
Novel High-Throughput Multiplex qPCRs for the Detection of Canine Vector-Borne Pathogens in the Asia-Pacific




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Multiplex qPCR Development
2.2. Sensitivity and Specificity of Quadruplex qPCRs
2.3. Samples & DNA Extraction
2.4. Methodological Comparison and Statistical Analysis
3. Results
3.1. Optimisation of Multiplex qPCRs for Canine VBPs
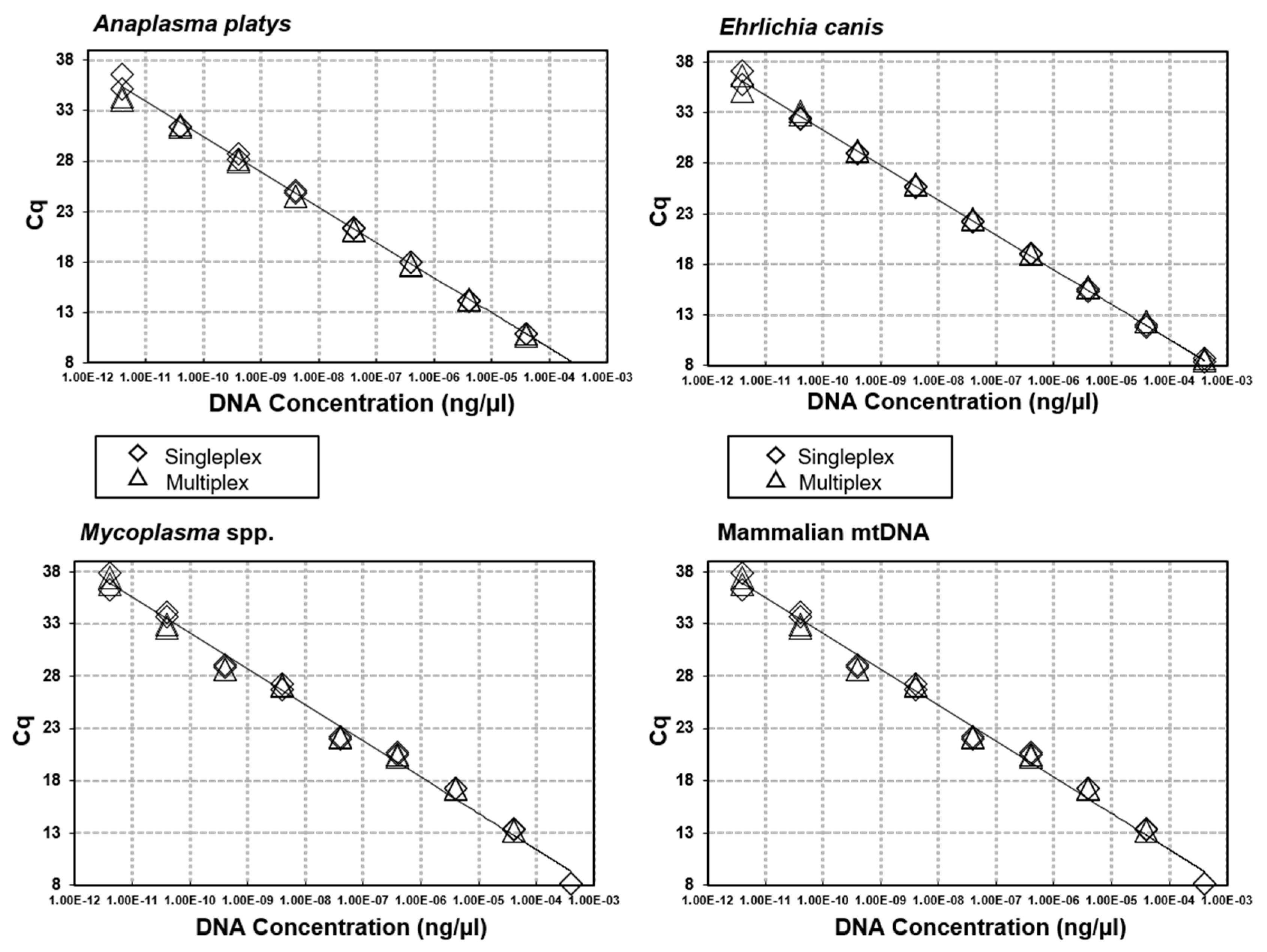
3.2. Sensitivity & Specificity of Multiplex qPCRs for Canine VBPs
3.3. Performance of Multiplex qPCRs on Field Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beugnet, F.; Marié, J.L. Emerging arthropod-borne diseases of companion animals in Europe. Vet. Parasitol. 2009, 163, 298–305. [Google Scholar] [CrossRef]
- Day, M.J. One health: The importance of companion animal vector-borne diseases. Parasites Vectors 2011, 4, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardy, J.L.; Loman, N.J. Towards a genomics-informed, real-time, global pathogen surveillance system. Nat. Rev. Genet. 2018, 19, 9–20. [Google Scholar] [CrossRef]
- Wang, S.F.; Wang, W.H.; Chang, K.; Chen, Y.H.; Tseng, S.P.; Yen, C.H.; Wu, D.C.; Chen, Y.M.A. Severe dengue fever outbreak in Taiwan. Am. J. Trop. Med. Hyg. 2016, 94, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Miró, G.; Müller, A.; Montoya, A.; Checa, R.; Marino, V.; Marino, E.; Fuster, F.; Escacena, C.; Descalzo, M.A.; Gálvez, R. Epidemiological role of dogs since the human leishmaniosis outbreak in Madrid. Parasites Vectors 2017, 10, 209. [Google Scholar] [CrossRef] [Green Version]
- Saldaña, A.; Calzada, J.E.; Pineda, V.; Perea, M.; Rigg, C.; González, K.; Santamaria, A.M.; Gottdenker, N.L.; Chaves, L.F. Risk factors associated with Trypanosoma cruzi exposure in domestic dogs from a rural community in Panama. Mem. Inst. Oswaldo Cruz 2015, 110, 936–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, L.R.; Beard, C.B.; Visser, S.N. Combatting the increasing threat of vector-borne disease in the United States with a national vector-borne disease prevention and control system. Am. J. Trop. Med. Hyg. 2019, 100, 242–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidd, L. Optimal vector-borne disease screening in dogs using both serology-based and polymerase chain reaction–based diagnostic panels. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 703–718. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Singh, H.; Sharma, P.; Singh, N.K.; Kashyap, N.; Singh, N.K. Development and application of multiplex PCR assay for the simultaneous detection of Babesia vogeli, Ehrlichia canis and Hepatozoon canis in dogs. Acta Trop. 2020, 212, 105713. [Google Scholar] [CrossRef]
- Little, S.E. Ehrlichiosis and anaplasmosis in dogs and cats. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 1121–1140. [Google Scholar] [CrossRef] [PubMed]
- Irwin, P.J.; Jefferies, R. Arthropod-transmitted diseases of companion animals in Southeast Asia. Trends Parasitol. 2004, 20, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Colella, V.; Nguyen, V.L.; Tan, D.Y.; Lu, N.; Fang, F.; Zhijuan, Y.; Wang, J.; Liu, X.; Chen, X.; Dong, J.; et al. Zoonotic vectorborne pathogens and ectoparasites of dogs and cats in Asia. Emerg. Infect. Dis. 2020, 26. [Google Scholar] [CrossRef]
- Huggins, L.G.; Koehler, A.V.; Ng-Nguyen, D.; Wilcox, S.; Schunack, B.; Inpankaew, T.; Traub, R.J. A novel metabarcoding diagnostic tool to explore protozoan haemoparasite diversity in mammals: A proof-of-concept study using canines from the tropics. Sci. Rep. 2019, 9, 12644. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.; Tsachev, I. Hepatozoon canis and hepatozoonosis in the dog. Trakia J. Sci. 2008, 6, 27–35. [Google Scholar]
- Irwin, P.J. Canine babesiosis: From molecular taxonomy to control. Parasites Vectors 2009, 2, S4. [Google Scholar] [CrossRef] [Green Version]
- Solano-Gallego, L.; Baneth, G. Babesiosis in dogs and cats—Expanding parasitological and clinical spectra. Vet. Parasitol. 2011, 181, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G.; Mathew, J.S.; Shkap, V.; Macintire, D.K.; Barta, J.R.; Ewing, S.A. Canine hepatozoonosis: Two disease syndromes caused by separate Hepatozoon spp. Trends Parasitol. 2003, 19, 27–31. [Google Scholar] [CrossRef]
- Muhlnickel, C.J.; Jefferies, R.; Morgan-Ryan, U.M.; Irwin, P.J. Babesia gibsoni infection in three dogs in Victoria. Aust. Vet. J. 2002, 80, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Rymaszewska, A.; Grenda, S. Bacteria of the genus Anaplasma—Characteristics of Anaplasma and their vectors: A review. Vet. Med. 2008, 53, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Huggins, L.G.; Koehler, A.V.; Ng-Nguyen, D.; Wilcox, S.; Schunack, B.; Inpankaew, T.; Traub, R.J. Assessment of a metabarcoding approach for the characterisation of vector-borne bacteria in canines from Bangkok, Thailand. Parasites Vectors 2019, 12, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrus, S.; Waner, T. Diagnosis of canine monocytotropic ehrlichiosis (Ehrlichia canis): An overview. Vet. J. 2011, 187, 292–296. [Google Scholar] [CrossRef]
- Australian Government “Ehrlichiosis in Dogs”. Available online: https://www.outbreak.gov.au/current-responses-to-outbreaks/ehrlichiosis-dogs (accessed on 2 February 2021).
- Hii, S.F.; Kopp, S.R.; Thompson, M.F.; O’Leary, C.A.; Rees, R.L.; Traub, R.J. Canine vector-borne disease pathogens in dogs from south-east Queensland and north-east Northern Territory. Aust. Vet. J. 2012, 90, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Barker, E.N.; Tasker, S.; Day, M.J.; Warman, S.M.; Woolley, K.; Birtles, R.; Georges, K.C.; Ezeokoli, C.D.; Newaj-Fyzul, A.; Campbell, M.D.; et al. Development and use of real-time PCR to detect and quantify Mycoplasma haemocanis and “Candidatus Mycoplasma haematoparvum” in dogs. Vet. Microbiol. 2010, 140, 167–170. [Google Scholar] [CrossRef]
- Hulme-Moir, K.L.; Barker, E.N.; Stonelake, A.; Helps, C.R.; Tasker, S. Use of real-time quantitative polymerase chain reaction to monitor antibiotic therapy in a dog with naturally acquired Mycoplasma haemocanis infection. J. Vet. Diagn. Investig. 2010, 22, 582–587. [Google Scholar] [CrossRef] [Green Version]
- Wengi, N.; Willi, B.; Boretti, F.S.; Cattori, V.; Riond, B.; Meli, M.L.; Reusch, C.E.; Lutz, H.; Hofmann-Lehmann, R. Real-time PCR-based prevalence study, infection follow-up and molecular characterization of canine hemotropic mycoplasmas. Vet. Microbiol. 2008, 126, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Snellgrove, A.N.; Krapiunaya, I.; Ford, S.L.; Stanley, H.M.; Wickson, A.G.; Hartzer, K.L.; Levin, M.L. Vector competence of Rhipicephalus sanguineus sensu stricto for Anaplasma platys. Ticks Tick Borne Dis. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Eddlestone, S.M.; Gaunt, S.D.; Neer, T.M.; Boudreaux, C.M.; Gill, A.; Haschke, E.; Corstvet, R.E. PCR detection of Anaplasma platys in blood and tissue of dogs during acute phase of experimental infection. Exp. Parasitol. 2007, 115, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Gaunt, S.D.; Beall, M.J.; Stillman, B.A.; Lorentzen, L.; Diniz, P.P.V.P.; Chandrashekar, R.; Breitschwerdt, E.B. Experimental infection and co-infection of dogs with Anaplasma platys and Ehrlichia canis: Hematologic, serologic and molecular findings. Parasites Vectors 2010, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Calzada, J.E.; Saldaña, A.; González, K.; Rigg, C.; Pineda, V.; Santamaría, A.M.; Rodríguez, I.; Gottdenker, N.L.; Laurenti, M.D.; Chaves, L.F. Cutaneous Leishmaniasis in dogs: Is high seroprevalence indicative of a reservoir role? Parasitology 2015, 142, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Maggi, R.G.; Birkenheuer, A.J.; Hegarty, B.C.; Bradley, J.M.; Levy, M.G.; Breitschwerdt, E.B. Comparison of serological and molecular panels for diagnosis of vector-borne diseases in dogs. Parasites Vectors 2014, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Buddhachat, K.; Meerod, T.; Pradit, W.; Siengdee, P.; Chomdej, S.; Nganvongpanit, K. Simultaneous differential detection of canine blood parasites: Multiplex high-resolution melting analysis (mHRM). Ticks Tick Borne Dis. 2020, 101370. [Google Scholar] [CrossRef]
- Megat Abd Rani, P.A.; Irwin, P.J.; Gatne, M.; Coleman, G.T.; Traub, R.J. Canine vector-borne diseases in India: A review of the literature and identification of existing knowledge gaps. Parasites Vectors 2010, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Massetti, L.; Colella, V.; Zendejas, P.A.; Ng-Nguyen, D.; Harriott, L.; Marwedel, L.; Wiethoelter, A.; Traub, R.J. High-throughput multiplex qPCRs for the surveillance of zoonotic species of canine hookworms. PLoS Negl. Trop. Dis. 2020, 14, e0008392. [Google Scholar] [CrossRef]
- Hii, S.F.; Senevirathna, D.; Llewellyn, S.; Inpankaew, T.; Odermatt, P.; Khieu, V.; Muth, S.; McCarthy, J.; Traub, R.J. Development and evaluation of a multiplex quantitative real-time polymerase chain reaction for hookworm species in human stool. Am. J. Trop. Med. Hyg. 2018, 99, 1186–1193. [Google Scholar] [CrossRef] [Green Version]
- Espy, M.J.; Uhl, J.R.; Sloan, L.M.; Buckwalter, S.P.; Jones, M.F.; Vetter, E.A.; Yao, J.D.C.; Wengenack, N.L.; Rosenblatt, J.E.; Cockerill, F.R.; et al. Real-time PCR in clinical microbiology: Applications for routine laboratory testing. Clin. Microbiol. Rev. 2006, 19, 165–256. [Google Scholar] [CrossRef] [Green Version]
- Maurin, M. Real-time PCR as a diagnostic tool for bacterial diseases. Expert Rev. Mol. Diagn. 2012, 12, 731–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleg, O.; Baneth, G.; Eyal, O.; Inbar, J.; Harrus, S. Multiplex real-time qPCR for the detection of Ehrlichia canis and Babesia canis vogeli. Vet. Parasitol. 2010, 173, 292–299. [Google Scholar] [CrossRef]
- Carslake, R.J.; Hill, K.E.; Sjölander, K.; Hii, S.F.; Prattley, D.; Acke, E. Cross-sectional survey of health management and prevalence of vector-borne diseases, endoparasites and ectoparasites in Samoan dogs. Aust. Vet. J. 2017, 95, 462–468. [Google Scholar] [CrossRef]
- Barker, E.N.; Langton, D.A.; Helps, C.R.; Brown, G.; Malik, R.; Shaw, S.E.; Tasker, S. Haemoparasites of free-roaming dogs associated with several remote Aboriginal communities in Australia. BMC Vet. Res. 2012, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, A.J.; Brown, G.; Norris, J.M.; Bosward, K.L.; Marriot, D.J.; Balakrishnan, N.; Breitschwerdt, E.B.; Malik, R. Vector-borne and zoonotic diseases of dogs in north-west New South Wales and the Northern Territory, Australia. BMC Vet. Res. 2017, 13, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialasiewicz, S.; Whiley, D.M.; Buhrer-Skinner, M.; Bautista, C.; Barker, K.; Aitken, S.; Gordon, R.; Muller, R.; Lambert, S.B.; Debattista, J.; et al. A novel gel-based method for self-collection and ambient temperature postal transport of urine for PCR detection of Chlamydia trachomatis. Sex. Transm. Infect. 2009, 85, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Saah, A.J.; Hoover, D.R. “Sensitivity” and “specificity” reconsidered: The meaning of these terms in analytical and diagnostic settings. Ann. Intern. Med. 1997, 126, 91–94. [Google Scholar] [CrossRef]
- Jefferies, R.; Ryan, U.M.; Irwin, P.J. PCR–RFLP for the detection and differentiation of the canine piroplasm species and its use with filter paper-based technologies. Vet. Parasitol. 2007, 144, 20–27. [Google Scholar] [CrossRef]
- Inokuma, H.; Okuda, M.; Ohno, K.; Shimoda, K.; Onishi, T. Analysis of the 18S rRNA gene sequence of a Hepatozoon detected in two Japanese dogs. Vet. Parasitol. 2002, 106, 265–271. [Google Scholar] [CrossRef]
- Inokuma, H.; Fujii, K.; Okuda, M.; Onishi, T.; Beaufils, J.-P.; Raoult, D.; Brouqui, P. Determination of the nucleotide sequences of heat shock operon groESL and the citrate synthase gene (gltA) of Anaplasma (Ehrlichia) platys for phylogenetic and diagnostic studies. Clin. Diagn. Lab. Immunol. 2002, 9, 1132–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gal, A.; Loeb, E.; Yisaschar-Mekuzas, Y.; Baneth, G. Detection of Ehrlichia canis by PCR in different tissues obtained during necropsy from dogs surveyed for naturally occurring canine monocytic ehrlichiosis. Vet. J. 2008, 175, 212–217. [Google Scholar] [CrossRef]
- Criado-Fornelio, A.; Martinez-Marcos, A.; Buling-Saraña, A.; Barba-Carretero, J.C. Presence of Mycoplasma haemofelis, Mycoplasma haemominutum and piroplasmids in cats from southern Europe: A molecular study. Vet. Microbiol. 2003, 93, 307–317. [Google Scholar] [CrossRef]
- Hatta, T.; Matsubayashi, M.; Miyoshi, T.; Islam, M.K.; Alim, M.A.; Anisuzzaman; Yamaji, K.; Fujisaki, K.; Tsuji, N. Quantitative PCR-based parasite burden estimation of Babesia gibsoni in the vector tick, Haemaphysalis longicornis (acari: Ixodidae), fed on an experimentally infected dog. J. Vet. Med. Sci. 2013, 75, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Qurollo, B.A.; Archer, N.R.; Schreeg, M.E.; Marr, H.S.; Birkenheuer, A.J.; Haney, K.N.; Thomas, B.S.; Breitschwerdt, E.B. Improved molecular detection of Babesia infections in animals using a novel quantitative real-time PCR diagnostic assay targeting mitochondrial DNA. Parasites Vectors 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Troskie, M.; de Villiers, L.; Leisewitz, A.; Oosthuizen, M.C.; Quan, M. Development and validation of a multiplex, real-time PCR assay for Babesia rossi and Babesia vogeli. Ticks Tick Borne Dis. 2019, 10, 421–432. [Google Scholar] [CrossRef]
- Costa-Júnior, L.M.; Zahler-Rinder, M.; Ribeiro, M.F.B.; Rembeck, K.; Rabelo, E.M.L.; Pfister, K.; Passos, L.M.F. Use of a Real Time PCR for detecting subspecies of Babesia canis. Vet. Parasitol. 2012, 188, 160–163. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Zhang, J.; Shi, Z.; Song, C.; Zheng, X.; Zhang, Y.; Hao, Y.; Dong, H.; Wei, L.; El-Mahallawy, H.S.; et al. Molecular detection of vector-borne agents in dogs from ten provinces of China. Parasites Vectors 2015, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirigireddy, K.R.; Ganta, R.R. Multiplex detection of Ehrlichia and Anaplasma species pathogens in peripheral blood by real-time reverse transcriptase-polymerase chain reaction. J. Mol. Diagn. 2005, 7, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Zhang, M.Z.; Stich, R.W.; Mitchell, W.J.; Zhang, S. Development of a tick-borne pathogen qPCR panel for detection of Anaplasma, Ehrlichia, Rickettsia, and Lyme disease Borrelia in animals. J. Microbiol. Methods 2018, 151, 83–89. [Google Scholar] [CrossRef]
- Koh, F.X.; Panchadcharam, C.; Tay, S.T. Vector-borne diseases in stray dogs in peninsular Malaysia and molecular detection of Anaplasma and Ehrlichia spp. from Rhipicephalus sanguineus (Acari: Ixodidae) ticks. J. Med. Entomol. 2016, 53, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Inpankaew, T.; Hii, S.F.; Chimnoi, W.; Traub, R.J. Canine vector-borne pathogens in semi-domesticated dogs residing in northern Cambodia. Parasites Vectors 2016, 9, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Ruttayaporn, N.; Saechan, V.; Jirapattharasate, C.; Vudriko, P.; Moumouni, P.F.A.; Cao, S.; Inpankaew, T.; Ybañez, A.P.; Suzuki, H.; et al. Molecular survey of canine vector-borne diseases in stray dogs in Thailand. Parasitol. Int. 2016, 65, 357–361. [Google Scholar] [CrossRef]
- Mason, R.J.; Lee, J.M.; Curran, J.M.; Moss, A.; Van Der Heide, B.; Daniels, P.W. Serological survey for Ehrlichia canis in urban dogs from the major population centres of northern Australia. Aust. Vet. J. 2001, 79, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Jefferies, R.; Ryan, U.M.; Jardine, J.; Broughton, D.K.; Robertson, I.D.; Irwin, P.J. Blood, bull Terriers and babesiosis: Further evidence for direct transmission of Babesia gibsoni in dogs. Aust. Vet. J. 2007, 85, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Greay, T.L.; Barbosa, A.D.; Rees, R.L.; Paparini, A.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. An Australian dog diagnosed with an exotic tick-borne infection: Should Australia still be considered free from Hepatozoon canis? Int. J. Parasitol. 2018, 48, 805–815. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Moreira, B.G.; Behlke, M.A.; Owczarzy, R. Design of LNA probes that improve mismatch discrimination. Nucleic Acids Res. 2006, 34, e60. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Fan, C.; Gao, H.; Yin, Y.; Wang, X.; Zhang, Y.; Wang, H. Leishmaniasis diagnosis via metagenomic next-Generation sequencing. Front. Cell. Infect. Microbiol. 2020, 10, 528884. [Google Scholar] [CrossRef]
- Bogema, D.R.; Deutscher, A.T.; Fell, S.; Collins, D.; Eamens, G.J.; Jenkinsa, C. Development and validation of a quantitative PCR assay using multiplexed hydrolysis probes for detection and quantification of Theileria orientalis isolates and differentiation of clinically relevant subtypes. J. Clin. Microbiol. 2015, 53, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.; Dorsey, G.; Hubbard, A.E.; Rosenthal, P.J.; Greenhouse, B. Gel versus capillary electrophoresis genotyping for categorizing treatment outcomes in two anti-malarial trials in Uganda. Malar. J. 2010, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorburn, F.; Bennett, S.; Modha, S.; Murdoch, D.; Gunson, R.; Murcia, P.R. The use of next generation sequencing in the diagnosis and typing of respiratory infections. J. Clin. Virol. 2015, 69, 96–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anis, E.; Hawkins, I.K.; Ilha, M.R.S.; Woldemeskel, M.W.; Saliki, J.T.; Wilkes, R.P. Evaluation of targeted next-generation sequencing for detection of bovine pathogens in clinical samples. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Hofstaedter, C.E.; Zhao, C.; Mattei, L.; Tanes, C.; Clarke, E.; Lauder, A.; Sherrill-Mix, S.; Chehoud, C.; Kelsen, J.; et al. Optimizing methods and dodging pitfalls in microbiome research. Microbiome 2017, 5, 52. [Google Scholar] [CrossRef]
- Huggins, L.G.; Koehler, A.V.; Schunack, B.; Inpankaew, T.; Traub, R.J. A host-specific blocking primer combined with optimal DNA extraction improves the detection capability of a metabarcoding protocol for canine vector-borne bacteria. Pathogens 2020, 9, 258. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.S.Y.; Teng, J.L.L.; Poon, R.W.S.; Choi, G.K.Y.; Chan, K.H.; Yeung, M.L.; Hui, J.J.Y.; Yuen, K.Y. Comparative evaluation of a point-of-care immunochromatographic test SNAP 4Dx with molecular detection tests for vector-borne canine pathogens in Hong Kong. Vector Borne Zoonotic Dis. 2011, 11, 1269–1277. [Google Scholar] [CrossRef] [Green Version]
- Portillo, A.; de Sousa, R.; Santibáñez, S.; Duarte, A.; Edouard, S.; Fonseca, I.P.; Marques, C.; Novakova, M.; Palomar, A.M.; Santos, M.; et al. Guidelines for the detection of Rickettsia spp. Vector Borne Zoonotic Dis. 2017, 17, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Svec, D.; Tichopad, A.; Novosadova, V.; Pfaffl, M.W.; Kubista, M. How good is a PCR efficiency estimate: Recommendations for precise and robust qPCR efficiency assessments. Biomol. Detect. Quantif. 2015, 3, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Millán, J.; Travaini, A.; Cevidanes, A.; Sacristán, I.; Rodríguez, A. Assessing the natural circulation of canine vector-borne pathogens in foxes, ticks and fleas in protected areas of Argentine Patagonia with negligible dog participation. Int. J. Parasitol. Parasites Wildl. 2019, 8, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Millán, J.; Velarde, R.; Delicado, V.; Negre, N.; Ribas, A.; Oleaga, Á.; Llaneza, L.; Esperón, F. High diversity of hemotropic mycoplasmas in Iberian wild carnivores. Comp. Immunol. Microbiol. Infect. Dis. 2018, 60, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Traub, R.J.; Irwin, P.; Dantas-Torres, F.; Tort, G.P.; Labarthe, N.V.; Inpankaew, T.; Gatne, M.; Linh, B.K.; Schwan, V.; Watanabe, M.; et al. Toward the formation of a Companion Animal Parasite Council for the Tropics (CAPCT). Parasites Vectors 2015, 8, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primers and Probes | Sequence (5′–3′) | Gene Target | Size (bp) | Final Conc. (nm) | Source |
|---|---|---|---|---|---|---|
| B. gibsoni, B. vogeli & H. canis | Api-F | AGGAAGWTT RAG GCAATAACAG | 18S rRNA | 180 | 700 | This study |
| Api-R | CTA GGCATT CCT CGT TMAWGATT | 18S rRNA | 180 | 700 | This study | |
| B. gibsoni | Bgib-Probe | /56-FAM/TATCCCTG G/ZEN/CCGAGAG GTC C/3IABkFQ/ | 18S rRNA | 60 | This study | |
| B. vogeli | Bvog-affinity-Probe | /5HEX/AG +T+T+G +TTC +C+TYGG/3IABkFQ/ | 18S rRNA | 400 | This study | |
| H. canis | Hcan-probe | /56-ROXN/TGAATGTG CATCGTGATGGGAAT AGA/3IAbRQSp/ | 18S rRNA | 100 | This study | |
| Equine Herpes Virus 4 | EHV-F | GATGACACTAGCG ACTTCGA | gB gene | 80 | 80 | [42] |
| EHV-R | CAGGGCAGAAACC ATAGACA | gB gene | 80 | 80 | [42] | |
| EHV-Probe | /5Cy5/TTTCGCGT G/TAO/C CTC CTCCAG/3IAbRQSp/ | gB gene | 200 | [42] | ||
| A. platys & E. canis | Ehr/Ana-F | TCA GAA CGA ACGCTGGC | 16S rRNA | 145–149 | 200 | This study |
| Ehr/Ana-R | CACCATTTCTAR TGGCTATYCC | 16S rRNA | 145–149 | 200 | This study | |
| A. platys | APlat-Probe | /56-FAM/CG GAT TT+T +TGTCGTAGCTTG CT+ATG/3IABkFQ/ | 16S rRNA | 50 | This study | |
| E. canis | ECan-ALT-Probe | /56-ROXN/TA GCC TCT GGCTAT A+G+G AAA TTGT/3IAbRQSp/ | 16S rRNA | 150 | This study | |
| Haemotropic Mycoplasma spp. | Myco-F-D1 | CAM GTC AAG TCATCATGCCC | 16S rRNA | 134 | 250 | This study |
| Myco-R-Mod1 | CGA ATT GCA GCC TYYTAT CC | 16S rRNA | 134 | 250 | This study | |
| Myco-ALT-Probe | /5HEX/TG +CAAA+C+G TGCTACAATGG/3IABkFQ/ | 16S rRNA | 134 | 200 | This study | |
| Mammalian mtDNA | Mam-F | CGACCTCGATGT TGGATCAG | mtDNA | 92 | 100 | [35] |
| Mam-R | GAACTCAGA TCA CGT AGG ACT TT | mtDNA | 92 | 100 | [35] | |
| Mam-Probe | /5Cy5/CCTAATGGTGC AGCAGC+TA+TTAA GG/3IAbRQSp/ | mtDNA | 92 | 50 | This study |
| VBP | cPCR | qPCR | Total Agreement (%) | Kappa (Agreement) | Kappa SE | |
|---|---|---|---|---|---|---|
| NEG | POS | |||||
| A. platys | NEG | 75 | 13 | 87 | 0.581 (moderate) | 0.098 |
| POS | 0 | 12 | ||||
| E. canis | NEG | 53 | 14 | 85 | 0.692 (substantial) | 0.071 |
| POS | 1 | 32 | ||||
| Mycoplasma spp. | NEG | 61 | 0 | 98 | 0.958 (high) | 0.03 |
| POS | 2 | 37 | ||||
| B. vogeli | NEG | 86 | 1 | 99 | 0.957 (high) | 0.043 |
| POS | 0 | 13 | ||||
| H. canis | NEG | 57 | 26 | 72 | 0.365 (fair) | 0.084 |
| POS | 2 | 15 | ||||
| VBP | NGS | qPCR | Total Agreement (%) | Kappa (Agreement) | Kappa SE | |
|---|---|---|---|---|---|---|
| NEG | POS | |||||
| A. platys | NEG | 73 | 2 | 96 | 0.893 (high) | 0.052 |
| POS | 2 | 23 | ||||
| E. canis | NEG | 51 | 8 | 89 | 0.777 (substantial) | 0.063 |
| POS | 3 | 38 | ||||
| Mycoplasma spp. | NEG | 60 | 2 | 95 | 0.893 (high) | 0.046 |
| POS | 3 | 35 | ||||
| B. vogeli | NEG | 85 | 2 | 97 | 0.872 (high) | 0.073 |
| POS | 1 | 12 | ||||
| H. canis | NEG | 55 | 7 | 89 | 0.770 (substantial) | 0.065 |
| POS | 4 | 34 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huggins, L.; Massetti, L.; Schunack, B.; Colella, V.; Traub, R. Novel High-Throughput Multiplex qPCRs for the Detection of Canine Vector-Borne Pathogens in the Asia-Pacific. Microorganisms 2021, 9, 1092. https://doi.org/10.3390/microorganisms9051092
Huggins L, Massetti L, Schunack B, Colella V, Traub R. Novel High-Throughput Multiplex qPCRs for the Detection of Canine Vector-Borne Pathogens in the Asia-Pacific. Microorganisms. 2021; 9(5):1092. https://doi.org/10.3390/microorganisms9051092
Chicago/Turabian StyleHuggins, Lucas, Luca Massetti, Bettina Schunack, Vito Colella, and Rebecca Traub. 2021. "Novel High-Throughput Multiplex qPCRs for the Detection of Canine Vector-Borne Pathogens in the Asia-Pacific" Microorganisms 9, no. 5: 1092. https://doi.org/10.3390/microorganisms9051092
APA StyleHuggins, L., Massetti, L., Schunack, B., Colella, V., & Traub, R. (2021). Novel High-Throughput Multiplex qPCRs for the Detection of Canine Vector-Borne Pathogens in the Asia-Pacific. Microorganisms, 9(5), 1092. https://doi.org/10.3390/microorganisms9051092