Khoo, C.; Gebauer, S.K.; Charron, C.S. Cranberry Juice Consumption Lowers Markers of Cardiometabolic Risk, Including Blood Pressure and Circulating C-Reactive Protein, Triglyceride, and Glucose Concentrations in Adults. J. Nutr. 2015, 145, 1185–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foo, L.Y.; Lu, Y.; Howell, A.B.; Vorsa, N. A-Type Proanthocyanidin Trimers from Cranberry that Inhibit Adherence of Uropathogenic P-Fimbriated Escherichia coli. J. Nat. Prod. 2000, 63, 1225–1228. [Google Scholar] [CrossRef]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus plantarum with Functional Properties: An Approach to Increase Safety and Shelf-Life of Fermented Foods. Biomed Res. Int. 2018, 2018, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- De Vries, M.C.; Vaughan, E.E.; Kleerebezem, M.; de Vos, W.M. Lactobacillus plantarum—survival, functional and potential probiotic properties in the human intestinal tract. Int. Dairy J. 2006, 16, 1018–1028. [Google Scholar] [CrossRef]
- Rodríguez-Costa, S.; Cardelle-Cobas, A.; Roca-Saavedra, P.; Porto-Arias, J.J.; Miranda, J.M.; Cepeda, A. In vitro evaluation of the prebiotic effect of red and white grape polyphenolic extracts. J. Physiol. Biochem. 2018, 74, 101–110. [Google Scholar] [CrossRef]
- Nohynek, L.J.; Alakomi, H.-L.; Kähkönen, M.P.; Heinonen, M.; Helander, I.M.; Oksman-Caldentey, K.-M.; Puupponen-Pimiä, R.H. Berry phenolics: Antimicrobial properties and mechanisms of action against severe human pathogens. Nutr. Cancer 2006, 54, 18–32. [Google Scholar] [CrossRef]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Özcan, E.; Sun, J.; Rowley, D.C.; Sela, D.A. A Human Gut Commensal Ferments Cranberry Carbohydrates to Produce Formate. Appl. Environ. Microbiol. 2017, 83, e01097-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, C.M.; Ferreira, D. Oligosaccharides and Complex Carbohydrates: A New Paradigm for Cranberry Bioactivity. Molecules 2020, 25, 881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moro Cantu-Jungles, T.; do Nascimento, G.E.; Zhang, X.; Iacomini, M.; Cordeiro, L.M.C.; Hamaker, B.R. Soluble xyloglucan generates bigger bacterial community shifts than pectic polymers during in vitro fecal fermentation. Carbohydr. Polym. 2019, 206, 389–395. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, M.; Shi, T.; Guo, C.; Huang, Y.; Chen, Y.; Xie, M. Recovery of dietary fiber and polyphenol from grape juice pomace and evaluation of their functional properties and polyphenol compositions. Food Funct. 2017, 8, 341–351. [Google Scholar] [CrossRef] [PubMed]
- González-Aguilar, G.A.; Blancas-Benítez, F.J.; Sáyago-Ayerdi, S.G. Polyphenols associated with dietary fibers in plant foods: Molecular interactions and bioaccessibility. Curr. Opin. Food Sci. 2017, 13, 84–88. [Google Scholar] [CrossRef]
- Bazzocco, S.; Mattila, I.; Guyot, S.; Renard, C.M.G.C.; Aura, A.-M. Factors affecting the conversion of apple polyphenols to phenolic acids and fruit matrix to short-chain fatty acids by human faecal microbiota in vitro. Eur. J. Nutr. 2008, 47, 442–452. [Google Scholar] [CrossRef]
- Hotchkiss, A.T.; Nunez, A.; Strahan, G.D.; Chau, H.; White, A.; Marais, J.; Hom, K.; Vakkalanka, M.S.; Di, R.; Yam, K.L.; et al. Cranberry xyloglucan structure and inhibition of Escherichia coli adhesion to epithelial cells. J. Agric. Food Chem. 2015, 63, 5622–5633. [Google Scholar] [CrossRef]
- De Moura Bell, J.M.L.N.; Cohen, J.L.; de Aquino, L.F.M.C.; Lee, H.; de Melo Silva, V.L.; Liu, Y.; Domizio, P.; Barile, D. An integrated bioprocess to recover bovine milk oligosaccharides from colostrum whey permeate. J. Food Eng. 2018, 216, 27–35. [Google Scholar] [CrossRef]
- Krueger, C.G.; Chesmore, N.; Chen, X.; Parker, J.; Khoo, C.; Marais, J.P.J.; Shanmuganayagam, D.; Crump, P.; Reed, J.D. Critical reevaluation of the 4-(dimethylamino)cinnamaldehyde assay: Cranberry proanthocyanidin standard is superior to procyanidin A2 dimer for accurate quantification of proanthocyanidins in cranberry products. J. Funct. Foods 2016, 22, 13–19. [Google Scholar] [CrossRef]
- Sprouffske, K.; Wagner, A. Growthcurver: An R package for obtaining interpretable metrics from microbial growth curves. BMC Bioinform. 2016, 17, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Girón, A.; Queipo-Ortuño, M.I.; Boto-Ordóñez, M.; Muñoz-González, I.; Sánchez-Patán, F.; Monagas, M.; Martín-Álvarez, P.J.; Murri, M.; Tinahones, F.J.; Andrés-Lacueva, C.; et al. Comparative Study of Microbial-Derived Phenolic Metabolites in Human Feces after Intake of Gin, Red Wine, and Dealcoholized Red Wine. J. Agric. Food Chem. 2013, 61, 3909–3915. [Google Scholar] [CrossRef] [PubMed]
- Duary, R.K.; Batish, V.K.; Grover, S. Expression of the atpD gene in probiotic Lactobacillus plantarum strains under in vitro acidic conditions using RT-qPCR. Res. Microbiol. 2010, 161, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, B. BBMap Download | SourceForge.net. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 17 June 2020).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Project, G.; et al. The Sequence Alignment/Map format and SAMtools. Bioinforma. Appl. NOTE 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Blighe, K.; Rana, S.; Maintainer, M.L. Publication-Ready Volcano Plots with Enhanced Colouring and Labeling Version 1.4.0. 2019. Available online: https://git.bioconductor.org/packages/EnhancedVolcano (accessed on 21 January 2020).
- Young, M.D.; Davidson, N.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. goseq: Gene Ontology Testing for RNA-seq Datasets. 2017. Available online: http://129.217.206.11/packages/3.7/bioc/vignettes/goseq/inst/doc/goseq.pdf (accessed on 31 January 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Schwab, C.; Gänzle, M. Lactic acid bacteria fermentation of human milk oligosaccharide components, human milk oligosaccharides and galactooligosaccharides. FEMS Microbiol. Lett. 2011, 315, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kontula, P.; von Wright, A.; Mattila-Sandholm, T. Oat bran β-gluco- and xylo-oligosaccharides as fermentative substrates for lactic acid bacteria. Int. J. Food Microbiol. 1998, 45, 163–169. [Google Scholar] [CrossRef]
- Özcan, E.; Sela, D.A. Inefficient Metabolism of the Human Milk Oligosaccharides Lacto-N-tetraose and Lacto-N-neotetraose Shifts Bifidobacterium longum subsp. infantis Physiology. Front. Nutr. 2018, 5, 46. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and tannin-like compounds—Nature, occurrence, dietary intake and effects on nutrition and health. J. Sci. Food Agric. 2000, 1117, 1094–1117. [Google Scholar] [CrossRef]
- Engemann, A.; Hübner, F.; Rzeppa, S.; Humpf, H.-U. Intestinal Metabolism of Two A-type Procyanidins Using the Pig Cecum Model: Detailed Structure Elucidation of Unknown Catabolites with Fourier Transform Mass Spectrometry (FTMS). J. Agric. Food Chem. 2012, 60, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Teusink, B.; van Enckevort, F.H.J.; Francke, C.; Wiersma, A.; Wegkamp, A.; Smid, E.J.; Siezen, R.J. In Silico Reconstruction of the Metabolic Pathways of Lactobacillus plantarum: Comparing Predictions of Nutrient Requirements with Those from Growth Experiments. Appl. Environ. Microbiol. 2005, 71, 7253–7262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, N.; Curiel, J.A.; Reverón, I.; de Las Rivas, B.; Muñoz, R. Uncovering the Lactobacillus plantarum WCFS1 gallate decarboxylase involved in tannin degradation. Appl. Environ. Microbiol. 2013, 79, 4253–4263. [Google Scholar] [CrossRef] [Green Version]
- Reverón, I.; Jiménez, N.; Curiel, J.A.; Peñas, E.; López de Felipe, F.; de las Rivas, B.; Muñoz, R. Differential Gene Expression by Lactobacillus plantarum WCFS1 in Response to Phenolic Compounds Reveals New Genes Involved in Tannin Degradation. Appl. Environ. Microbiol. 2017, 83, e03387-16. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, L.; Reverón, I.; López de Felipe, F.; de las Rivas, B.; Muñoz, R. Unravelling the Reduction Pathway as an Alternative Metabolic Route to Hydroxycinnamate Decarboxylation in Lactobacillus plantarum. Appl. Environ. Microbiol. 2018, 84, e01123-18. [Google Scholar] [CrossRef] [Green Version]
- Déprez, S.; Brezillon, C.; Rabot, S.; Philippe, C.; Mila, I.; Lapierre, C.; Scalbert, A. Polymeric Proanthocyanidins Are Catabolized by Human Colonic Microflora into Low-Molecular-Weight Phenolic Acids. J. Nutr. 2000, 130, 2733–2738. [Google Scholar] [CrossRef] [Green Version]
- Marzorati, M.; Qin, B.; Hildebrand, F.; Klosterbuer, A.; Roughead, Z.; Roessle, C.; Rochat, F.; Raes, J.; Possemiers, S. Addition of acacia gum to a FOS/inulin blend improves its fermentation profile in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME®). J. Funct. Foods 2015, 16, 211–222. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Duysburgh, C.; Jiang, T.A.; Rebaza, M.; Pinheiro, I.; Marzorati, M. A combination of xylooligosaccharides and a polyphenol blend affect microbial composition and activity in the distal colon exerting immunomodulating properties on human cells. J. Funct. Foods 2018, 47, 163–171. [Google Scholar] [CrossRef]
- De Montijo-Prieto, S.; Castro, D.J.; Reina, J.C.; Jimenez-Valera, M.; Ruiz-Bravo, A. Draft genome sequence of Lactobacillus plantarum C4 (CECT 9567), a potential probiotic strain isolated from kefir. Arch. Microbiol. 2019, 201, 409–414. [Google Scholar] [CrossRef]
- Brooijmans, R.J.W.; de Vos, W.M.; Hugenholtz, J. Lactobacillus plantarum WCFS1 Electron Transport Chains. Appl. Environ. Microbiol. 2009, 75, 3580–3585. [Google Scholar] [CrossRef] [Green Version]
- McFeeters, R.F.; Chen, K.H. Utilization of electron acceptors for anaerobic mannitol metabolism by Lactobacillus plantarum. Compounds which serve as electron acceptors. Food Microbiol. 1986, 3, 73–81. [Google Scholar] [CrossRef]
- Lorenzen, J.; Steinwachs, S.; Unden, G. DMSO respiration by the anaerobic rumen bacterium Wolinella succinogenes. Arch. Microbiol. 1994, 162, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, J.; Huttunen, M.T.; Konings, W.N. Anaerobic transport in Escherichia coli membrane vesicles. J. Biol. Chem. 1975, 250, 6792–6798. [Google Scholar] [CrossRef]
- Serra, A.; Macià, A.; Romero, M.-P.; Anglés, N.; Morelló, J.-R.; Motilva, M.-J. Metabolic pathways of the colonic metabolism of procyanidins (monomers and dimers) and alkaloids. Food Chem. 2011, 126, 1127–1137. [Google Scholar] [CrossRef]
- Appeldoorn, M.M.; Vincken, J.-P.; Aura, A.-M.; Hollman, P.C.H.; Gruppen, H. Procyanidin Dimers Are Metabolized by Human Microbiota with 2-(3,4-Dihydroxyphenyl)acetic Acid and 5-(3,4-Dihydroxyphenyl)-γ-valerolactone as the Major Metabolites. J. Agric. Food Chem. 2009, 57, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.E.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef] [PubMed]
- López de Felipe, F.; Curiel, J.A.; Muñoz, R. Improvement of the fermentation performance of Lactobacillus plantarum by the flavanol catechin is uncoupled from its degradation. J. Appl. Microbiol. 2010, 109, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Francis, F.J.; Markakis, P.C. Food colorants: Anthocyanins. Crit. Rev. Food Sci. Nutr. 1989, 28, 273–314. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PACs in the Absence of Fermentable Carbohydrates | ||||||||
| PAC1 Pre | PAC1 Post | Delta | % Change | PAC2 Pre | PAC2 Post | Delta | % Change | |
| Procyanidin A2 | 8.60 | 2.59 ± 0.66 | −6.01 ± 0.66 | −69.86 ± 7.68 | 6.01 | 4.31 ± 0.59 | −1.70 ± 0.59 | −28.31 ± 9.86 |
| Procyanidin B2 | 0.02 | 0.88 ± 1.51 | 0.86 ± 1.51 | NA | 5.99 | 2.95 ± 2.86 | −3.04 ± 2.86 | −50.73 ± 47.85 |
| Epicatechin | 0.29 | 0.32 ± 0.04 | 0.03 ± 0.04 | 8.73 ± 13.79b | 1.19 | 0.81 ± 0.15 | −0.38 ± 0.15 | −31.65 ± 12.75b |
| 3-(3,4-dihydroxyphenyl)propionic acid | 0.00 | 0.00 ± 0.00 | NA | NA | 0.09 | 7.89 ± 2.97 | 7.80 ± 2.97 | 8558 ± 3254 |
| 3-(4-hydroxyphenyl)propionic acid | 0.00 | 0.24 ± 0.10 | 0.24 ±0.10 | NA | 0.20 | 48.38 ± 7.25 | 48.18 ± 7.25 | 24,054 ± 3618a |
| 3,4-dihydroxyphenylacetic acid | 0.02 | 0.02 ± 0.04 | 0.00 ± 0.00 | −1.82 ± 170.05 | 0.02 | 0.06 ± 0.02 | 0.04 ± 0.02 | 144.30 ± 88.06 |
| 4-hydroxyphenylacetic acid | 0.50 | 0.37 ± 0.18 | −0.13 ± 0.18 | −25.09 ± 35.96 | 0.46 | 0.47 ± 0.15 | 0.01 ± 0.15 | 0.71 ± 33.42 |
| Protocatechuic acid | 1.48 | 1.95 ± 0.59 | 0.47 ± 0.59 | 31.98 ± 35.91a * | 7.31 | 2.97 ± 0.38 | −4.35 ± 0.59 | −59.45 ± 5.21 * |
| Catechol | 0.00 | 0.65 ± 0.56 | 0.65 ± 0.56 | NA | 0.00 | 5.04 ± 0.92 | 5.04 ± 0.92 | NA |
| Syringic acid | 0.10 | 0.09 ± 0.02 | −0.01 ± 0.02 | −9.50 ± 20.14 * | 0.02 | 0.21 ± 0.08 | 0.19 ± 0.08 | 1049 ± 451b * |
| p-coumaric acid | 0.02 | 0.00 ±0.00 | −0.01 ± 0.00 | −79.77 ± 17.52b | 29.51 | 0.15 ± 0.02 | −29.36 ± 0.02 | −99.50 ± 0.06 |
| Xyloglucans | ||||||||
| PAC1 Pre | PAC1 Post | Delta | % Change | PAC2 Pre | PAC2 Post | Delta | % Change | |
| Procyanidin A2 | 4.38 | 0.36 ± 0.06 | −4.02 ±0.06 | −91.71 ± 1.45 | 6.83 | 3.25 ± 0.85 | −3.58 ± 0.85 | −52.40 ± 12.51 |
| Procyanidin B2 | 0.36 | 0.08 ± 0.07 | −0.28 ±0.07 | −78.24 ± 18.86 | 0.41 | 0.25 ± 0.05 | −0.16 ± 0.05 | −38.83 ± 11.24 |
| Epicatechin | 0.07 | 0.08 ± 0.00 | 0.01 ± 0.00 | 13.89 ± 4.81b | 0.23 | 0.18 ± 0.04 | −0.05 ± 0.04 | −21.37 ± 12.75b |
| 3-(3,4-dihydroxyphenyl)propionic acid | 0.00 | 0.18 ± 0.10 | 0.18 ± 0.10 | NA | 0.14 | 6.98 ± 0.80 | 6.84 ± 0.80 | 4815 ± 566 |
| 3-(4-hydroxyphenyl)propionic acid | 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | NA | 1.65 | 2.65 ± 0.38 | 1.00 ± 0.38 | 60.38 ± 23.19c |
| 3,4-dihydroxyphenylacetic acid | 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | NA | 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | NA |
| 4-hydroxyphenylacetic acid | 0.00 | 0.06 ± 0.05 | 0.06 ± 0.05 | NA | 0.00 | 0.06 ± 0.05 | 0.06 ± 0.05 | NA |
| Protocatechuic acid | 1.67 | 0.26 ± 0.03 | −1.41 ± 0.03 | −84.60 ± 1.93b | 8.72 | 0.18 ± 0.01 | −8.54 ± 0.01 | −97.74 ± 0.08 |
| Catechol | 0.27 | 2.42 ± 1.16 | 2.15 ± 1.16 | 800 ± 433 | 0.00 | 11.34 ± 1.01 | 11.34 ± 1.01 | NA |
| Syringic acid | 0.17 | 0.31 ± 0.10 | 0.14 ± 0.10 | 79.63 ± 58.88 | 0.33 | 0.29 ± 0.09 | −0.04 ± 0.09 | −11.43 ± 27.26c |
| p-coumaric acid | 0.04 | 1.60 ± 0.08 | 1.56 ± 0.08 | 3887 ± 197a * | 23.82 | 2.72 ± 0.30 | −21.12 ± 0.30 | −88.59 ± 1.26 * |
| Fructooligosaccharides | ||||||||
| PAC1 Pre | PAC1 Post | Delta | % Change | PAC2 Pre | PAC2 Post | Delta | % Change | |
| Procyanidin A2 | 0.00 | 0.04 ± 0.04 | 0.04 ± 0.04 | NA | 0.04 | 0.04 ± 0.04 | 0.00 ± 0.04 | −2.03 ± 100.69 |
| Procyanidin B2 | 0.03 | 0.09 ± 0.03 | 0.06 ± 0.03 | 207 ± 113 | 0.07 | 0.59 ± 0.07 | 0.52 ± 0.07 | 719.11 ± 94.31 |
| Epicatechin | 0.05 | 0.13 ± 0.02 | 0.08 ± 0.02 | 163.37 ± 30.82a | 0.17 | 0.42 ± 0.07 | 0.26 ± 0.07 | 155.60 ± 46.27a |
| 3-(3,4-dihydroxyphenyl)propionic acid | 0.04 | 0.00 ± 0.00 | −0.04 ± 0.00 | −100 ±0 | 0.00 | 6.51 ± 2.88 | 6.51 ± 2.88 | NA |
| 3-(4-hydroxyphenyl)propionic acid | 0.00 | 0.16 ± 0.06 | 0.16 ± 0.06 | NA | 0.20 | 54.55 ± 2.79 | 54.35 ± 2.79 | 26,959 ± 1386a |
| 3,4-dihydroxyphenylacetic acid | 0.03 | 0.02 ± 0.03 | −0.02 ± 0.03 | −56.42 ± 75.48 | 0.04 | 0.09 ± 0.07 | 0.05 ± 0.07 | 115.68 ± 155.38 |
| 4-hydroxyphenylacetic acid | 0.36 | 0.36 ±0.13 | −0.01 ± 0.13 | −1.50 ± 34.50 | 0.40 | 0.54 ± 0.10 | 0.13 ± 0.10 | 31.60 ± 25.11 |
| Protocatechuic acid | 1.39 | 2.15 ± 0.38 | 0.75 ± 0.38 | 54.16 ± 27.16a * | 6.26 | 2.64 ± 0.28 | −3.62 ± 0.28 | −57.79 ± 4.46 * |
| Catechol | 0.00 | 0.61 ± 0.56 | 0.61 ± 0.56 | NA | 0.00 | 2.95 ± 0.61 | 2.95 ± 0.61 | NA |
| Syringic acid | 0.08 | 0.07 ± 0.05 | −0.01 ± 0.05 | −8.14 ± 56.34 * | 0.01 | 0.18 ± 0.03 | 0.18 ± 0.03 | 3251 ± 553a * |
| p-coumaric acid | 0.02 | 0.00 ± 0.00 | −0.02 ± 0.00 | −98.76 ± 0.07b | 30.29 | 0.37 ± 0.02 | −29.92 ± 0.02 | −98.76 ± 94.31 |
| Human Milk Oligosaccharides | ||||||||
| PAC1 Pre | PAC1 Post | Delta | % Change | PAC2 Pre | PAC2 Post | Delta | % Change | |
| Procyanidin A2 | 5.90 | 4.05 ± 0.24 | −1.85 ± 0.24 | −31.34 ± 4.14 | 6.79 | 5.74 ± 0.14 | −1.04 ± 0.14 | −15.39 ± 2.13 |
| Procyanidin B2 | 0.01 | 0.90 ± 0.79 | 0.88 ± 0.79 | 6306 ± 5641 | 6.99 | 1.19 ± 0.99 | −5.80 ± 0.99 | −83.04 ± 14.10 |
| Epicatechin | 0.28 | 0.21 ± 0.05 | −0.07 ± 0.05 | −25.45 ± 17.03b | 1.13 | 0.50 ± 0.03 | −0.63 ± 0.03 | −55.69 ± 3.02b |
| 3-(3,4-dihydroxyphenyl)propionic acid | 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | NA | 0.00 | 6.03 ± 1.32 | 6.03 ± 1.32 | NA |
| 3-(4-hydroxyphenyl)propionic acid | 0.00 | 0.27 ± 0.16 | 0.27 ± 0.16 | NA | 0.37 | 48.25 ± 1.75 | 47.87 ± 1.75 | 12,846 ± 470b |
| 3,4-dihydroxyphenylacetic acid | 0.03 | 0.04 ± 0.01 | 0.01 ± 0.01 | 45.59 ± 34.87 | 0.08 | 0.14 ± 0.02 | 0.06 ±0.02 | 78.96 ± 27.39 |
| 4-hydroxyphenylacetic acid | 1.67 | 2.50 ± 0.38 | 0.83 ± 0.38 | 49.81 ± 23.05 | 1.25 | 1.91 ± 0.32 | 0.66 ± 0.32 | 52.46 ± 25.40 |
| Protocatechuic acid | 1.23 | 0.17 ± 0.02 | −1.07 ± 0.02 | −86.46 ± 1.50b | 7.91 | 0.27 ± 0.03 | −7.65 ± 0.03 | −96.59 ± 0.44 |
| Catechol | 0.00 | 2.46 ± 0.52 | 2.46 ± 0.52 | NA | 0.00 | 9.59 ± 2.77 | 9.59 ± 2.77 | NA |
| Syringic acid | 0.04 | 0.06 ± 0.02 | 0.02 ± 0.02 | 47.31 ± 48.26 | 0.03 | 0.25 ± 0.04 | 0.21 ± 0.04 | 639 ± 113bc |
| p-coumaric acid | 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | −73.49 ± 45.92b | 24.97 | 0.12 ± 0.02 | −24.85 ± 0.02 | −99.50 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Özcan, E.; Rozycki, M.R.; Sela, D.A. Cranberry Proanthocyanidins and Dietary Oligosaccharides Synergistically Modulate Lactobacillus plantarum Physiology. Microorganisms 2021, 9, 656. https://doi.org/10.3390/microorganisms9030656
Özcan E, Rozycki MR, Sela DA. Cranberry Proanthocyanidins and Dietary Oligosaccharides Synergistically Modulate Lactobacillus plantarum Physiology. Microorganisms. 2021; 9(3):656. https://doi.org/10.3390/microorganisms9030656
Chicago/Turabian StyleÖzcan, Ezgi, Michelle R. Rozycki, and David A. Sela. 2021. "Cranberry Proanthocyanidins and Dietary Oligosaccharides Synergistically Modulate Lactobacillus plantarum Physiology" Microorganisms 9, no. 3: 656. https://doi.org/10.3390/microorganisms9030656
APA StyleÖzcan, E., Rozycki, M. R., & Sela, D. A. (2021). Cranberry Proanthocyanidins and Dietary Oligosaccharides Synergistically Modulate Lactobacillus plantarum Physiology. Microorganisms, 9(3), 656. https://doi.org/10.3390/microorganisms9030656