
Borrelia burgdorferi Sensu Lato in Questing and Engorged Ticks from Different Habitat Types in Southern Germany

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Collection of Samples
2.3. DNA Extraction
2.4. PCR Screening
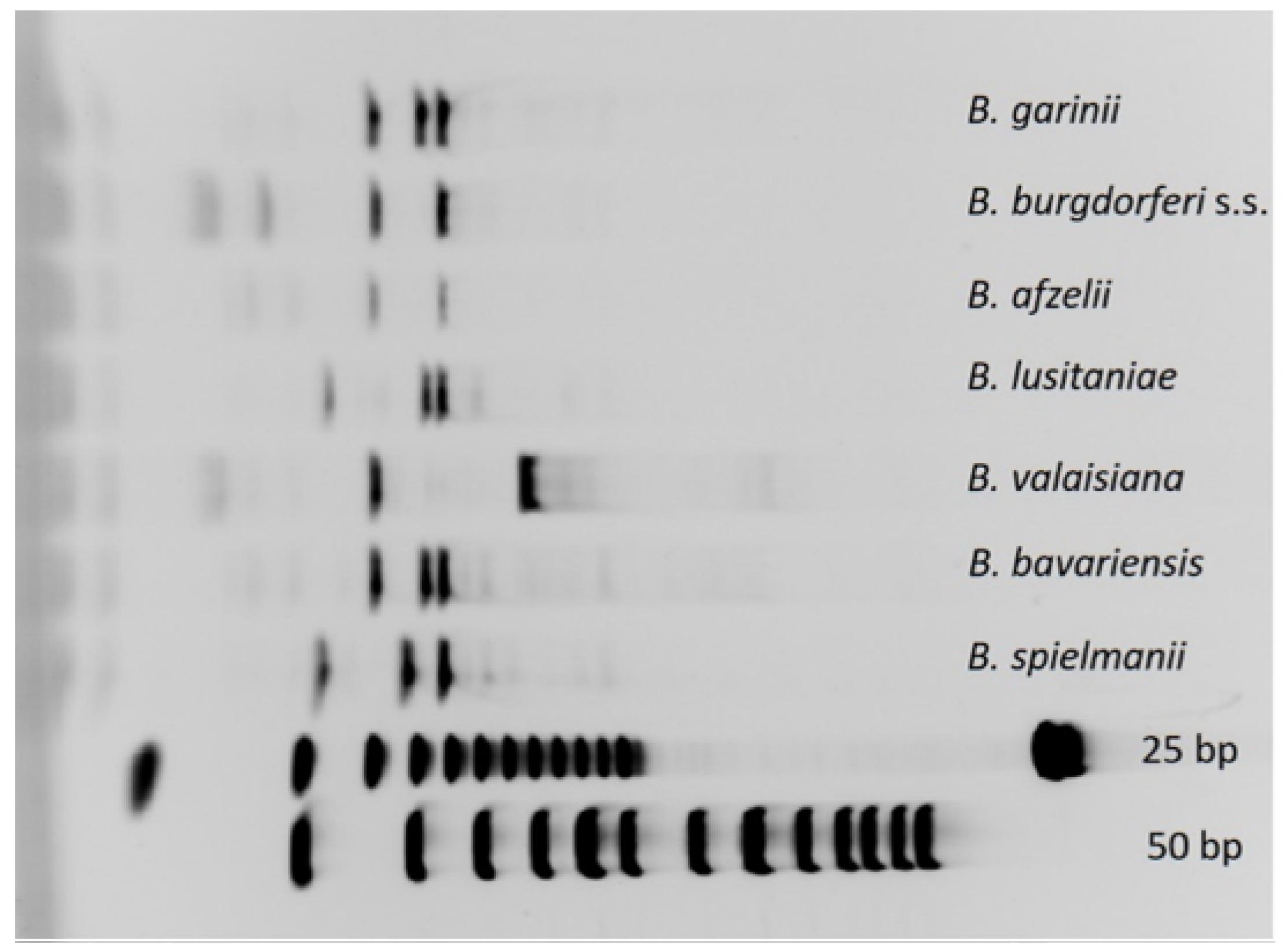
2.5. Species Identification by RFLP and Sequencing
2.6. Statistical Analysis
3. Results
3.1. Questing Ticks
3.2. Prevalence of Borrelia Burgdorferi Sensu Lato
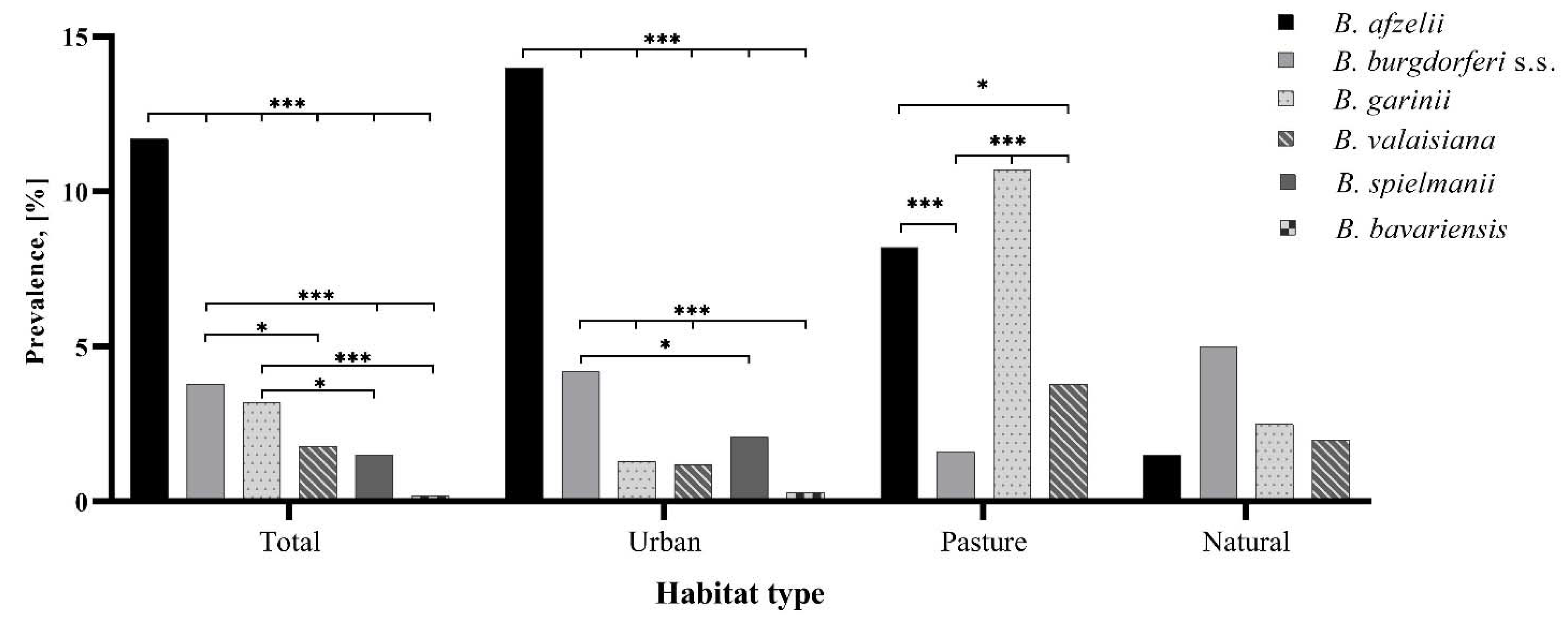
3.3. Species Identification of the B. burgdorferi s.l. Complex
3.3.1. Questing Ticks
3.3.2. Engorged Ticks (Cattle)
3.3.3. Engorged Ticks (Roe Deer)
3.3.4. Engorged Ticks (Wild Boar)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rizzoli, A.; Hauffe, H.; Carpi, G.; Vourc, H.G.; Neteler, M.; Rosa, R. Lyme borreliosis in Europe. Euro. Surveill. 2011, 16, 19906. [Google Scholar] [CrossRef]
- European Concerted Action on Lyme Borreliosis (EUCALB). Welcome to EUCALB.com! An Information Resource of the ESCMID Study Group, ESGBOR. Available online: http://www.eucalb.com/ (accessed on 4 June 2012).
- Grochowska, A.; Milewski, R.; Pancewicz, S.; Dunaj, J.; Czupryna, P.; Milewska, A.J.; Rog-Makal, M.; Grygorczuk, S.; Moniuszko-Malinowska, A. Comparison of tick-borne pathogen prevalence in Ixodes ricinus ticks collected in urban areas of Europe. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Schotthoefer, A.M.; Frost, H.M. Ecology and epidemiology of lyme borreliosis. Clin. Lab. Med. 2015, 35, 723. [Google Scholar] [CrossRef]
- Stanek, G.; Reiter, M. The expanding Lyme Borrelia complex—Clinical significance of genomic species? Clin. Microbiol. Infect. 2011, 17, 487–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strnad, M.; Honig, V.; Ruzek, D.; Grubhoffer, L.; Rego, R.O.M. Europe-wide meta-analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus ticks. Appl. Environ. Microb. 2017, 83, e00609-17. [Google Scholar] [CrossRef] [Green Version]
- Margos, G.; Fingerle, V.; Reynolds, S. Borrelia bavariensis: Vector switch, niche invasion, and geographical spread of a tick-borne bacterial parasite. Front Ecol. Evol. 2019, 7, 401. [Google Scholar] [CrossRef] [Green Version]
- Mysterud, A.; Stigum, V.M.; Jaarsma, R.I.; Sprong, H. Genospecies of Borrelia burgdorferi sensu lato detected in 16 mammal species and questing ticks from northern Europe. Sci. Rep. 2019, 9, 5088. [Google Scholar] [CrossRef] [PubMed]
- Halos, L.; Bord, S.; Cotte, V.; Gasqui, P.; Abrial, D.; Barnouin, J.; Boulouis, H.J.; Vayssier-Taussat, M.; Vourc’h, G. Ecological factors characterizing the prevalence of bacterial tick-borne pathogens in Ixodes ricinus ticks in pastures and woodlands. Appl. Environ. Microbiol. 2010, 76, 4413–4420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okeyo, M.; Hepner, S.; Rollins, R.E.; Hartberger, C.; Straubinger, R.K.; Marosevic, D.; Bannister, S.A.; Bormane, A.; Donaghy, M.; Sing, A.; et al. Longitudinal study of prevalence and spatio-temporal distribution of Borrelia burgdorferi sensu lato in ticks from three defined habitats in Latvia, 1999–2010. Environ. Microbiol. 2020, 22, 5033–5047. [Google Scholar] [CrossRef]
- Millins, C.; Gilbert, L.; Medlock, J.; Hansford, K.; Thompson, D.B.A.; Biek, R. Effects of conservation management of landscapes and vertebrate communities on Lyme borreliosis risk in the United Kingdom. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, B.F.; Keesing, F.; Ostfeld, R.S. Effect of forest fragmentation on Lyme disease risk. Conserv. Biol. 2003, 17, 267–272. [Google Scholar] [CrossRef] [Green Version]
- LoGiudice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: Effects of host diversity and community composition on Lyme disease risk. Proc. Natl. Acad. Sci. USA 2003, 100, 567–571. [Google Scholar] [CrossRef] [Green Version]
- Hansford, K.M.; Fonville, M.; Gillingham, E.L.; Coipan, E.C.; Pietzsch, M.E.; Krawczyk, A.I.; Vaux, A.G.C.; Cull, B.; Sprong, H.; Medlock, J.M. Ticks and Borrelia in urban and peri-urban green space habitats in a city in southern England. Ticks Tick-Borne Dis. 2017, 8, 353–361. [Google Scholar] [CrossRef]
- Kurtenbach, K.; Hanincova, K.; Tsao, J.I.; Margos, G.; Fish, D.; Ogden, N.H. Fundamental processes in the evolutionary ecology of Lyme borreliosis. Nat. Rev. Microbiol. 2006, 4, 660–669. [Google Scholar] [CrossRef]
- Margos, G.; Vollmer, S.A.; Cornet, M.; Garnier, M.; Fingerle, V.; Wilske, B.; Bormane, A.; Vitorino, L.; Collares-Pereira, M.; Drancourt, M.; et al. A new Borrelia species defined by multilocus sequence analysis of housekeeping genes. Appl. Environ. Microbiol. 2009, 75, 5410–5416. [Google Scholar] [CrossRef] [Green Version]
- Richter, D.; Matuschka, F.R. Modulatory effect of cattle on risk for Lyme disease. Emerg. Infect. Dis. 2006, 12, 1919–1923. [Google Scholar] [CrossRef] [PubMed]
- Tsao, J.I. Reviewing molecular adaptations of Lyme borreliosis spirochetes in the context of reproductive fitness in natural transmission cycles. Vet. Res. 2009, 40, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venczel, R.; Knoke, L.; Pavlovic, M.; Dzaferovic, E.; Vaculova, T.; Silaghi, C.; Overzier, E.; Konrad, R.; Kolencik, S.; Derdakova, M.; et al. A novel duplex real-time PCR permits simultaneous detection and differentiation of Borrelia miyamotoi and Borrelia burgdorferi sensu lato. Infection 2016, 44, 47–55. [Google Scholar] [CrossRef]
- Tappe, J.; Jordan, D.; Janecek, E.; Fingerle, V.; Strube, C. Revisited: Borrelia burgdorferi sensu lato infections in hard ticks (Ixodes ricinus) in the city of Hanover (Germany). Parasite Vector 2014, 7, 441. [Google Scholar] [CrossRef] [Green Version]
- May, K.; Jordan, D.; Fingerle, V.; Strube, C. Borrelia burgdorferi sensu lato and co-infections with Anaplasma phagocytophilum and Rickettsia spp. in Ixodes ricinus in Hamburg, Germany. Med. Vet. Entomol. 2015, 29, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Franke, J.; Hildebrandt, A.; Meier, F.; Straube, E.; Dorn, W. Prevalence of Lyme disease agents and several emerging pathogens in questing ticks from the german baltic coast. J. Med. Entomol. 2011, 48, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Fingerle, V.; Munderloh, U.G.; Liegl, G.; Wilske, B. Coexistence of ehrlichiae of the phagocytophila group with Borrelia burgdorferi in Ixodes ricinus from Southern Germany. Med. Microbiol. Immun. 1999, 188, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Blazejak, K.; Raulf, M.K.; Janecek, E.; Jordan, D.; Fingerle, V.; Strube, C. Shifts in Borrelia burgdorferi (s.l.) geno-species infections in Ixodes ricinus over a 10-year surveillance period in the city of Hanover (Germany) and Borrelia miyamotoi-specific Reverse Line Blot detection. Parasite Vector 2018, 11, 304. [Google Scholar] [CrossRef] [Green Version]
- Eshoo, M.W.; Crowder, C.D.; Carolan, H.E.; Rounds, M.A.; Ecker, D.J.; Haag, H.; Mothes, B.; Nolte, O. Broad-Range survey of tick-borne pathogens in southern germany reveals a high prevalence of Babesia microti and a diversity of other tick-borne pathogens. Vector-Borne Zoonot 2014, 14, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Bingsohn, L.; Beckert, A.; Zehner, R.; Kuch, U.; Oehme, R.; Kraiczy, P.; Amendt, J. Prevalences of tick-borne encephalitis virus and Borrelia burgdorferi sensu lato in Ixodes ricinus populations of the Rhine-Main region, Germany. Ticks Tick-Borne Dis. 2013, 4, 207–213. [Google Scholar] [CrossRef]
- Schicht, S.; Junge, S.; Schnieder, T.; Strube, C. Prevalence of Anaplasma phagocytophilum and Coinfection with Borrelia burgdorferi Sensu Lato in the Hard Tick Ixodes ricinus in the City of Hanover (Germany). Vector-Borne Zoonot 2011, 11, 1595–1597. [Google Scholar] [CrossRef]
- Raileanu, C.; Tauchmann, O.; Vasic, A.; Wohnke, E.; Silaghi, C. Borrelia miyamotoi and Borrelia burgdorferi (sensu lato) identification and survey of tick-borne encephalitis virus in ticks from north-eastern Germany. Parasite Vector 2020, 13, 106. [Google Scholar] [CrossRef]
- Zubrikova, D.; Wittmann, M.; Honig, V.; Svec, P.; Vichova, B.; Essbauer, S.; Dobler, G.; Grubhoffer, L.; Pfister, K. Prevalence of tick-borne encephalitis virus and Borrelia burgdorferi sensu lato in Ixodes ricinus ticks in Lower Bavaria and Upper Palatinate, Germany. Ticks Tick-Borne Dis. 2020, 11, 101375. [Google Scholar] [CrossRef]
- Overzier, E.; Pfister, K.; Thiel, C.; Herb, I.; Mahling, M.; Silaghi, C. Diversity of Babesia and Rickettsia Species in Questing Ixodes ricinus: A longitudinal study in urban, pasture, and natural habitats. Vector-Borne Zoonot 2013, 13, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, C.; Pfister, K.; Overzier, E. Molecular investigation for bacterial and protozoan tick-borne pathogens in wild boars (Sus scrofa) from Southern Germany. Vector-Borne Zoonot 2014, 14, 371–373. [Google Scholar] [CrossRef] [Green Version]
- Derdakova, M.; Beati, L.; Pet’ko, B.; Stanko, M.; Fish, D. Genetic variability within Borrelia burgdorferi sensu lato genospecies established by PCR-single-strand conformation polymorphism analysis of the rrfA-rrlB intergenic spacer in Ixodes ricinus ticks from the Czech Republic. Appl. Environ. Microbiol. 2003, 69, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Mehlhorn, H.; Mehlhorn, T.; Muller, M.; Vogt, M.; Rissland, J. Tick survey for prevalent pathogens in peri-urban recreation sites in Saarland and Rhineland-Palatinate (Germany). Parasitol. Res. 2016, 115, 1167–1172. [Google Scholar] [CrossRef]
- Szekeres, S.; Lugner, J.; Fingerle, V.; Margos, G.; Foldvari, G. Prevalence of Borrelia miyamotoi and Borrelia burgdorferi sensu lato in questing ticks from a recreational coniferous forest of East Saxony, Germany. Ticks Tick-Borne Dis. 2017, 8, 922–927. [Google Scholar] [CrossRef]
- Schwarz, A.; Honig, V.; Vavruskova, Z.; Grubhoffer, L.; Balczun, C.; Albring, A.; Schaub, G.A. Abundance of Ixodes ricinus and prevalence of Borrelia burgdorferi s.l. in the nature reserve Siebengebirge, Germany, in comparison to three former studies from 1978 onwards. Parasite Vector 2012, 5, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galfsky, D.; Krol, N.; Pfeffer, M.; Obiegala, A. Long-term trends of tick-borne pathogens in regard to small mammal and tick populations from Saxony, Germany. Parasite Vector 2019, 12, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fingerle, V.; Schulte-Spechtel, U.C.; Ruzic-Sabljic, E.; Leonhard, S.; Hofmann, H.; Weber, K.; Pfister, K.; Strle, F.; Wilske, B. Epidemiological aspects and molecular characterization of Borrelia burgdorferi s.l. from southern Germany with special respect to the new species Borrelia spielmanii sp. nov. Int. J. Med. Microbiol. 2008, 298, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Rauter, C.; Hartung, T. Prevalence of Borrelia burgdorferi sensu lato genospecies in Ixodes ricinus ticks in Europe: A metaanalysis. Appl. Environ. Microb. 2005, 71, 7203–7216. [Google Scholar] [CrossRef] [Green Version]
- Matuschka, F.R.; Fischer, P.; Heiler, M.; Blumcke, S.; Spielman, A. Stage-associated risk of transmission of the lyme-disease spirochete by European ixodes ticks. Parasitol Res. 1992, 78, 695–698. [Google Scholar] [CrossRef]
- Ehrmann, S.; Ruyts, S.C.; Scherer-Lorenzen, M.; Bauhus, J.; Brunet, J.; Cousins, S.A.O.; Deconchat, M.; Decocq, G.; De Frenne, P.; De Smedt, P.; et al. Habitat properties are key drivers of Borrelia burgdorferi (s.l.) prevalence in Ixodes ricinus populations of deciduous forest fragments. Parasit Vectors 2018, 11, 23. [Google Scholar] [CrossRef]
- Reis, C.; Cote, M.; Paul, R.E.; Bonnet, S. Questing ticks in suburban forest are infected by at least six tick-borne pathogens. Vector Borne Zoonotic Dis. 2011, 11, 907–916. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Talleklint, L. Incompetence of roe deer as reservoirs of the Lyme borreliosis spirochete. J. Med. Entomol. 1992, 29, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Kurtenbach, K.; De Michelis, S.; Etti, S.; Schafer, S.M.; Sewell, H.S.; Brade, V.; Kraiczy, P. Host association of Borrelia burgdorferi sensu lato—The key role of host complement. Trends Microbiol. 2002, 10, 74–79. [Google Scholar] [CrossRef]
- Fingerle, V.; Wilske, B. Abschlußbericht zur im Jahr 2004 durchgeführten Studie “Epidemiologische Aspekte zeckenübertragener Erkrankungen in Bayern: Lyme-Borreliose”. Abschlußbericht Gesundh. Bayern Akt. 2005, 17. [Google Scholar]
- Rosef, O.; Paulauskas, A.; Radzijevskaja, J. Prevalence of Borrelia burgdorferi sensu lato and Anaplasma phagocytophilum in questing Ixodes ricinus ticks in relation to the density of wild cervids. Acta Vet. Scand. 2009, 51, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grygorczuk, S.; Peter, O.; Kondrusik, M.; Moniuszko, A.; Zajkowska, J.; Dunaj, J.; Zukiewicz-Sobczak, W.; Pancewicz, S. Assessment of the frequency of different Borrelia burgdorferi sensu lato species in patients with Lyme borreliosis from north-east Poland by studying preferential serologic response and DNA isolates. Ann. Agric. Environ. Med. 2013, 20, 21–29. [Google Scholar]
- Norte, A.C.; Margos, G.; Becker, N.S.; Albino Ramos, J.; Nuncio, M.S.; Fingerle, V.; Araujo, P.M.; Adamik, P.; Alivizatos, H.; Barba, E.; et al. Host dispersal shapes the population structure of a tick-borne bacterial pathogen. Mol. Ecol. 2020, 29, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Heylen, D.; Sprong, H.; van Oers, K.; Fonville, M.; Leirs, H.; Matthysen, E. Are the specialized bird ticks, Ixodes arboricola and I. frontalis, competent vectors for Borrelia burgdorferi sensu lato? Environ. Microbiol. 2014, 16, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Vollmer, S.A.; Ogden, N.H.; Fish, D. Population genetics, taxonomy, phylogeny and evolution of Borrelia burgdorferi sensu lato. Infect Genet. Evol. 2011, 11, 1545–1563. [Google Scholar] [CrossRef] [Green Version]
- Kilpatrick, H.J.; Labonte, A.M.; Stafford, K.C. The relationship between deer density, tick abundance, and human cases of lyme disease in a residential community. J. Med. Entomol. 2014, 51, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Telford, S.R., 3rd; Mather, T.N.; Moore, S.I.; Wilson, M.L.; Spielman, A. Incompetence of deer as reservoirs of the Lyme disease spirochete. Am. J. Trop. Med. Hyg. 1988, 39, 105–109. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Feeding Status | Habitat | Site | Female | Male | Nymph | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pos/No. Total | % | 95% CI | Pos/No. Total | % | 95% CI | Pos/No. Total | % | 95% CI | Pos/No. Total | % | 95% CI | |||
| Questing ticks | Urban | M2 | 46/120 | 38.3 | 29.6–47.7 | 47/120 | 39.2 | 30.4–48.5 | 19/120 | 15.8 | 9.8–23.6 | 112/360 | 31.1 | 26.5–36.1 |
| M3 | 28/135 | 20.7 | 14.3–28.6 | 30/124 | 24.2 | 17.0–32.7 | 22/140 | 15.7 | 10.1–22.8 | 80/399 | 20.1 | 16.4–24.3 | ||
| R1 | 45/120 | 37.5 | 28.8–46.8 | 32/120 | 26.7 | 19.0–35.5 | 29/120 | 24.2 | 16.8–32.8 | 106/360 | 29.4 | 24.9–34.4 | ||
| B | 29/79 | 36.7 | 26.1–48.3 | 13/71 | 18.3 | 10.1–29.3 | 0/120 | 0.0 | 0.0–3.0 | 42/270 | 15.6 | 11.7–20.4 | ||
| Total | 148/454 | 32.6 | 28.3–37.1 | 122/435 | 28.0 | 23.9–32.5 | 70/500 | 14.0 | 11.1–17.4 | 340/1389 | 24.5 | 22.3–26.8 | ||
| Pasture | K | 33/93 | 35.5 | 25.8–46.1 | 35/132 | 26.5 | 19.2–34.9 | 34/140 | 24.3 | 17.4–32.3 | 102/365 | 27.9 | 23.6–32.8 | |
| Natural | T | 9/33 | 27.3 | 13.3–45.5 | 8/46 | 17.4 | 7.8–31.4 | 10/120 | 8.3 | 4.1–14.8 | 27/199 | 13.6 | 9.5–19.0 | |
| All sites | Total | 190/580 | 32.8 | 29.0–36.7 | 165/613 | 26.9 | 23.4–30.6 | 114/760 | 15.0 | 12.5–17.7 | 469/1953 | 24.0 | 22.2–26.0 | |
| Cattle | Pasture | K | 4/60 | 6.7 | 1/4 | 25 | na (1) | na (1) | na (1) | na (1) | na (1) | na (1) | ||
| Roe deer | Natural | T | 3/206 | 1.5 | 13/125 | 10.4 | na (1) | na (1) | na (1) | na (1) | na (1) | na (1) | ||
| Wild boar | 1/15 | 6.7 | 0/1 | 0.0 | na (1) | na (1) | na (1) | na (1) | na (1) | na (1) | ||||
| Ixodes ricinus | Habitat | Site | Total Pos | B. afzelii (n/%) | B. burgdorferi Sensu Stricto (n/%) | B. garinii (n/%) | B. valaisiana (n/%) | B. spielmanii (n/%) | Uncultured Borrelia (n/%) | B. bavariensis (n/%) | Borrelia sp. (n/%) | B. garinii/B. bavariensis (n/%) | Unknown (n/%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Questing ticks | Urban | M2 | 112 | 71/63.4 | 13/11.6 | 13/11.6 | 5/4.5 | 2/1.8 | 0 | 2/1.8 | 0 | 0 | 6/5.4 |
| M3 | 80 | 37/46.3 | 25/31.3 | 4/5.0 | 1/1.3 | 5/6.3 | 6/7.5 | 0 | 0 | 0 | 2/2.5 | ||
| R1 | 106 | 66/62.3 | 6/5.7 | 0 | 7/6.6 | 22/20.8 | 0 | 2/1.9 | 0 | 0 | 3/2.8 | ||
| B | 42 | 21/50.0 | 15/35.7 | 1/2.4 | 4/9.5 | 0 | 0 | 0 | 0 | 0 | 1/2.4 | ||
| Total | 340 | 195/57.4 | 59/17.4 | 18/5.3 | 17/5.0 | 29/8.5 | 6/1.8 | 4/1.2 | 0 | 0 | 12/3.5 | ||
| Pasture | K | 102 | 30/29.4 | 6/5.9 | 39/38.2 | 14/13.7 | 0 | 1/1.0 | 0 | 0 | 4/3.9 | 8/7.8 | |
| Natural | T | 27 | 3/11.1 | 10/37.0 | 5/18.5 | 4/14.8 | 0 | 1/3.7 | 0 | 1/3.7 | 0 | 3/11.1 | |
| All sites | Total | 469 | 228/48.6 | 75/16.0 | 62/13.2 | 35/7.5 | 29/6.2 | 8/1.7 | 4/0.9 | 1/0.2 | 4/0.9 | 23/4.9 | |
| Cattle | Pasture | K | 5 | 2/40.0 | 0 | 2/40.0 | 1/20.0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Roe deer | Natural | T | 16 | 4/25.0 | 2/12.5 | 7/43.8 | 0 | 0 | 0 | 0 | 1/6.3 | 0 | 2/12.5 |
| Wild boar | 1 | 0 | 1/100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Răileanu, C.; Silaghi, C.; Fingerle, V.; Margos, G.; Thiel, C.; Pfister, K.; Overzier, E. Borrelia burgdorferi Sensu Lato in Questing and Engorged Ticks from Different Habitat Types in Southern Germany. Microorganisms 2021, 9, 1266. https://doi.org/10.3390/microorganisms9061266
Răileanu C, Silaghi C, Fingerle V, Margos G, Thiel C, Pfister K, Overzier E. Borrelia burgdorferi Sensu Lato in Questing and Engorged Ticks from Different Habitat Types in Southern Germany. Microorganisms. 2021; 9(6):1266. https://doi.org/10.3390/microorganisms9061266
Chicago/Turabian StyleRăileanu, Cristian, Cornelia Silaghi, Volker Fingerle, Gabriele Margos, Claudia Thiel, Kurt Pfister, and Evelyn Overzier. 2021. "Borrelia burgdorferi Sensu Lato in Questing and Engorged Ticks from Different Habitat Types in Southern Germany" Microorganisms 9, no. 6: 1266. https://doi.org/10.3390/microorganisms9061266
APA StyleRăileanu, C., Silaghi, C., Fingerle, V., Margos, G., Thiel, C., Pfister, K., & Overzier, E. (2021). Borrelia burgdorferi Sensu Lato in Questing and Engorged Ticks from Different Habitat Types in Southern Germany. Microorganisms, 9(6), 1266. https://doi.org/10.3390/microorganisms9061266