
Rhodotorula kratochvilovae CCY 20-2-26—The Source of Multifunctional Metabolites


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Yeast Strain and Media for Optimizing the Culture Conditions
2.3. Preparation of Yeast Biomass for the Glucan, Lipid and Carotenoids Analysis
2.4. Analysis of Glucans by Yeast and Mushroom β-glucan Assay Kit
2.5. Total Lipid Content and Analysis of Fatty Acid Profile
2.6. Analysis of Carotenoids
2.7. Isolation and Characterization of Extracellular Glycolipids
3. Results
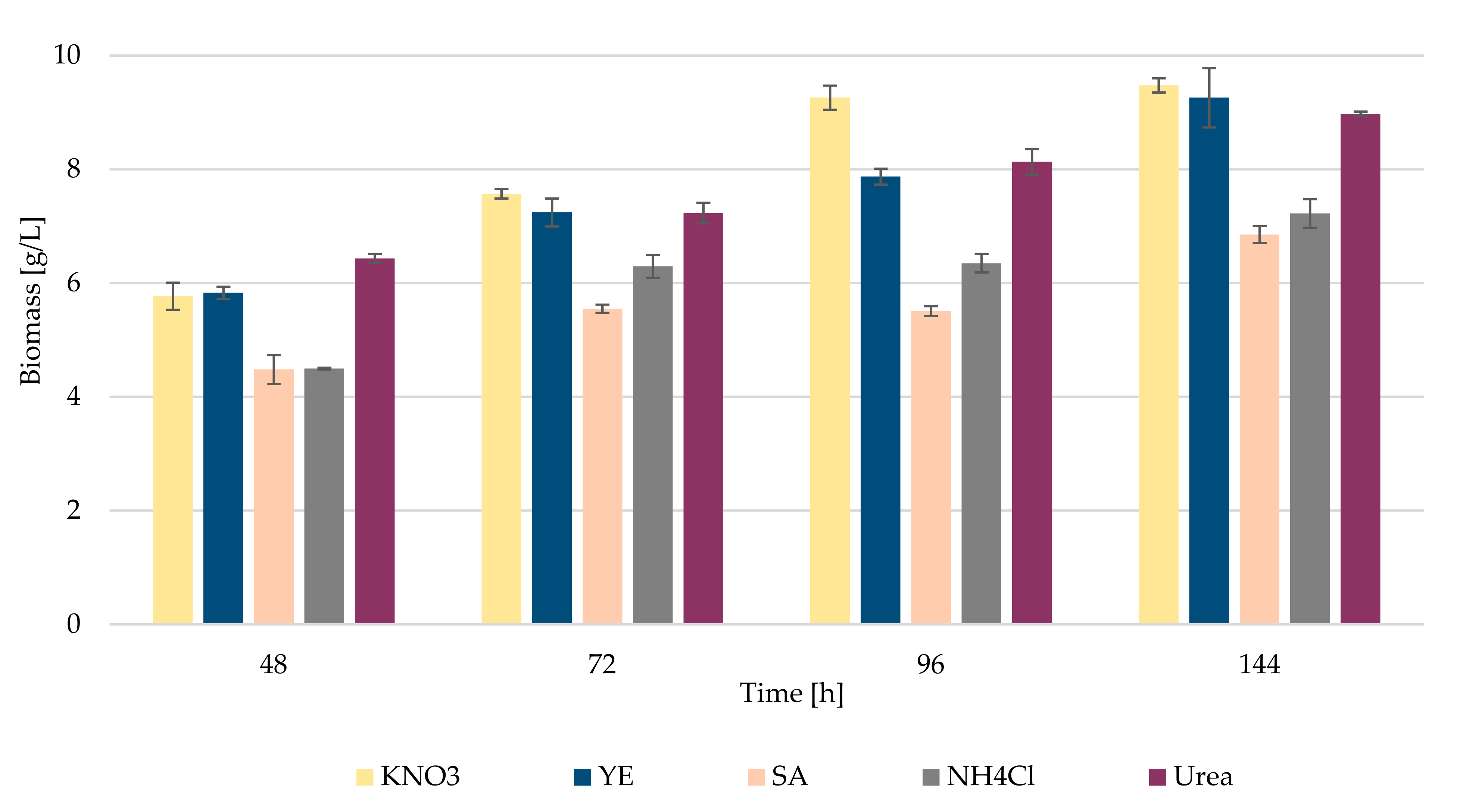
3.1. Biomass and Biochemical Profile of R. kratochvilovae CCY 20-2-26 Growth on Diverse Nitrogen Sources
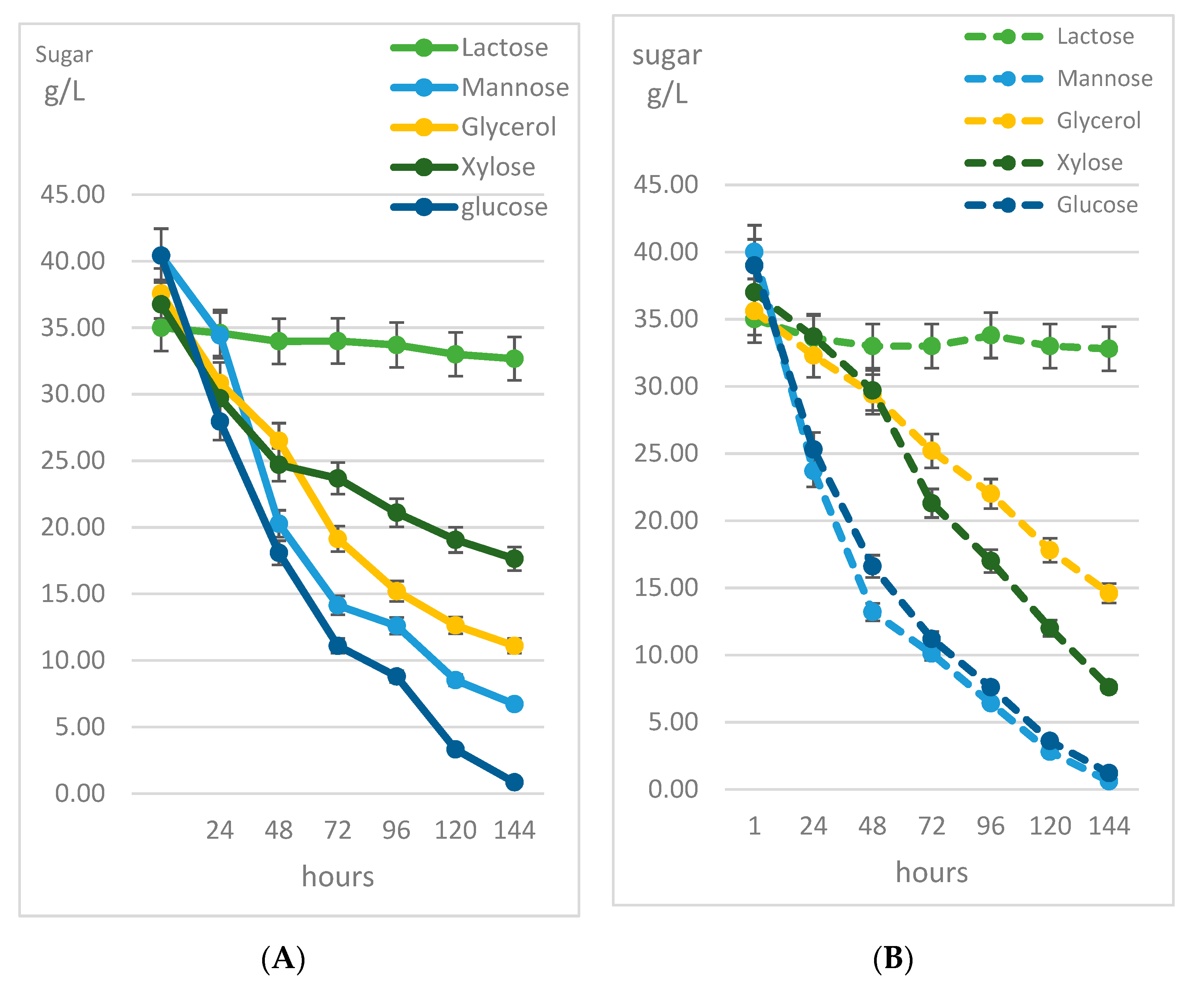
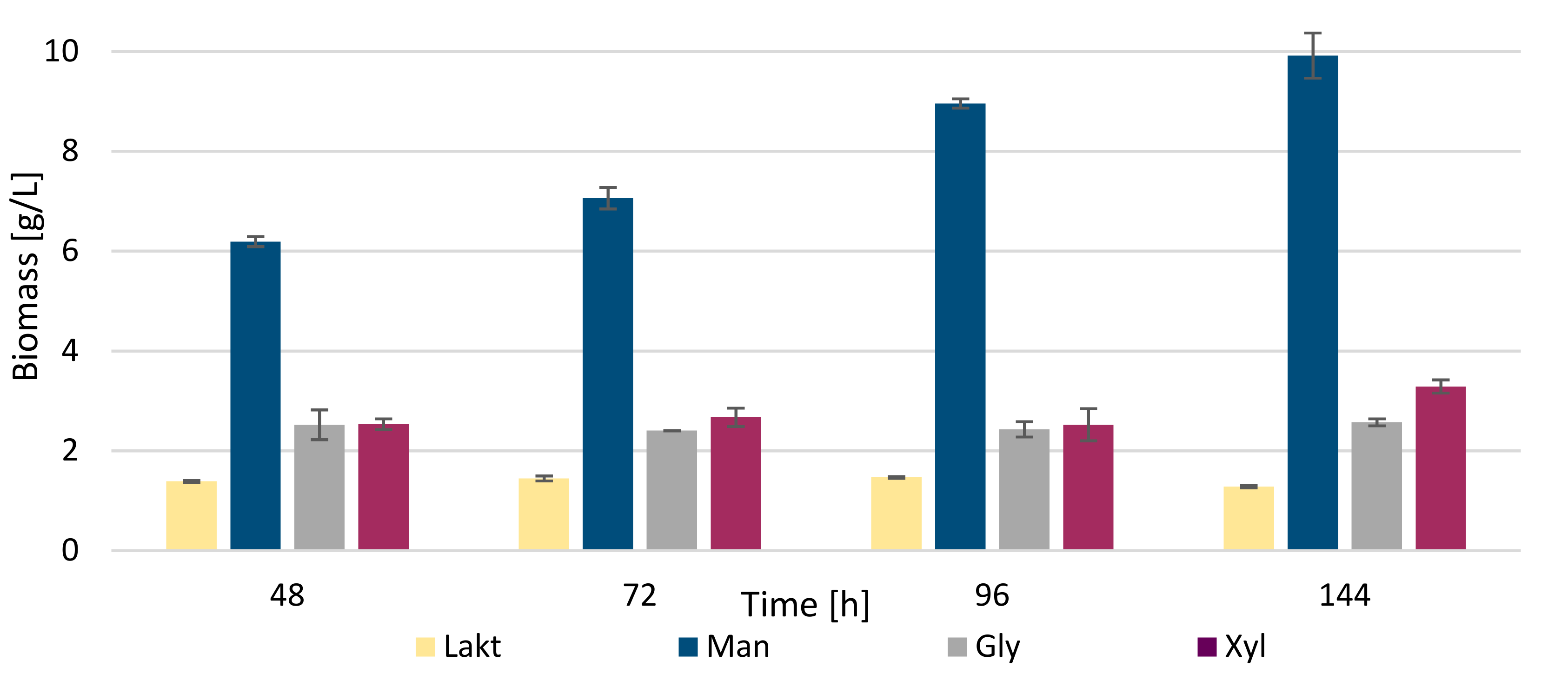
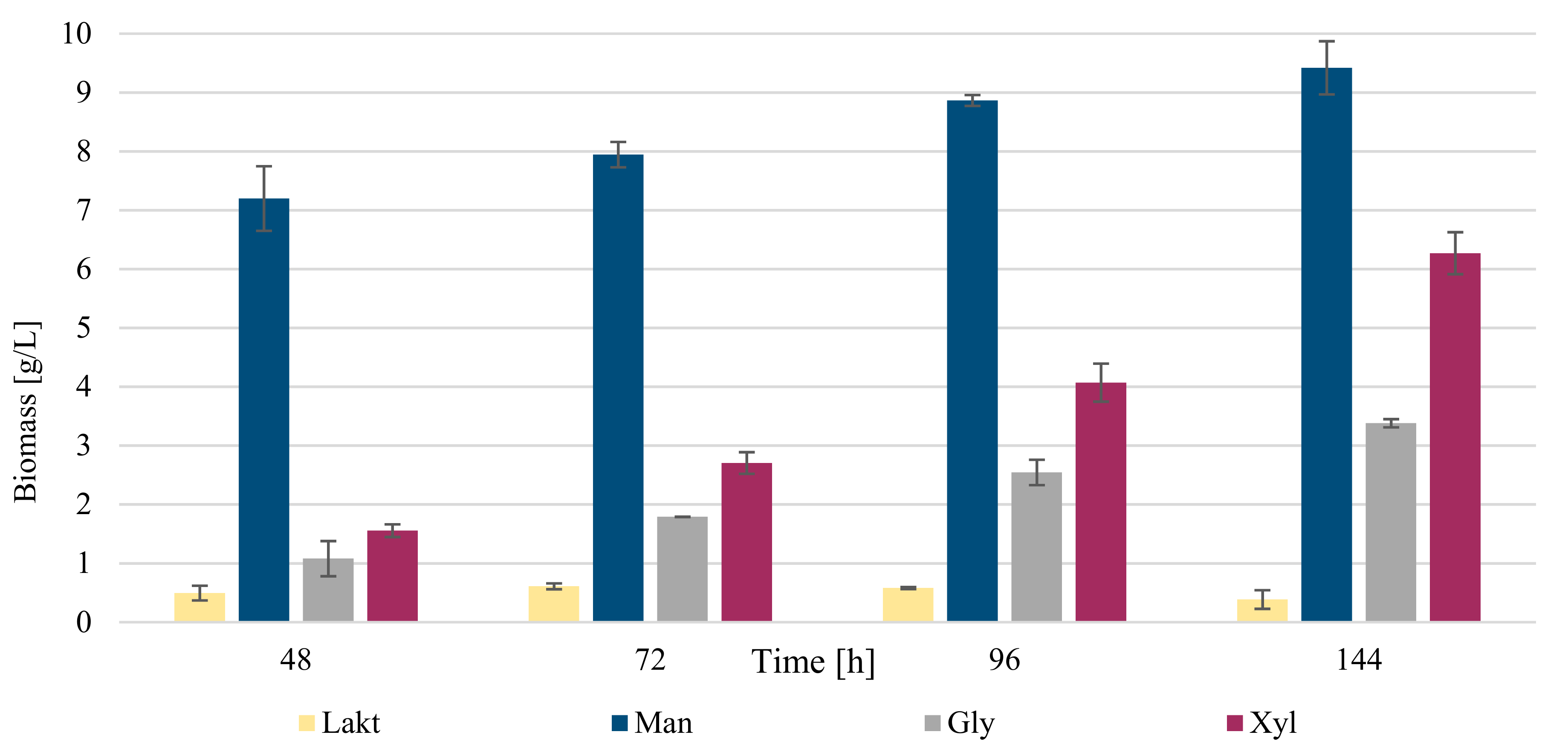
3.2. Biochemical Profile of R. kratochvilovae CCY 20-2-26 Growth on Diverse Carbon Sources
3.3. Influence of Different Carbon: Nitrogen Ratio on Growth and Metabolism of Rhodotorula kratochvilovae CCY 20-2-26
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, J.; Perez-Pinera, P.; Lowenhaupt, K.; Wu, M.R.; Purcell, O.; Fuente-Nunez, C.; Lu, T.K. Versatile and on-demand biologics co-production in yeast. Nat. Commun. 2018, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Xu, J.Z.; Liu, Z.M.; Qian, H.; Zhang, W.G. Co-production of microbial oil and exopolysaccharides by the oleaginous yeast Sporidiobolus pararoseus grown in fed-batch culture. RSC Adv. 2018, 8, 3348–3356. [Google Scholar] [CrossRef] [Green Version]
- Kot, A.M.; Blazejak, S.; Kieliszek, M.; Gientka, I.; Piwowarek, K.; Brzezinska, R. Production of lipids and carotenoids by Rhodotorula gracilis ATCC 10788 yeast in a bioreactor using low-cost wastes. Biocatal. Agric. Biotechnol. 2020, 26, 101634. [Google Scholar] [CrossRef]
- Pi, H.W.; Anandharaj, M.; Kao, Y.Y.; Lin, Y.J.; Chang, J.J.; Li, W.H. Engineering the oleaginous red east Rhodotorula glutinis for simultaneous β-carotene and cellulase production. Sci. Rep. 2018, 8, 10850. [Google Scholar] [CrossRef] [PubMed]
- Money, N.P. Fungal diversity. In The Fungi, 3rd ed.; Watkinson, S.C., Boddy, L., Money, N.P., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 1–36. [Google Scholar]
- Kot, A.M.; Blazejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula glutinis—Potential source of lipids, carotenoids, and enzymes for use in industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Yang, J.M.; Xu, X.; Zhang, L.; Nie, Q.J.; Xian, M. Biodiesel production from oleaginous microorganisms. Renew. Energy 2009, 34, 1–5. [Google Scholar] [CrossRef]
- Marova, I.; Carnecka, M.; Halienova, A.; Breierova, E.; Koci, R. Production of carotenoid-/ergosterol-supplemented biomass by red yeast Rhodotorula glutinis grown under external stress. Food Technol. Biotechnol. 2010, 48, 56–61. [Google Scholar]
- Lario, L.D.; Pillaca-Pullo, O.S.; Sette, L.D.; Converti, A.; Casati, P.; Spampinato, C.; Pessoa, A. Optimization of protease production and sequence analysis of the purified enzyme from the cold adapted yeast Rhodotorula mucilaginosa CBMAI 1528. Biotechnol. Rep. 2020, 28, e00546. [Google Scholar] [CrossRef]
- Li, H.; Huang, L.; Zhang, Y.; Yan, Y. Production, characterization and immunomodulatory activity of an extracellular polysaccharide from Rhodotorula mucilaginosa YL-1 isolated from sea salt field. Mar. Drugs 2020, 18, 595. [Google Scholar] [CrossRef] [PubMed]
- Byrtusova, D.; Shapaval, V.; Holub, J.; Rapta, M.; Szotkowski, M.; Kohler, A.; Marova, I. Revealing the potential of lipid and β-glucans coproduction in Basidiomycetes yeast. Microorganisms 2020, 8, 1034. [Google Scholar] [CrossRef]
- Mata-Gomez, L.C.; Montanez, J.C.; Mendez-Zavala, A.; Aguilar, C.N. Biotechnological production of carotenoids by yeast: An overview. Microb. Cell Fact. 2014, 13, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Aspects Med. 2005, 6, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Chew, B.P.; Park, J.S. Carotenoid action on the immune response. J. Nutr. 2004, 1, 257S–261S. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as a source of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef]
- Kot, A.M.; Blazejak, S.; Gientka, I.; Kieliszek, M.; Brys, J. Torulene and torularhodin: “new” fungal carotenoids for industry? Microb. Cell Fact. 2018, 17, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiukova, I.A.; Brandenburg, J.; Blomqvist, J.; Sampels, S.; Mikkelsen, N.; Skaugen, M.; Arntzen, M.; Nielsen, J.; Sandgren, M.; Kerkhoven, E.J. Proteome analysis of xylose metabolism in Rhodotorula toruloides during lipid production. Biotechnol. Biofuels 2019, 12, 137. [Google Scholar] [CrossRef] [Green Version]
- Stier, H.; Ebbeskotte, V.; Gruenwald, J. Immune-modulatory effects of dietary yeast beta-1,3/1,6-D-glucan. Nutr. J. 2014, 13, 1. [Google Scholar]
- Rop, O.; Mlcek, J.; Jurikova, T. Beta-glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar]
- Sima, P.; Vannucci, L.; Vetvicka, V.; Pradhan, B.B.; Chatterjee, S.; Nilson, L.; Wang, W. Β-glucans and cholesterol (Review). Int. J. Mol. Med. 2018, 52, 275–288. [Google Scholar]
- Velasques, G.L.; de Lima, O.F.; Boffo, E.F.; Santos, J.D.G.; da Silva, B.C.; de Assis, S.A. Extraction optimization and antinociceptive activity of (1→3)-β-d-glucan from Rhodotorula mucilaginosa: An overview. Carbohydr. Polym. 2014, 105, 293–299. [Google Scholar] [CrossRef]
- Gründemann, C.; Garcia-Kaufer, M.; Sauer, B.; Scheer, R.; Merdivan, S.; Bettin, P.; Huber, R.; Lindequist, U. Comparative chemical and biological investigations of β-glucan-containing products from shiitake mushrooms: An overview. J. Funct. Foods 2015, 18, 692–702. [Google Scholar] [CrossRef]
- Synytsya, A.; Mickova, K.; Synytsya, A.; Jablonsky, I.; Spevacek, J.; Erban, V.; Kovariková, E.; Copiková, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Szotkowski, M.; Byrtusova, D.; Haronikova, A.; Vysoka, M.; Rapta, M.; Shapaval, V.; Marova, I. Study of metabolic adaptation of red yeast to waste animal fat substrate. Microorganisms 2019, 7, 578. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Mao, W.; Wang, X.; Li, F.; Wang, J.; Chi, Z.; Chi, Z.; Liu, G. Efficient simultaneous production of extracellular polyol esters of fatty acids and intracellular lipids from inulin by a deep-sea yeast Rhodotorula paludigena P4R5. Microb. Cell Fact. 2019, 18, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunwald, T.; Schwemmlein, L.; Graeff-Hönninger, S.; French, W.T.; Hernandez, R.; Holmes, W.E.; Claupein, W. Effect of different C/N ratios on carotenoid and lipid production by Rhodotorula glutinis. Appl. Microbiol. 2013, 97, 6581–6588. [Google Scholar] [CrossRef] [PubMed]
- Jiru, T.M.; Groenewald, M.; Pohl, C.; Steyn, L.; Kiggundu, N.; Abate, D. Optimization of cultivation conditions for biotechnological production of lipid by Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 for biodiesel preparation. 3Biotech 2017, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dourou, M.; Aggeli, D.; Papanikolaou, S.; Aggelis, G. Critical steps in carbon metabolism affecting lipid accumulation and their regulation in oleaginous microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 2509–2523. [Google Scholar] [CrossRef]
- Tchakouteu, S.S.; Chatzifragkou, A.; Kalantzi, O.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Oleaginous yeast Cryoptococcus curvatus exhibits interplay between biosynthesis of intracellular sugars and lipids. Eur. J. Lipid Sci. Technol. 2015, 117, 657–672. [Google Scholar] [CrossRef]
- Diamantopoulou, P.; Stoforos, N.G.; Xenopoulos, E.; Sarrisa, D.; Psarianos, D.; Philippoussis, A.; Papanikolaou, S. Lipid production by Cryptococcus curvatus growing on commercial xylose and subsequent valorization of fermentation waste-waters for the production of edible and medicinal mushrooms. Biochem. Engineer. J. 2020, 162, 107706. [Google Scholar] [CrossRef]
- Tchakouteu, S.S.; Kalantzi, O.; Gardeli, C.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Lipid production by yeasts growing on biodiesel-derived crude glycerol: Strain selection and impact of substrate concentration on the fermentation efficiency. J. Appl. Microbiol. 2015, 118, 911–927. [Google Scholar] [CrossRef]
- Sarantou, S.; Stoforos, N.G.; Kalantzi, O.; Papanikolaou, S. Biotechnological valorization of biodiesel-derived glycerol: Trials with the non-conventional yeasts Yarrowia lipolytica and Rhodosporidium sp. Carb. Resour. Conver. 2021, 4, 61–75. [Google Scholar]
- Kostovova, I.; Byrtusova, D.; Rapta, M.; Babak, V.; Marova, I. The variability of carotenoid pigments and fatty acids produced by some yeasts within Sporidiobolales and Cystofilobasidiales. Chem. Papers 2021, 75, 3353–3362. [Google Scholar] [CrossRef]
- Garay, L.A.; Sitepu, I.R.; Cajka, T.; Fiehn, O.; Cathcart, E.; Fry, R.W.; Kanti, A.; Nugroho, A.J.; Faulina, S.A.; Stephanandra, S.; et al. Discovery of synthesis and secretion of polyol esters of fatty acids by four basidiomycetous yeast species in the order Sporidiobolales. J. Ind. Microbiol. Biotechnol. 2017, 44, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Guerfali, M.; Ayadi, I.; Mohamed, N.; Ayadi, W.; Belghith, H.; Bronze, M.R.; Ribeiro, M.H.L.; Gargouri, A. Triacylglycerols accumulation and glycolipids secretion by the oleaginous yeast Rhodotorula babjevae Y-SL7: Structural identification and biotechnological applications. Bioresour. Technol. 2019, 273, 326–334. [Google Scholar] [CrossRef]
- Garay, L.A.; Sitepu, I.R.; Cajka, T.; Cathcart, E.; Fiehn, O.; German, J.B.; Block, D.E.; Boundy-Mills, K.L. Simultaneous production of intracellular triacylglycerols and extracellular polyol esters of fatty acids by Rhodotorula babjevae and Rhodotorula aff. paludigena. J. Ind. Microbiol. Biotechnol. 2017, 44, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Blazejak, S.; Kieliszek, M.; Gientka, I.; Brys, J. Simultaneous production of lipids and carotenoids by the red yeast Rhodotorula from waste glycerol fraction and potato wastewater. Appl. Biochem. Biotechnol. 2019, 189, 589–607. [Google Scholar] [CrossRef] [PubMed]
- Garay, L.; Sitepu, I.; Cajka, T.; Xu, J.; Teh, H.E.; German, J.B.; Pan, Z.; Dungan, S.R.; Block, D.E.; Boundy-Mills, K. L Extracellular fungal polyol lipids: A new class of potential high value lipids. Biotechnol. Adv. 2018, 36, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Aimanianda, V.; Clavaud, C.; Sinemel, C.; Fontaine, T.; Delepierre, M.; Latgé, J.P. Cell wall β-(1,6)-glucan of Saccharomyces cerevisiae: Structural characterization and in situ synthesis. J. Biol. Chem. 2009, 284, 13401–13412. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
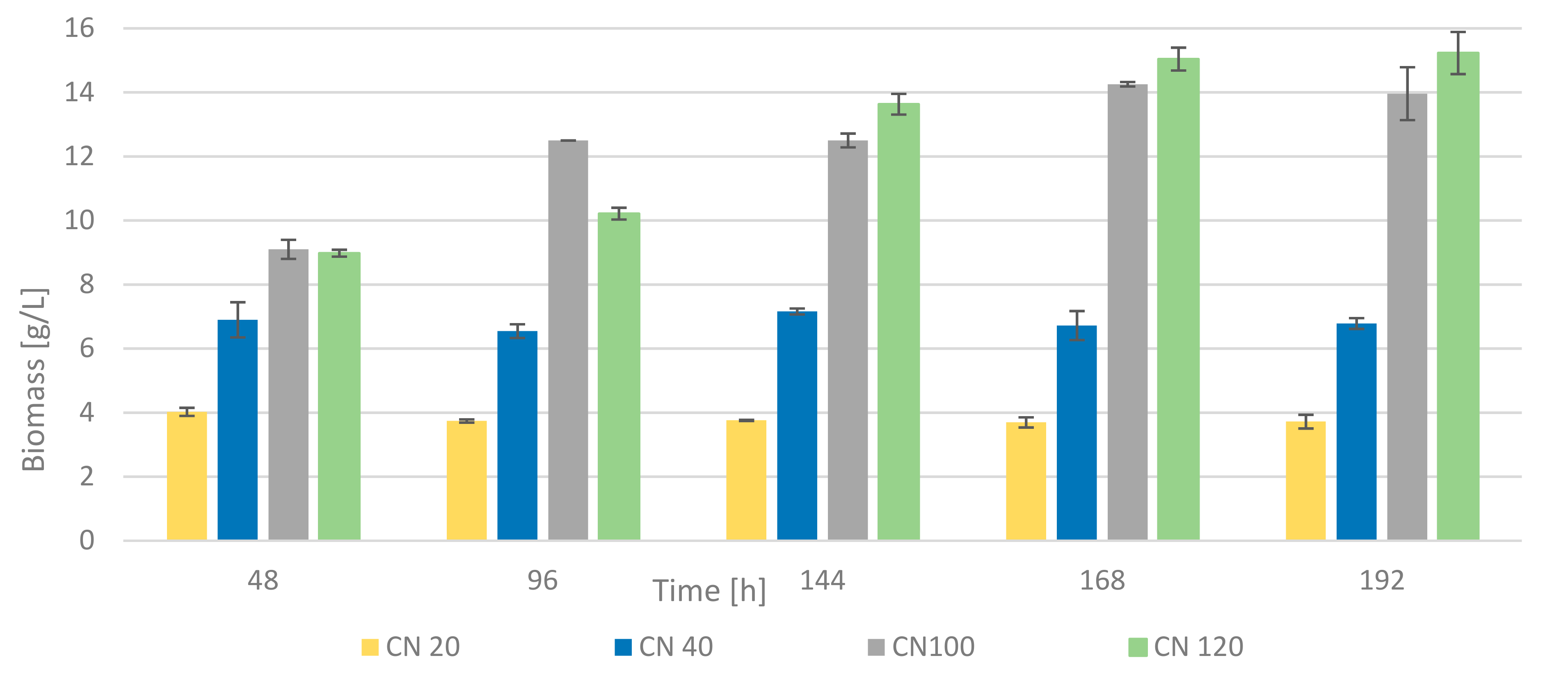{kind=link}
| Component | g/L |
| Glucose | 40.42 |
| KH2PO4 | 4 |
| MgSO4·7H2O | 0.7 |
| Nitrogen sources | g/L |
| Yeast extract | 2 |
| KNO3 | 1.52 |
| NH4Cl | 0.80 |
| (NH2)2SO4 | 0.9906 |
| Urea | 0.45 |
| Component 1 [g/L] | Component 2 [g/L] |
|---|---|
| Urea—0.45 | KNO3—1.52 |
| KH2PO4—4 | KH2PO4—4 |
| MgSO4·7H2O—0.7 | MgSO4·7H2O—0.7 |
| Carbon sources | g/L |
| Mannose | 40.42 |
| Lactose | 34.94 |
| Glycerol | 37.57 |
| Xylose | 36.75 |
| Glucose | 40.42 |
| Retention Time (min) | Gradient (°C/min) | Target Temperature (°C) | Hold (min) |
|---|---|---|---|
| 0 | - | - | - |
| 1 | 0 | 80 | 1 |
| 5 | 15 | 140 | 0 |
| 21.7 | 3 | 190 | 0 |
| 25.5 | 25 | 260 | 1 |
| 25.5 | STOP | - | - |
| Column | Kinetex, EVO 150 × 4.6 mm, 2.6 µm; Phenomenex |
|---|---|
| Volume of the sample | 20 µL |
| Elution | Gradient 0–13 min: from 100% A to 100% B linearly 13–19 min: 100% B 19–20 min: from 100% B to 100% A linearly 20–25 min: 100% A |
| PDA | 285 (ergosterol), 435, 450 and 680 nm (carotenoids) |
| Temperature | 25 °C |
| Time of analysis | 25 min |
| Nitrogen | Metabolite | 48 h | 72 h | 96 h | 144 h |
|---|---|---|---|---|---|
| KNO3 | Total lipids (% of DCW) | 30.2 ± 1.9 | 42.5 ± 2.1 | 59.6 ± 1.3 | 49.8 ± 2.5 |
| Total carotenoids (mg/g) | 1.00 ± 0.27 | 1.25 ± 0.13 | 1.36 ± 0.14 | 2.48 ± 0.13 | |
| Extr. glycolipids (g/L) | 0.62 ± 0.02 | 1.73 ± 0.09 | 1.51 ± 0.20 | 1.83 ± 0.07 | |
| Beta-glucan (% of DCW) | 22.05 ± 2.21 | 18.31 ± 1.83 | 15.24 ± 1.24 | 14.39 ± 0.77 | |
| YE | Total lipids (% of DCW) | 37.0 ± 1.0 | 43.8 ± 1.8 | 58.8 ± 1.5 | 56.2 ± 1.2 |
| Total carotenoids (mg/g) | 1.19 ± 0.11 | 1.08 ± 0.05 | 1.25 ± 0.03 | 1.16 ± 0.09 | |
| Extr. glycolipids (g/L) | 0.51 ± 0.04 | 0.75 ± 0.05 | 1.17 ± 0.20 | 1.28 ± 0.08 | |
| Beta-glucan (% of DCW) | 24.99 ± 1.69 | 21.43 ± 1.57 | 16.70 ± 1.33 | 12.62 ± 1.11 | |
| SA | Total lipids (% of DCW) | 25.1 ± 2.0 | 34.6 ± 2.4 | 37.2 ± 0.5 | 27.0 ± 2.1 |
| Total carotenoids (mg/g) | 1.30 ± 0.02 | 1.48 ± 0.16 | 1.61 ± 0.03 | 1.38 ± 0.06 | |
| Extr. glycolipids (g/L) | 0.12 ± 0.01 | 0.05 ± 0.02 | 0.08 ± 0.03 | 0.07 ± 0.02 | |
| Beta-glucan (% of DCW) | 12.41 ± 1.02 | 13.98 ± 1.04 | 16.54 ± 1.00 | 17.60 ± 1.84 | |
| NH4Cl | Total lipids (% of DCW) | 30.5 ± 2.4 | 43.6 ± 2.1 | 41.3 ± 1.2 | 38.8 ± 3.0 |
| Total carotenoids (mg/g) | 1.34 ± 0.10 | 1.54 ± 0.09 | 1.52 ± 0.16 | 1.52 ± 0.04 | |
| Extr. glycolipids (g/L) | 0.13 ± 0.01 | 0.31 ± 0.05 | 0.32 ± 0.09 | 0.23 ± 0.08 | |
| Beta-glucan (% of DCW) | 14.80 ± 0.81 | 15.71 ± 0.66 | 13.33 ± 0.85 | 11.30 ± 0.75 | |
| Urea | Total lipids (% of DCW) | 31.3 ± 1.1 | 42.2 ± 2.0 | 56.8 ± 1.2 | 51.7 ± 3.1 |
| Total carotenoids (mg/g) | 0.97 ± 0.01 | 1.04 ± 0.00 | 1.14 ± 0.46 | 2.83 ± 0.21 | |
| Extr. glycolipids (g/L) | 0.58 ± 0.02 | 0.88 ± 0.05 | 1.51 ± 0.07 | 1.57 ± 0.06 | |
| Beta-glucan (% of DCW) | 20.94 ± 1.25 | 20.15 ± 1.30 | 14.67 ± 0.59 | 16.93 ± 1.13 |
| Carbon | Metabolite | 48 h | 72 h | 96 h | 144 h |
|---|---|---|---|---|---|
| Lactose | Total lipids (% of DCW) | 9.0 ± 2.3 | 8.6 ± 1.6 | 8.9 ± 1.0 | 9.2 ± 2.1 |
| Total carotenoids (mg/g) | 0.32 ± 0.08 | 0.87 ± 0.13 | 0.82 ± 0.19 | 0.70 ± 0.10 | |
| Extr. glycolipids (g/L) | 0.10 ± 0.01 | 0.01 ± 0.00 | 0.03 ± 0.02 | 0.03 ± 0.01 | |
| Beta-glucan (% of DCW) | - | - | - | - | |
| Mannose | Total lipids (% of DCW) | 37.0 ± 2.3 | 32.4 ± 2.2 | 55.8 ± 3.8 | 51.1 ± 4.1 |
| Total carotenoids (mg/g) | 1.01 ± 0.18 | 1.02 ± 0.20 | 1.13 ± 0.22 | 1.39 ± 0.19 | |
| Extr. glycolipids (g/L) | 0.51 ± 0.02 | 0.61 ± 0.05 | 1.06 ± 0.08 | 1.42 ± 0.10 | |
| Beta-glucan (% of DCW) | 13.75 ± 0.99 | 10.59 ± 0.76 | 10.28 ± 1.83 | 9.87 ± 0.67 | |
| Glycerol | Total lipids (% of DCW) | 14.9 ± 3.6 | 16.8 ± 2.8 | 15.5 ± 1.4 | 17.2 ± 2.3 |
| Total carotenoids (mg/g) | 0.97 ± 0.11 | 1.14 ± 0.23 | 1.18 ± 0.25 | 1.21 ± 0.18 | |
| Extr. glycolipids (g/L) | 0.14 ± 0.00 | 0.09 ± 0.01 | 0.23 ± 0.02 | 0.10 ± 0.03 | |
| Beta-glucan (% of DCW) | 3.83 ± 0.54 | 4.19 ± 0.67 | 5.00 ± 0.37 | 5.37 ± 0.80 | |
| Xylose | Total lipids (% of DCW) | 15.6 ± 0.1 | 15.7 ± 2.5 | 19.2 ± 0.9 | 20.3 ± 2.4 |
| Total carotenoids (mg/g) | 0.97 ± 0.22 | 1.47 ± 0.32 | 1.13 ± 0.21 | 1.52 ± 0.30 | |
| Extr. glycolipids (g/L) | 0.10 ± 0.01 | 0.12 ± 0.02 | 0.15 ± 0.05 | 0.03 ± 0.01 | |
| Beta-glucan (% of DCW) | 2.96 ± 0.39 | 8.28 ± 1.13 | 7.59 ± 0.91 | 7.04 ± 0.76 |
| Carbon | Metabolite | 48 h | 72 h | 96 h | 144 h |
|---|---|---|---|---|---|
| Lactose | Total lipids (% of DCW) | 12.5 ± 0.9 | 16.9 ± 0.6 | 18.7 ± 1.1 | 20.2 ± 0.9 |
| Total carotenoids (mg/g) | 0.53 ± 0.10 | 0.68 ± 0.11 | 0.78 ± 0.18 | 0.40 ± 0.09 | |
| Extr. glycolipids (g/L) | 0.50 ± 0.11 | 0.07 ± 0.02 | 0.05 ± 0.02 | 0.02 ± 0.01 | |
| Beta-glucan (% of DCW) | - | - | - | - | |
| Mannose | Total lipids (% of DCW) | 34.1 ± 2.3 | 40.0 ± 2.9 | 51.0 ± 2.5 | 57.5 ± 3.1 |
| Total carotenoids (mg/g) | 1.10 ± 0.11 | 1.11 ± 0.15 | 1.31 ± 0.20 | 2.21 ± 0.16 | |
| Extr. glycolipids (g/L) | 0.83 ± 0.17 | 1.35 ± 0.24 | 1.68 ± 0.33 | 1.99 ± 0.36 | |
| Beta-glucan (% of DCW) | 17.13 ± 1.24 | 15.62 ± 1.67 | 18.14 ± 0.58 | 19.46 ± 1.85 | |
| Glycerol | Total lipids (% of DCW) | 17.3 ± 1.9 | 18.0 ± 1.8 | 18.1 ± 2.0 | 26.7 ± 2.1 |
| Total carotenoids (mg/g) | 0.66 ± 0.12 | 0.90 ± 0.09 | 1.29 ± 0.14 | 2.72 ± 0.24 | |
| Extr. glycolipids (g/L) | 0.05 ± 0.01 | 0.06 ± 0.01 | 0.08 ± 0.02 | 0.13 ± 0.03 | |
| Beta-glucan (% of DCW) | 6.61 ± 0.49 | 5.36 ± 0.41 | 3.08 ± 0.67 | 2.69 ± 0.34 | |
| Xylose | Total lipids (% of DCW) | 13.6 ± 1.9 | 15.2 ± 1.2 | 22.4 ± 1.7 | 34.9 ± 2.2 |
| Total carotenoids (mg/g) | 0.72 ± 0.14 | 1.12 ± 0.20 | 1.13 ± 0.11 | 1.97 ± 0.23 | |
| Extr. glycolipids (g/L) | 0.05 ± 0.02 | 0.22 ± 0.04 | 0.22 ± 0.05 | 0.25 ± 0.03 | |
| Beta-glucan (% of DCW) | 14.08 ± 1.05 | 13.58 ± 0.81 | 11.85 ± 1.45 | 10.00 ± 1.27 |
| C/N Ratio | Metabolite | 48 h | 96 h | 144 h | 168 h |
|---|---|---|---|---|---|
| 20:1 | Total lipids (% of DCW) | 16.2 ± 1.5 | 16.4 ± 1.5 | 14.3 ± 4.2 | 14.2 ± 1.5 |
| Extr. glycolipids (g/L) | 0.10 ± 0.02 | 0.06 ± 0.01 | 0.19 ± 0.03 | 0.16 ± 0.01 | |
| Total carotenoids (mg/g) | 1.33 ± 0.10 | 0.72 ± 0.06 | 0.90 ± 0.03 | 0.85 ± 0.08 | |
| Beta-glucan (% of DCW) | 10.85 ± 0.68 | 9.30 ± 0.23 | 6.54 ± 0.25 | 7.38 ± 0.63 | |
| 40:1 | Total lipids (% of DCW) | 33.3 ± 1.32 | 40.7 ± 1.88 | 28.3 ± 3.6 | 26.2 ± 1.3 |
| Extr. glycolipids (g/L) | 0.32 ± 0.06 | 0.35 ± 0.04 | 0.46 ± 0.02 | 0.48 ± 0.05 | |
| Total carotenoids (mg/g) | 1.07 ± 0.12 | 0.90 ± 0.20 | 1.10 ± 0.08 | 1.03 ± 0.11 | |
| Beta-glucan (% of DCW) | 18.45 ± 1.26 | 15.48 ± 1.41 | 13.33 ± 1.19 | 12.83 ± 1.23 | |
| 100:1 | Total lipids (% of DCW) | 40.1 ± 2.8 | 53.5 ± 2.9 | 61.3 ± 3.1 | 62.1 ± 2.5 |
| Extr. glycolipids (g/L) | 0.95 ± 0.11 | 1.35 ± 0.15 | 2.21 ± 0.06 | 2.25 ± 0.13 | |
| Total carotenoids (mg/g) | 1.08 ± 0.14 | 0.76 ± 0.12 | 1.48 ± 0.21 | 1.57 ± 0.22 | |
| Beta-glucan (% of DCW) | 19.75 ± 0.91 | 14.10 ± 1.33 | 12.04 ± 0.42 | 10.25 ± 0.98 | |
| 120:1 | Total lipids (% of DCW) | 42.2 ± 3.86 | 64.5 ± 4.32 | 65.3 ± 3.20 | 66.7 ± 1.50 |
| Extr. glycolipids (g/L) | 0.9 ± 0.03 | 1.40 ± 0.05 | 2.19 ± 0.06 | 2.42 ± 0.08 | |
| Total carotenoids (mg/g) | 1.41 ± 0.15 | 0.76 ± 0.12 | 1.21 ± 0.09 | 1.35 ± 0.11 | |
| Beta-glucan (% of DCW) | 20.75 ± 0.85 | 12.08 ± 1.49 | 14.76 ± 0.95 | 11.33 ± 1.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byrtusová, D.; Szotkowski, M.; Kurowska, K.; Shapaval, V.; Márová, I. Rhodotorula kratochvilovae CCY 20-2-26—The Source of Multifunctional Metabolites. Microorganisms 2021, 9, 1280. https://doi.org/10.3390/microorganisms9061280
Byrtusová D, Szotkowski M, Kurowska K, Shapaval V, Márová I. Rhodotorula kratochvilovae CCY 20-2-26—The Source of Multifunctional Metabolites. Microorganisms. 2021; 9(6):1280. https://doi.org/10.3390/microorganisms9061280
Chicago/Turabian StyleByrtusová, Dana, Martin Szotkowski, Klára Kurowska, Volha Shapaval, and Ivana Márová. 2021. "Rhodotorula kratochvilovae CCY 20-2-26—The Source of Multifunctional Metabolites" Microorganisms 9, no. 6: 1280. https://doi.org/10.3390/microorganisms9061280
APA StyleByrtusová, D., Szotkowski, M., Kurowska, K., Shapaval, V., & Márová, I. (2021). Rhodotorula kratochvilovae CCY 20-2-26—The Source of Multifunctional Metabolites. Microorganisms, 9(6), 1280. https://doi.org/10.3390/microorganisms9061280