
The Effect of Probiotics on Health Outcomes in the Elderly: A Systematic Review of Randomized, Placebo-Controlled Studies

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Protocols and Registration
2.2. Information Sources and Search Strategy
2.3. Eligibility Criteria
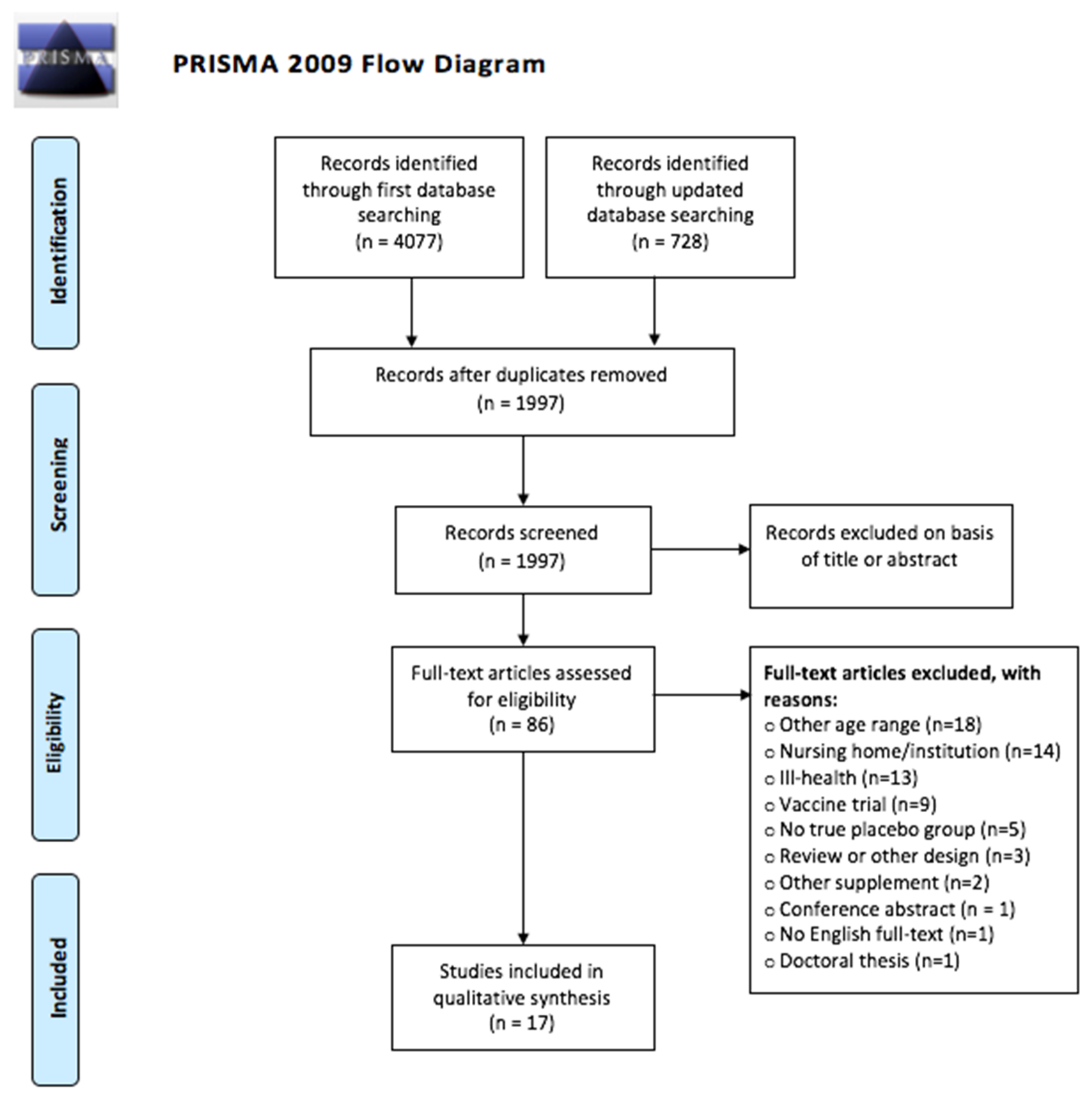
2.4. Study Selection
2.5. Data Extraction
2.6. Risk of Bias and Study Quality Assessment
3. Results
3.1. Assessment of Study Quality
3.2. Microbiota Composition
3.2.1. Probiotics
3.2.2. Synbiotics
3.3. Immune-Related Measurements
3.3.1. Markers of Humoral Immunity
3.3.2. Immune Cell Activity and Population Changes
3.3.3. Common Cold and Other Infection Episodes
3.3.4. Fecal Immune Function
3.4. Digestive Health
3.5. General Well-Being and Cognitive Function
3.6. Lipids and Other Biomarkers
4. Discussion
4.1. The Effects of Probiotic Use as an Intervention in Healthy Elderly
4.1.1. Microbiota Composition
4.1.2. Immune-Related Measurements
4.1.3. Digestive Health
4.1.4. General Well-Being and Cognitive Function
4.1.5. Lipids and Other Biomarkers
4.2. Study Quality of the Research Field and Methodological Considerations within Studies
4.3. Strengths and Limitations of This Systematic Review
4.4. Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, J.E. Human population: The next half century. Science 2003, 302, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbach, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Michaud, M.; Balardy, L.; Moulis, G.; Gaudin, C.; Peyrot, C.; Vellas, B.; Cesari, M.; Nourhashemi, F. Proinflammatory cytokines, aging, and age-related diseases. J. Am. Med. Dir. Assoc. 2013, 14, 877–882. [Google Scholar] [CrossRef]
- Fane, M.; Weeraratna, A.T. How the ageing microenvironment influences tumor progression. Nat. Rev. Cancer 2019, 20, 89–106. [Google Scholar] [CrossRef] [PubMed]
- North, B.J.; Sinclair, D.A. The intersection between aging and cardiovascular disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- WHO (World Health Organisation). Risk Factors of Ill Health among Older People. Health Topics, Life-Course Approach, Healthy Ageing, Data and Statistics. Available online: https://www.euro.who.int/en/health-topics/Life-stages/healthy-ageing/data-and-statistics/risk-factors-of-ill-health-among-older-people (accessed on 1 March 2021).
- López-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungvari, Z.; Taratini, S.; Donato, A.J.; Galvan, V.; Csiszar, A. Mechanisms of vascular aging. Circ. Res. 2018, 123, 849–867. [Google Scholar] [CrossRef] [PubMed]
- Pataky, M.W.; Young, W.F.; Nair, K.S. Hormonal and metabolic changes of aging and the influence of lifestyle modifications. Mayo Clin. Proc. 2021, 96, 788–814. [Google Scholar] [CrossRef] [PubMed]
- Hedden, T.; Gabrieli, J.D.E. Insights into the ageing mind: A view from cognitive neuroscine. Nat. Rev. Neurosci. 2004, 5, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Bosco, N.; Bourdet-Sicard, R.; Capuron, L.; Delzenne, N.; Doré, J.; Franceschi, C.; Lehtinen, M.; Recker, T.; Salvioli, S.; et al. Health relevance of modification of low grade inflammation in aging (inflammageing) and the role of nutrition. Ageing Res. Rev. 2017, 40, 95–119. [Google Scholar] [CrossRef]
- Hutchinson, A.N.; Tingö, L.; Brummer, R.J. The potential effects of probiotics and omega-3 fatty acids on chronic-low grade inflammation. Nutrients 2020, 12, 2402. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.J.; Cusack, S.; O’Sullivan, O.; Greene-Diniz, R.; de Weerd, H.; Flannery, E.; Marchesi, J.R.; Falush, D.; Dinan, T.; Fitzgerald, G.; et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl. Acad. Sci. USA 2011, 108, 4591–4596. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, P.W.; Jeffery, I.B. Gut microbiota and aging. Science 2015, 350, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Biagi, E.; Franceschi, C.; Rampellli, S.; Severgnini, M.; Ostan, R.; Turroni, S.; Consolandi, C.; Quercia, S.; Scurti, M.; Monti, D.; et al. Gut microbiota and extreme longevity. Curr. Biol. 2016, 26, 1480–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drago, L.; Toscano, M.; Rodighiero, V.; De Vecchi, E.; Mogna, G. Cultivable and pyrosequenced fecal microflora in centenarians and young subjects. J. Clin. Gastroenterol. 2012, 46, S81–S84. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Uhr, T. Compartmentalization of the mucosal immune responses to commensal intestinal bacteria. Ann. N. Y. Acad. Sci. 2004, 1029, 36–43. [Google Scholar] [CrossRef]
- Grenham, S.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain-gut-microbe communication in health and disease. Front. Physiol. 2011, 2, 94. [Google Scholar] [CrossRef] [Green Version]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M. A role for the gut microbiota in IBS. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 497–505. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Mayer, E.A. Gut feelings: The emerging biology of gut-brain communication. Nat. Rev. Neurosci. 2011, 12, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress. 2017, 7, 124–136. [Google Scholar]
- Ho, P.; Ross, D.A. More than a gut feeling: The implications of the gut microbiota in psychiatry. Biol. Psychiatry 2017, 81, e35–e37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, P.W.; Claesson, M.J. Gut microbiota: Changes throughout the lifespan from infancy to elderly. Int. Dairy 2010, 20, 281–291. [Google Scholar] [CrossRef]
- Rondanelli, M.; Giacosa, A.; Faliva, M.A.; Perna, S.; Allieri, F.; Castellazzi, A.M. Review on microbiota and effectiveness of probiotics use in older. World J. Clin. Cases 2015, 3, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimaraes, V.D.; Sokol, H.; Doré, J.; Corthier, J.; Furet, J.P. The Firmicutes/Bacteriodetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Mäkivuokko, H.; Tiihonen, K.; Tynkkynen, S.; Paulin, L.; Rautonen, N. The effect of age and non-steroidal anti-inflammatory drugs on human intestinal microbiota composition. Br. J. Nutr. 2010, 103, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, H.J.; Quagliarello, J.V. The microbiota and microbiome in aging: Potential implications in health and age-related diseases. J. Am. Geriatr. Soc. 2015, 63, 776–781. [Google Scholar] [CrossRef] [Green Version]
- Kostic, A.D.; Gevers, D.; Pedamallu Cs Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; Baselga, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, M.J.; Macfarlane, G.T. Changes in predominant bacterial populations in human faeces with age and with Clostridium difficile infection. J. Med. Microbiol. 2002, 51, 448–454. [Google Scholar] [CrossRef]
- Van Tongeren, S.P.; Slaets, J.P.J.; Harmsen, H.J.M.; Welling, G.W. Fecal microbiota composition and frailty. Appl. Environ. Microbiol. 2005, 71, 6438–6442. [Google Scholar] [CrossRef] [Green Version]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.B.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. Gut instincts: Microbiota as a key regulator of brain development, ageing, and neurodegeneration. J. Physiol. 2017, 595, 489–503. [Google Scholar] [CrossRef]
- Scheperjans, F.; Aho, V.; Pereira, P.A.B.; Koskinen, K.; Paulin, L.; Pekkonen, E.; Haapaniemi, E.; Kaakola, S.; Eerola-Rautio, J.; Pohja, M.; et al. Gut microbiota are related to Parkinson’s Disease and clinical phenotype. Mov. Disord. 2015, 30, 350–358. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haran, J.P.; Bahattarai, S.K.; Foley, S.E.; Dutta, P.; Ward, D.V.; Bucci, V.; McCormick, B.A. Alzheimer’s disease microbiome is associated with dysregulation of the anti-inflammatory p-glycoprotein pahtway. MBio 2019, 10, e00632-19. [Google Scholar] [CrossRef] [Green Version]
- Vernaya, M.; McAdam, J.; Hampton, M.D. Effectiveness of probiotics in reducing the incidence of Clostridium difficile-associated diarrhea in elderly patients: A systematic review. JBI Database Syst. Rev. Implement. Rep. 2017, 15, 140–164. [Google Scholar] [CrossRef] [PubMed]
- Krüger, J.F.; Hillesheim, E.; Pereira, A.C.S.N.; Camargo, C.Q.; Rabito, E.I. Probiotics for dementia: A systematic review and meta-analysis of randomized controlled trials. Nutr. Rev. 2021, 79, 160–170. [Google Scholar] [CrossRef]
- Martinez-Martinez, M.I.; Calabuig-Tolsá, R.; Cauli, O. The effect of probiotics as a treatment for constipation in elderly people: A systematic review. Arch. Gerontol. Geriatr. 2017, 71, 142–149. [Google Scholar] [CrossRef]
- Miller, L.E.; Lehtoranta, L.; Lehtinen, M.J. Short-term probiotic supplementation enhances cellular immune function in healthy elderly: Systematic review and meta-analysis of controlled studies. Nutr. Res. 2019, 64, 1–8. [Google Scholar] [CrossRef]
- Xie, C.; Li, J.; Wang, K.; Li, Q.; Chen, D. Probiotics for the prevention of antibiotic-associated diarrhoea in older patients: A systematic review. Travel Med. Infect. Dis. 2015, 13, 128–134. [Google Scholar] [CrossRef]
- Wachholz, P.A.; Nunes, V.D.S.; Polachini do Valle, A.; Jacinto, A.F.; Villas-Boas, P.J.F. Effectiveness of probiotics on the occurrence of infections in older people: A systematic review and meta-analysis. Age Ageing 2018, 47, 527–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillemard, E.; Tondu, F.; Lacoin, F.; Schrezenmeir, J. Consumption of a fermented dairy product containing the probiotic Lactobacillus casei DN-114 001 reduces the duration of respiratory infections in the elderly in a randomized controlled trial. Br. J. Nutr. 2010, 103, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Gohel, M.K.; Prajapati, J.B.; Mudgal, S.V.; Pandya, H.V.; Singh, U.S.; Trivedi, S.S.; Phatak, A.G.; Patel, R.M. Effect of probiotic dietary intervention on calcium and haematological parameters in geriatrics. J. Clin. Diagn. Res. 2016, 10, LC05–LC09. [Google Scholar] [CrossRef]
- Finamore, A.; Roselli, M.; Donini, L.; Brasili, E.; Rami, R.; Carnevali, P.; Mistura, L.; Pinto, A.; Giusti, A.; Mengheri, E. Supplementation with Bifidobacterium longum Bar33 and Lactobacillus helveticus Bar13 mixture improves immunity in elderly humans (over 75 years) and aged mice. Nutrition 2019, 63–64, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Bergillos-Meca, T.; Rasinkangas, P.; Korpela, K.; de Vos, W.M.; Gibson, G.R. Effects of soluble corn fiber alone or in synbiotic combination with Lactobacillus rhamnosus GG and the pilus-deficient derivative GG-PB12 on fecal microbiota, metabolism, and markers of immune function: A randomized, double-blind, placebo-controlled, crossover study in healthy elderly (Saimes study). Front. Immunol. 2017, 8, 1443. [Google Scholar]
- Bartosch, S.; Woodmansey, E.J.; Paterson, J.C.M.; McMurdo, M.E.T.; Macfarlane, G.T. Microbiological effects of consuming a synbiotic containing Bifidobacterium bifidum, Bifidobacterium lactis, and oligofructose in elderly persons, determined by real-time polymerase chain reaction and counting of viable bacteria. Clin. Infect. Dis. 2005, 40, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.; Prasad, J.; Gill, H.; Svenson, L.; Gopal, P. Impact of consumption of different levels of Bifidobacterium lactis HN019 on the intestinal microflora of elderly human subjects. J. Nutr. Health Aging 2007, 11, 26–31. [Google Scholar]
- Macfarlane, S.; Cleary, S.; Bahrami, B.; Reynolds, N.; Macfarlane, G.T. Synbiotic consumption changes the metabolism and composition of the gut microbiota in older people and modifies inflammatory processes: A randomized, double-blind, placebo-controlled crossover study. Aliment. Pharmacol. Ther. 2013, 38, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Kobayashi, Y.; Mori, N.; Sakagawa, M.; Xiao, J.; Moritani, T.; Sakane, N.; Nagai, N. Effect of combined bifidobacteria supplementation and resistance training on cognitive function, body composition and bowel habits of healthy elderly subjects. Benef. Microbes 2018, 9, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, K.; Gill, H.S.; Chandra, R.K. Enhancement of natural immune function by dietary consumption of Bifidobacterium lactis (HN019). Eur. J. Clin. Nutr. 2000, 54, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Moro-Garcia, M.A.; Alonso-Arias, R.; Baltadijeva, M.; Benitez, C.F.; Barrial, M.A.F.; Ruisánchez, E.D.; Santos, R.A.; Sánchez, M.A.; Miiján, J.S.; López-Larrea, C. Oral supplementation with Lactobacillus delbrueckii subsp. Bulgaricus 8481 enhances systemic immunity in elderly subjects. Age 2013, 35, 1311–1326. [Google Scholar] [CrossRef]
- Spaiser, S.J.; Culpepper, T.; Nieves, C.; Ukhanova, M.; Mai, V.; Percival, S.S.; Christman, M.C.; Langkamp-Henken, B. Lactobacillus gasseri KS-13, Bifidobacterium bifidum G9-1, and Bifidobacterium longum MM-2 ingestion induces a less inflammatory cytokine profile and a potentially beneficial shift in gut microbiota in older adults: A randomized, double-blind, placebo-controlled, crossover study. J. Am. Coll. Nutr. 2015, 34, 459–469. [Google Scholar] [PubMed]
- Shinkai, S.; Toba, M.; Saito, T.; Sato, I.; Tsubouchi, M.; Taira, K.; Kakumoto, K.; Inamatsu, T.; Yoshida, H.; Fujiwara, Y.; et al. Immunoprotective effects of oral intake of heat-killed Lactobacillus pentosus strain b240 in elderly adults: A randomized, double-blind, placebo-controlled trial. Br. J. Nutr. 2013, 109, 1856–1865. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.C.; Tiihonen, K.; Saarinen, M.; Putaala, H.; Rautonen, N. Influence of a combination of Lactobacillus acidophilus NCFM and lacitol on healthy elderly: Intestinal and immune parameters. Br. J. Nutr. 2009, 101, 367–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Östlund-Lgerström, L.; Kihlgren, A.; Repsilber, D.; Björkstén, B.; Brummer, R.J.; Schoultz, I. Probiotic administration among free-living older adults: A double blinded, randomized, placebo-controlled clinical trial. Nutr. J. 2016, 15, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyangale, E.P.; Farmer, S.; Cash, H.A.; Keller, D.; Chernoff, D.; Gibson, G.R. Bacillus coagulans GBI-30, 6086 modulates Faecalibacterium prausnitzii in older men and women. J. Nutr. 2015, 145, 1446–1452. [Google Scholar] [CrossRef] [Green Version]
- Manzoni, M.S.J.; Rossi, E.A.; Pauly-Silveira, N.D.; Pinto, R.A.; Roselino, M.N.; Carlos, I.Z.; Quilles, M.B.; Glória, M.B.A.; Cavallini, D.C.U. Consumption effect of a synbiotic beverage made form soy and yacon extracts containing Bifidobacterium animalis ssp. lactis BB-12 on the intestinal polyamine concentrations in elderly individuals. Food Res. Int. 2017, 99, 495–500. [Google Scholar] [CrossRef] [Green Version]
- Makino, S.; Ikegami, S.; Kume, A.; Horiuchi, H.; Sasaki, H.; Orii, N. Reducing the risk of infection in the elderly by dietary intake of yoghurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. Br. J. Nutr. 2010, 104, 998–1006. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Cha, L.; Sim, M.; Jung, S.; Chun, W.Y.; Baik, H.W.; Shin, D. Probiotic supplementation improves cognitive function and mood with changes in gut microbiota in community-dwelling older adults: A randomized, double-blind, placebo-controlled, multicenter trial. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 32–40. [Google Scholar] [CrossRef]
- Gao, B.; Chi, L.; Zhu, Y.; Shi, X.; Tu, P.; Li, B.; Yin, J.; Gao, N.; Shen, W.; Schnabi, B. An Introduction to Next Generation Sequencing Bioinformatic Analysis in Gut Microbiome Studies. Biomolecules 2021, 11, 530. [Google Scholar] [CrossRef]
- Prodan, A.; Tremaroli, V.; Brolin, H.; Zwinderman, A.H.; Nieuwdorp, M.; Levin, E. Comparing bioinformatics pipelines for microbial 16S rRNA amplicon sequencing. PLoS ONE 2020, 15, e0227434. [Google Scholar] [CrossRef] [Green Version]
- Graspeuntner, S.; Loeper, N.; Knuzel, S.; Baines, J.F.; Rupp, J. Selection of validated hypervariable regions is crucial in 16S-based microbiota studies of the female genital tract. Sci. Rep. 2018, 8, 9678. [Google Scholar] [CrossRef] [PubMed]
- Thorsen, J.; Brejnrod, A.; Mortensen, M.; Rasmussen, M.A.; Stokholm, J.; Al-Soud, W.A.; Sørensen, S.; Bisgaard, H.; Waage, J. Large-scale benchmarking reveals false discoveries and count transformation sensitivity in 16S rRNA gene amplicon data analysis methods used in microbiome studies. Microbiome 2016, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.; McDonald, D.; et al. Best practices or analyzing micrbiomes. Nat. Rev. Microbiol. 2018, 16, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y.; et al. Faecalibacterium prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm. Bowel Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef] [Green Version]
- Miquel, S.; Martin, R.; Rossi, O.; Bermudez-Humaran, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Shen, J.; Ran, Z.H. Association between Faecalibacterium prausnitzii Reduction and Inflammatory Bowel Disease: A Meta-Analysis and Systematic Review of the Literature. Gastroenterol. Res. Pract. 2014, 2014, 872725. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Hlder, C.V.; de Sousa Faria, A.V.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Gabherr, F.; Grander, C.; Effenberger, M.; Adolph, T.E.; Tilg, H. Gut dysfunction and non-alcoholic fatty liver disease. Front. Endocrinol. 2019, 10, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, M.A.; Jeffery, I.B.; Beaumont, M.; Bell, J.T.; Clark, A.G.; Ley, R.E.; O’Toole, P.W.; Spector, T.D.; Steves, C.J. Signatures of early frailty in the gut microbiota. Genome Med. 2016, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qui, X.; Zhang, M.; Yang, X.; Hong, N.; Yu, C. Faecalibacterium prausnitzii upregulates regulatory T cells and anti-inflammatory cytokines in treating TNBS-induced colitis. J. Chrohns Colitis 2013, 7, e558–e568. [Google Scholar]
- Matsuzaki, T.; Takagi, A.; Ikemura, H.; Matsuguchi, T.; Yokokura, T. Instestinal microflora: Probiotics and autoimmunity. J. Nutr. 2007, 137, 798S–802S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, S.; Watzl, B. Inulin and oligofructose: Review of experimental data on immune modulation. J. Nutr. 2007, 137, 2563S–2567S. [Google Scholar] [CrossRef]
- Butcher, S.K.; Chahal, H.; Nayak, L.; Sinclair, A.; Henriquez, N.V.; Sapey, E.; O’Mahony, D.; Lord, J.M. Senescene in innate immune responses: Reduced neutrophil phagocytic capacity and CD16 expression in elderly humans. J. Lukoc. Biol. 2001, 70, 881–886. [Google Scholar]
- Pawelec, G. Hallmarks of human “immunosenescen”: Adaptation or dysregulation? Immun. Ageing 2012, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Goronzy, J.J.; Lee, W.W.; Weyand, C.M. Aging and T-cell diversity. Exp. Gerontol. 2007, 42, 400–406. [Google Scholar] [CrossRef] [Green Version]
- Saurwein-Teissl, M.; Lung, T.L.; Marx, F.; Gschosser, C.; Asch, E.; Blasko, I.; Parson, W.; Böck, G.; Schönitzer, D.; Trannoy, E.; et al. Lack of antibody production following immunization in old age: Association with CD8(+)CD28(−) T cell clonal expansions and an imbalance in the production of Th1 and Th2 cytokines. J. Immunol. 2002, 168, 5893–5899. [Google Scholar] [CrossRef] [Green Version]
- Grimble, G.; Patil, D.; Silk, D. Assimilation of lactitol, an “unabsorbed” disaccharide in the normal human colon. Gut 1988, 29, 1666–1671. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.E.; Tennila, J.; Ouwehand, A.C. Efficacy and tolerance of lactitol supplementation for adult constipation: A systematic review and meta-analysis. Clin. Exp. Gastroenterol. 2014, 7, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Pinto-Sanchez, M.I.; Hall, G.B.; Ghajar, K.; Nardelli, A.; Bolino, C.; Lau, J.T.; Martin, F.; Cominetti, O.; Welsh, C.; Rieder, A.; et al. Probiotic Bifidobacterium longum NCC3001 reduces depression scores and alters brain activity: A pilot study in patients with irritable bowel syndrome. Gastroenterology 2017, 153, 448–459. [Google Scholar] [CrossRef]
- Foster, J.A.; Neufeld, K.A.M. Gut-brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Bagga, D.; Reichert, J.L.; Koschutnig, K.; Aigner, C.S.; Hozer, P.; Koskinen, K.; Moissl-Eichinger, C.; Schöpf, V. Probiotics drive gut microbiome triggering emotional brain signatures. Gut Microbes 2018, 9, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Tillisch, K.; Labus, J.; Kilpatrick, L.; Jiang, Z.; Stains, J.; Ebrat, B.; Guyonnet, D.; Legrain-Raspaud, S.; Trotin, B.; Naliboff, B.; et al. Consumption of fermented milk product with probiotic modulates brain activity. Gastroenterology 2013, 144, 1394–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerdá, B.; Pérez, M.; Pérez-Santiago, J.; Tornero-Aguilera, J.F.; Gonzálex-Soltero, R.; Larrosa, M. Gut microbiota modification: Another piece in the puzzle of the benefits of physical exercise in health? Front. Physiol. 2016, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- De Sire, A.; de Sire, R.; Petito, V.; Masi, L.; Cisari, C.; Gasbarrini, A.; Scaldaferri, F.; Invernizzi, M. Gut-joint axis: The role of physical exercise on gut microbiota modulation in older people with osteoarthritis. Nutrients 2020, 12, 574. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yu, D.; Xue, L.; Li, H.; Du, J. Probiotics modulate the microbiota-gut-brain axis and improve memory deficits in aged SAMP8 mice. Acta Pharm. Sin. B 2020, 10, 475–487. [Google Scholar] [CrossRef]
- Ni, Y.; Yang, X.; Zheng, L.; Wang, Z.; Wu, L.; Jiang, J.; Yang, T.; Ma, L.; Fu, Z. Lactobacillus and Bifidobacterium imporves physiological function and cognitive ability in aged mice by the regulation of gut microbiota. Mol. Nutr. Food Res. 2019, 63, e1900603. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Shin, M.; Mun, D.; Jeong, S.Y.; Jeong, D.Y. Probiotic Lactobacillus fermentum strain JDFM216 improves cognitive behavior and modulates immune response with gut microbiota. Sci. Rep. 2020, 10, 21701. [Google Scholar] [CrossRef]
- Lloyd-Jones, D.; Adams, R.J.; Brown, T.M.; Carnethon, M.; Dai, S.; Simone, G.D.; Ferguson, T.B.; Ford, E.; Furie, K.; Gillespie, C.; et al. Heart disease and stroke statistics—2010 update: A report from the American Heart Association. Circulation 2010, 121, e46–e215. [Google Scholar] [PubMed]
- Sabatine, M.S.; Wiviott, S.D.; Im, K.; Murphy, S.A.; Giuglano, R.P. Efficacy and safety of further lowering of low-density lipoprotein cholesterol in patients with very low levels: A meta-analysis. JAMA Cardiol 2018, 3, 823–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.Y.; Jiao, R.; Ma, K.Y. Cholesterol-lowering nutraceuticals and functional foods. J. Agric. Food Chem 2008, 56, 8761–8773. [Google Scholar] [CrossRef]
- Shimizu, M.; Hashiquchi, M.; Shiga, T.; Tamura, H.O.; Mochizuki, M. Meta analysis: Effects of probiotic supplementation on lipid profiles in normal to mildly hypercholesterolemic individuals. PLoS ONE 2015, 10, e0139795. [Google Scholar] [CrossRef] [PubMed]
- Lupsa, B.C.; Insogna, K. Bone health and osteoporosis. Endocrinol. Metab. Clin. N. Am. 2015, 44, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.C.; Biver, E.; Kaufman, J.; Bauer, J.; Branco, J.; Brandi, M.L.; Bruyère, O.; Coxam, V.; Cruz-Jentoft, A.; Czerwinski, E.; et al. The role of calcium supplementation in healthy musculoskeletal ageing: An expert consensus meeting of the European Society for Clinical and Economic Aspects of Osteoporosis, Osteoarthritis and Musculoskeletal Diseases (ESCEO) and the International Foundation for Osteoporosis (IOF). Osteoporos Int. 2017, 28, 447–462. [Google Scholar]
- Rizzoli, R.; Biver, E. Are probiotics the new calcium and vitamin D for bone health? Curr. Osteopros. Rep. 2020, 18, 273–284. [Google Scholar] [CrossRef]
- Valentini, L.; Pinto, A.; Bourdel-Marchasson, I.; Ostan, R.; Brigidi, P.; Turroni, S.; Hrelia, S.; Hrelia, P.; Bereswill, S.; Fischer, A.; et al. Impact of personalized diet and probiotic supplementation on inflammation, nutritional parameters and instestinal microbiota- The “RISTOMED project”: Randomized controlled trial in healthy older people. Clin. Nutr. 2015, 34, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Ford, A.L.; Nagulesapillai, V.; Piano, A.; Auger, J.; Girard, S.; Christman, M.; Tompkins, T.A.; Dahl, W.J. Microbiota stability and gastrointestinal tolerance in response to a high-protein diet with and without a prebiotic, probiotic, and synbiotic: A randomized, double-blind, placebo-controlled trial in older women. J. Acad. Nutr. Diet 2020, 120, 500.e10–516.e10. [Google Scholar] [CrossRef] [Green Version]
- Bourdel-Marchasson, I.; Ostan, R.; Regueme, S.C.; Pinto, A.; Pryen, F.; Charrouf, Z.; d’Alessio, P.; Baudron, C.R.; Guerville, F.; Durrieu, J.; et al. Quality of life: Psychological symptoms-effects of a 2-month healthy diet and nutraceutical intervention; a randomized, open-lbael intervention trial (RISTOMED). Nutrients 2020, 12, 800. [Google Scholar] [CrossRef] [Green Version]
- Meance, S.; Cayuela, C.; Turchet, P.; Raimondi, A.; Lucas, C.; Antoine, J. A fermented milk with a Bifidobacterium probiotic strain DN-173 010 shortened oro-fecal gut transit time in elderly. Microbiol. Ecol. Health Dis. 2001, 13, 217–222. [Google Scholar] [CrossRef]
- Matsumoto, M.; Ohishi, H.; Benno, Y. Impact of LKM512 yogurt on improvement of intestinal environment of the elderly. FEMS Immunol. Med. Microbiol. 2001, 31, 181–186. [Google Scholar] [CrossRef]
- Butler, C.C.; Lau, M.; Gillespie, D.; Owen-Jones, E.; Lown, M.; Wootton, M.; Calder, P.C.; Bayer, A.J.; Moore, M.; Little, P.; et al. Effect of probiotic use on antibiotic administration among care home residents: A randomized clinical trial. JAMA 2020, 324, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Castro-Herrera, V.M.; Fisk, H.L.; Wootton, M.; Lown, M.; Owen-Jones, E.; Lau, M.; Lowe, R.; Hood, K.; Gillespie, D.; Hobbs, F.D.R.; et al. Combination of the probiotics Lacticaseibacillus rhamnosus GG and Bifidobacterium animalis subsp. Lactis, BB-12 has limited effect on biomarkers of immunity and inflammation in older people resident in care homes: Results from the probiotics to reduce infections iN CarE home reSidentS randomized, controlled trial. Front. Immunol. 2021, 12, 643321. [Google Scholar] [PubMed]
- Yamamoto, Y.; Saruta, J.; Takahashi, T.; To, M.; Shimizu, T.; Hayashi, T.; Morozumi, T.; Kubota, N.; Kamata, Y.; Makino, S.; et al. Effect of ingesting yogurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1 on influenza virus-bound salivary IgA in elderly residents of nursing homes: A randomized controlled trial. Acta Odontol. Scand. 2019, 77, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Owen-Jones, E.; Lowe, R.; Lown, M.; Gillespie, D.; Addison, K.; Bayer, T.; Calder, P.C.; Davies, J.; Davoudianfar, M.; Downs, J.; et al. Protocol for a double-blind placebo-controlled trial to evaluate the efficacy of probiotics in reducing antibiotics for infection in care home residents: The Probiotics to Reduce Infections iN CarE home reSidentS (PRINCESS) trial. BMJ Open 2019, 9, e027513. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Study and Country | Design and Sample Size * | Sex (Female/Male) | Age # | Probiotic/Synbiotic and Dosage in CFU/day | Duration of Therapy | Outcomes Studied | Reported Treatment Effects § |
|---|---|---|---|---|---|---|---|
| Ahmed 2007 New Zealand | Parallel n = 80 | NR | 60–87 | B. lactis HN019 5 × 109 1 × 109 or 6.5 × 107 | 4 w | Changes in fecal microflora | Bifidobacteria ↑ Lactobacilli ↑ Streptococci ↑ Coliforms ↑ |
| Arunachalam 2000 New Zealand | Parallel n = 25 | 18/7 | 60–83 | B. lactis HN019 1.5 × 1011 (twice daily) | 6 w | Immunomodulation (and presence of B. lactis in stool) | IFN-a ↑ phagocytic capacity ↑ bactericidal activity ↑ (both groups) |
| Bartosch 2004 UK | Parallel n = 18 | 18/0 | 63–90 | B. bifidum BB-02 B. lactis BL-01 ORAFTI’s Raftilose® Synergy 1 (chicory inulin and oligofructose) ∼3.5 × 1010 (of each bacterium) | 4 w | Changes in fecal Bifidobacteria and Lactobacillus | TotalBifidobacteria ↑ B. angulatum ↑ (B. bifidum ↑) (B. lactis ↑) Total lactobacilli ↑ |
| Costabile 2017 UK | Crossover n = 40 | NR | 60–80 | L. rhamnosus GG pilus-deficient L. rhamnosus GG-PB12 Promitor™ (corn fiber) 12 × 1010 | 3 w (for each supplement) | Changes in fecal microflora, immunity, and blood lipids | Parabacteroides ↑ Ruminococcaceae ↑ Oscillospira ↓ Desulfovibrio ↓ (L. rhamnosus GG ↑) Total cholesterol ↓ LDL-cholesterol ↓ |
| Finamore 2019 Italy | Parallel n = 98 | 29/69 | 84.6 (mean) | B. longum Bar33 L. helveticus Bar13 1 × 109 | 30 d | Improvements in innate and adaptive immunity, anthropometrics and wellbeing | Regulatory T cells ↑ B cells ↑ natural killer activity ↑ CD4+ naive T cells ↑ CD8+ naive T cells ↑ CD8+ activated memory cells ↑ CD4+ effector memory cells ↓ |
| Gohel 2016 India | Crossover n = 76 | 38/38 | 64–74 | L. helveticus MTCC 5463 S. thermophilus MTCC 5460 (in honey supplemented fermented milk) > 108 CFU/mL L. helveticus (dose: 200 mL/day) S. thermo. NR | 4 w | Effect on serum calcium and hematological parameters | Serum calcium level ↑ |
| Guillemard 2010 France | Parallel (multi-center) n = 1072 | 672 /400 | 69–95 | L. casei DN-114 001 S. thermophilus L. delbrueckii subsp. bulgaricus L. casei >1010 CFU/100 g The other two at >109 CFU/100 g (dose: 200 g/day) | 3 m | Resistance to common infectious disease (and prescence of L. paracasei in stool) | Duration of CID ↓ |
| Inoue 2018 Japan | Parallell n = 38 | 24/14 | 66–78 | B. longum BB536, B. breve M-16V B. infantis M-63 B. breve B-3 (dextrin) and moderate resistance training ∼1.25 × 1010 (of each bacterium) | 12 w | Cognitive function | MOCA-J ↑ (both groups) Defecation frequency↑ Mental state ↓ body mass/BMI ↓ Anxiety ↓ (placebo) |
| Kim 2021 Korea | Parallel (multi-center) n = 53 | NR | Treatm: 72 Placebo: 71.1 (mean) | B. bifidum BGN4 B. longum BORI 1 × 109 | 12 w | Intestinal and brain health | inflammation-causing gut bacteria ↓ Mental flexibility ↑ BDNF ↑ (placebo) |
| Macfarlane 2013 UK | Crossover n = 43 | 22/21 | 65–90 | B. longum ORAFTI’s Raftilose® Synergy 1 (chicory inulin and oligofructose) Ca 2 × 1011 | 4 w | Changes in fecal Bifidobacteria counts, changes in fecal microflora, inflammatory markers, bowel habit and health status | Bifidobacteria ↑ B. adolescentis ↑ B. angulatum ↑ B. bifidum ↑ (B. longum ↑) Actinobacteria ↑ Firmicutes ↑ Proteobacteria ↓ Butyrate ↑ Isobutyrate ↑ Acetate ↑ TNF-a ↓ |
| Manzoni 2017 Brazil | Parallel n = 29 | NR | Treatm: 67 Placebo: 71 (mean) | B.animalis ssp. lactis BB-12 Yacon (prebiotic source) and soy extracts 1010 CFU/100 mL (dose: 150 mL/day) | 4 w | Changes in counts of fecal: Bifidobacterium spp., Clostridium spp., Enterobacteria. Additionally, polyamines and inflammatory cytokines | Polyamine levels ↑ (both groups) |
| Moro-Gracía 2013 Spain | Parallel (multi-center) n = 47 | 7/40 | Treatm: 65–82 Placebo: 65–90 | L. delbrueckii subsp. bulgaricus 8481 3 × 107/capsule (dose: 3 capsules/day) | 6 mo | Immune cell populations, cytokines, T cell receptor excision circle (TREC), human β-defensin-2 (hBD-2) concentrations, cytomegalovirus (CMV) IgG titers | NK cells ↑ CD8+ T cell ↓ CD4/CD8 ratio ↑ Senescent T cells ↓ NAÏVE CD4+ T cells ↑ Memory T cells ↓ TREC ↑ IL-8 ↓ CMV titers ↑ (placebo) |
| Nyangale 2015 UK | Crossover n = 36 | 25/17 | 65–80 | Bac. coagulans GBI-30, 6086 (BC30) 1 × 109 | 28 d | Immunomodulation, changes in fecal microflora, Calprotectin and SCFA. Additionally, digestive health and mood diaries. | F. prausnitzii ↑ IL-10 ↑ TNF-a ↑ (placebo) SCFA ↑ (both groups) |
| Ouwehand 2009 Finland | Parallel n = 47 | 35/12 | Treatm: 70.3 Placebo: 71.7 (mean) | L. acidophillus NCFM Lactitol (prebiotic) 2 × 109 CFU/g (dose: 5–5.5 g, twice a day) | 2 w | Changes in fecal: Bifidobacteria L acidophilus L acidophilus NCFM C. perfringes Sulphate reducers (i.e., D. intestinalis) | Stool frequency ↑ Bifidobacteria ↑ Spermidine ↑ Fecal PGE2 ↑ Fecal IgA ↑ (L. acidophilus NCFM ↑) |
| Shinkai 2013 Japan | Parallel n = 278 | 140/138 | >65 yrs | L. pentosus strain b240 2 × 109 or 2 × 1010 | 20 w | Common cold, QoL | Common cold incidence ↓ General health perception ↑ |
| Spaiser 2015 USA | Crossover n = 32 | 22/10 | 69.8 (mean) | L. gasseri KS-13 B. bifidum G9-1 B. longum MM2 1.5 × 109/capsule (Dose: 2 capsules/day) | 3 w | Changes in fecal: Bifidobacteria Lactic acid bacteria E. coli Circulating CD4+ lymphocytes and PHA stimulated cytokine release Digestive health | Bifidobacteria ↑ Lactic acid bacteria ↑ E. coli ↓ IL-10 ↑ IL-5 ↑ CD4+ ↓ (placebo) |
| Östlund-Lagerström 2016 Sweden | Parallel n = 249 | 152/97 | Treatm: 72.6 Placebo: 72 (mean) | L. reuteri DSM 17938 Rhamnose, galactooligosaccharide and maltodextrin 108 CFU/day | 12 w | Digestive health Wellbeing | No significant effects |
| Author, Year | Probiotic/Synbiotic | Total study Duration | Sampling Timepoints | Number of Subjects’ Feces Analyzed | Microbiota Assessed | Methods |
|---|---|---|---|---|---|---|
| Synbiotic interventions | ||||||
| Bartosch [48] | B. bifidum BB-02 B. lactis BL-01 ORAFTI’s Raftilose® Synergy 1 (chicory inulin and oligofructose) | prefeeding (1 week) feeding (4 weeks) postfeeding (week 8) | 1 w, 4 w, 8 w | 18 n = 9 (placebo) n = 9 (synbiotic) | Agar plate cultures | |
| Total anaerobes | Wilkins-Chalgren agar | |||||
| Bifidobacteria | Beerens medium | |||||
| Lactobacilli | Rogosa | |||||
| B. lactis, B. bifidum, Bifidobacteria genus | qPCR (DNA primers) | |||||
| Costabile [47] | L. rhamnosus GG pilus-deficient L. rhamnosus GG-PB12 Promitor™ (corn fiber) | 147 d (2 w run-in, 3 w intervention, 3 weeks washout) | 0, 21, 63, 105, 147 d | 111 LLG-PB12 + SCF (n = 37) LGG + SCF (n = 37) SCF (n = 37) | Quantities of the L. rhamnosus GG strains and total bacteria | qPCR (16s rRNA) |
| Phylogenetic analysis | 16s rRNA Illumina Miseg sequencing and profiling (V3,V4 variable region sequencing using primers) | |||||
| Macfarlane [50] | B. longum ORAFTI’s Raftilose® Synergy 1 (chicory inulin and oligofructose) | 12 w (4 w intervention) | baseline, 2 w (mid-intervention), 4 w (end) | 43 (crossover) | Firmicutes (Clostridium cluster XIVa, F. prausnitzii group, Ruminococci, Roseburia intestinalis, lactic acid bacteria) | FISH (16s rRNA) |
| Bacteroidetes (Bacteroides/Prevotella), Actinobacteria (Atopobium group, Bifidobacteria) | ||||||
| Proteobacteria (Enterobacteriaceae, Desulphovibrio) | ||||||
| Bifidobacteria | ||||||
| Total bacteria (Eubacterial probe) | ||||||
| Manzoni [59] | B. animalis ssp. lactis BB-12 Yacon (prebiotic source) and soy extracts | 8 w (2 w prefeeding, 4 w feeding, 2 w postfeeding) | 1 w, 6 w (end of feeding), 8 w (end of washout) | 12 n = 6 (intervention) n = 6 (placebo) | Agar plate cultures | |
| Clostridium | Reinforced Clostridial Agar | |||||
| Enterobacteriaceae | MacConkey medium | |||||
| Bifidobacteria | Iodoacetate Medium-25 (BIM25) | |||||
| Ouwehand [56] | L acidophillus NCFM Lactitol (prebiotic) | 6 w (2 w run in, 2 w intervention, 2 w washout) | baseline, 2 weeks, 4 weeks | 47 n = 24 (intervention) n = 23 (placebo) | Bifidobacteria L. acidophilus L. acidophilus NCFM C. perfringes Sulphate reducers (D. intestinalis) | qPCR (16s rRNA probes) |
| Total bacteria counts | Flow cytometry | |||||
| Probiotic interventions | ||||||
| Ahmed [49] | B. lactis HN019 (in skim milk) | 8 w (2 w run in, 4 w intervention, 2 w washout) | 0,2,4,6,7,8 w | 66 Bifidobacterium: low (n = 18), med (n = 15), high (n = 19), placebo (n = 14) | Agar plate cultures | |
| Bifidobacteria | Beerens medium | |||||
| Lactobacilli | Rogosa SL Agar | |||||
| Streptococci Enterobacteria | MacConkey agar and bile esulin azide agar | |||||
| Total anaerobes | Brucella agar | |||||
| Bacteroidetes | Bacteroidetes-bile-esculin | |||||
| Yeast and mold | Sabaroud dextrose agar | |||||
| Arunachalam [52] | B. lactis HN019 (in skim milk) | 6 w | NR | 25 n = 13 (intervention) n = 12 (placebo) | B. lactis HN019 | RAPD-DNA (strain-specific DNA probe) |
| Bacterial cell viability in test product | MRSC Agar plate cultures, Bifidobacteria- Beerens’ medium | |||||
| Guillemard [44] | L. casei DN-114 001 S. thermophilus L. delbrueckii subsp. bulgaricus (in fermented dairy product, Actimel) | 3 m (84 d) | 1,2,3,4 m | 63 n = 32 (intervention) n = 31 (placebo) | L. paracasei | qPCR (L. paracasei group specific) |
| Kim [61] | B. bifidum BGN4 B. longum BORI | 12 w | weekly | Not clear | Phylogenetic analysis | 16s rRNA Illumina Miseg sequencing and profiling (V3,V4 variable region sequencing using primers) |
| Nyangale [58] | Bac. coagulans GBI-30, 6086 (BC30) | 28 d | baseline, 28d | 36 (crossover) | FISH (DNA olig probes): | |
| Bifidobacterium spp. | Bif164 | |||||
| Lactobacillus spp. and Enterococcus spp. | LAB158 | |||||
| Clostridium coccoides, Eubacterium rectale group, (Clostridium cluster XIVa and XIVb) | EREC482 | |||||
| Clostridium lituseburense group (Clostridium cluster XI) | CLIT135 | |||||
| Bacteroidaceae spp., Prevotellaceae spp., some of Porphyromonadaceae spp. | BAC303 | |||||
| F. prausnitzii and relatives | Fprau645 | |||||
| Bacillus spp., including B. coagulans | Bcoa191 | |||||
| Spaiser [54] | L. gasseri KS-13 B. bifidum G9-1 B. longum MM2 | 3 w | baseline, 3 w | 28 (crossover) | Bifidobacteria | qPCR (DNA primers) |
| Lactic acid bacteria | ||||||
| E. coli | ||||||
| OTU Classification | pyrosequencing (ESPIRIT tree algorithm for binning for of sequences, similarity levels of 98% and 95%) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hutchinson, A.N.; Bergh, C.; Kruger, K.; Sűsserová, M.; Allen, J.; Améen, S.; Tingö, L. The Effect of Probiotics on Health Outcomes in the Elderly: A Systematic Review of Randomized, Placebo-Controlled Studies. Microorganisms 2021, 9, 1344. https://doi.org/10.3390/microorganisms9061344
Hutchinson AN, Bergh C, Kruger K, Sűsserová M, Allen J, Améen S, Tingö L. The Effect of Probiotics on Health Outcomes in the Elderly: A Systematic Review of Randomized, Placebo-Controlled Studies. Microorganisms. 2021; 9(6):1344. https://doi.org/10.3390/microorganisms9061344
Chicago/Turabian StyleHutchinson, Ashley N., Cecilia Bergh, Kirsten Kruger, Martina Sűsserová, Jessica Allen, Sophie Améen, and Lina Tingö. 2021. "The Effect of Probiotics on Health Outcomes in the Elderly: A Systematic Review of Randomized, Placebo-Controlled Studies" Microorganisms 9, no. 6: 1344. https://doi.org/10.3390/microorganisms9061344
APA StyleHutchinson, A. N., Bergh, C., Kruger, K., Sűsserová, M., Allen, J., Améen, S., & Tingö, L. (2021). The Effect of Probiotics on Health Outcomes in the Elderly: A Systematic Review of Randomized, Placebo-Controlled Studies. Microorganisms, 9(6), 1344. https://doi.org/10.3390/microorganisms9061344