
Molecular Subtyping of Blastocystis sp. Isolated from Farmed Animals in Southern Italy

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction and Sequencing of Blastocystis Isolates
2.3. Phylogenetic and Genetic Diversity Analyses
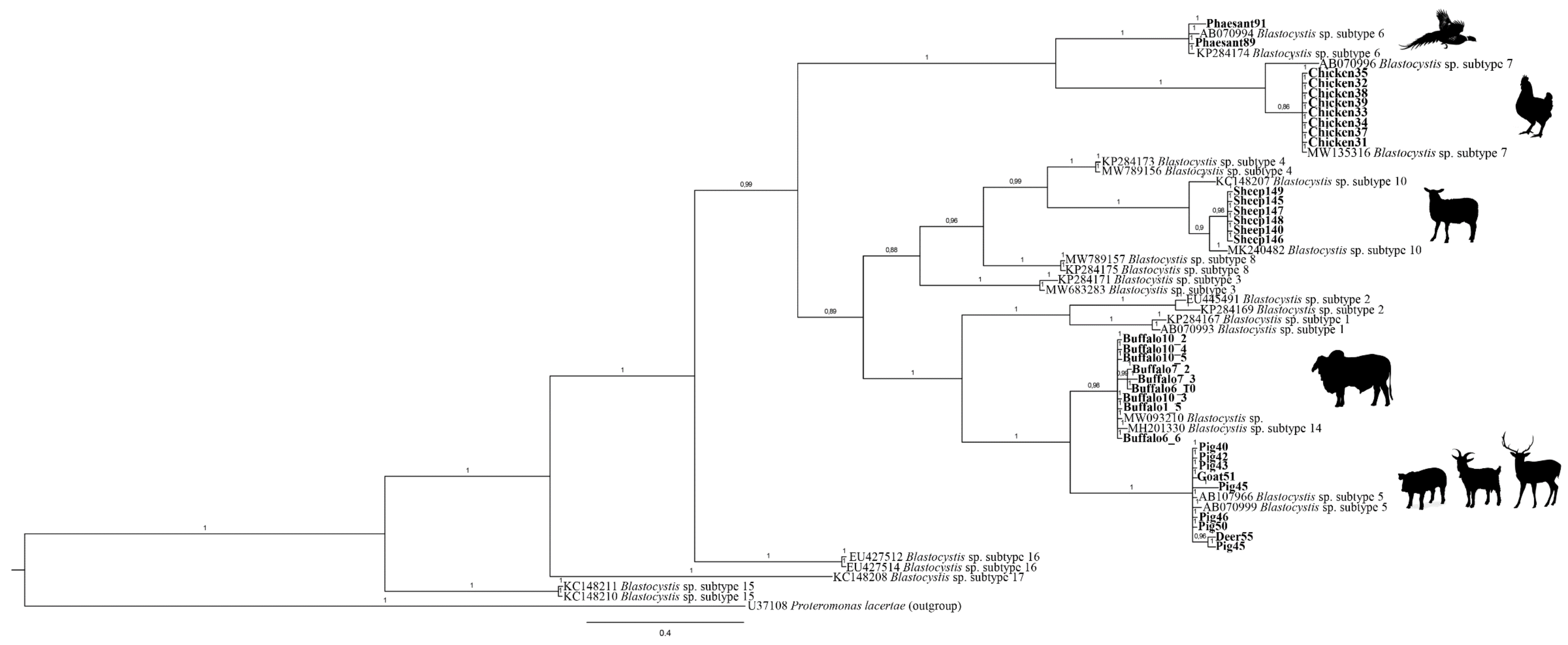
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Adl, S.M.; Bass, D.; Lane, C.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.; Burki, F.; et al. Revisions to the classification, nomenclature, and diversity of eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [Green Version]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.U.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Gabrielli, S.; Stensvold, C.R.; Mattiucci, S. Blastocystis. Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Suresh, K.; Ng, G.C.; Ramachandran, N.P.; Ho, L.C.; Yap, E.H.; Singh, M. In vitro encystment and experimental infections of Blastocystis hominis. Parasitol. Res. 1993, 79, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Yoshida, K.; Nakajima, A.; Yamanari, K.; Iwatani, S.; Kimata, I. Fecal-oral transmission of the cyst form of Blastocystis hominis in rats. Parasitol. Res. 2004, 94, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.S.W. New Insights on classification, identification, and clinical relevance of Blastocystis spp. Clin. Microbiol. Rev. 2008, 21, 639–665. [Google Scholar] [CrossRef] [Green Version]
- Stensvold, C.R.; Clark, C.G. Pre-empting pandora’s box: Blastocystis subtypes revisited. Trends Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Maloney, J.; Santin, M. Mind the gap: New full-length sequences of Blastocystis subtypes generated via oxford nanopore minion sequencing allow for comparisons between full-length and partial sequences of the small subunit of the ribosomal RNA gene. Microorganisms 2021, 9, 997. [Google Scholar] [CrossRef]
- Alfellani, M.A.; Taner-Mulla, D.; Jacob, A.; Imeede, C.A.; Yoshikawa, H.; Stensvold, C.R.; Clark, C.G. Genetic diversity of Blastocystis in livestock and zoo animals. Protist 2013, 164, 497–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cian, A.; El Safadi, D.; Osman, M.; Moriniere, R.; Gantois, N.; Benamrouz-Vanneste, S.; Delgado-Viscogliosi, P.; Guyot, K.; Li, L.-L.; Monchy, S.; et al. Molecular epidemiology of Blastocystis sp. in various animal groups from two French zoos and evaluation of potential zoonotic risk. PLoS ONE 2017, 12, e0169659. [Google Scholar] [CrossRef] [Green Version]
- Wawrzyniak, I.; Poirier, P.; Viscogliosi, E.; Dionigia, M.; Texier, C.; Delbac, F.; El Alaoui, H. Blastocystis, an unrecognized parasite: An overview of pathogenesis and diagnosis. Ther. Adv. Infect. Dis. 2013, 1, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Meloni, D.; Sanciu, G.; Poirier, P.; El Alaoui, H.; Chabé, M.; Delhaes, L.; Dei-Cas, E.; Delbac, F.; Fiori, P.L.; Di Cave, D.; et al. Molecular subtyping of Blastocystis sp. isolates from symptomatic patients in Italy. Parasitol. Res. 2011, 109, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Crisafi, B.; Gabrielli, S.; Paoletti, M.; Cancrini, G. Molecular epidemiology and genetic diversity of Blastocystis infection in humans in Italy. Epidemiol. Infect. 2015, 144, 635–646. [Google Scholar] [CrossRef]
- Sulekova, L.F.; Gabrielli, S.; Furzi, F.; Milardi, G.L.; Biliotti, E.; De Angelis, M.; Iaiani, G.; Fimiani, C.; Maiorano, M.; Mattiucci, S.; et al. Molecular characterization of Blastocystis subtypes in HIV-positive patients and evaluation of risk factors for colonization. BMC Infect. Dis. 2019, 19, 876–877. [Google Scholar] [CrossRef] [Green Version]
- Gabrielli, S.; Furzi, F.; Sulekova, L.F.; Taliani, G.; Mattiucci, S. Occurrence of Blastocystis-subtypes in patients from Italy revealed association of ST3 with a healthy gut microbiota. Parasite Epidemiol. Control 2020, 9, e00134. [Google Scholar] [CrossRef] [PubMed]
- Piubelli, C.; Soleymanpoor, H.; Giorli, G.; Formenti, F.; Buonfrate, D.; Bisoffi, Z.; Perandin, F. Blastocystis prevalence and subtypes in autochthonous and immigrant patients in a referral centre for parasitic infections in Italy. PLoS ONE 2019, 14, e0210171. [Google Scholar] [CrossRef]
- Caradonna, T.; Marangi, M.; Del Chierico, F.; Ferrari, N.; Reddel, S.; Bracaglia, G.; Normanno, G.; Putignani, L.; Giangaspero, A. Detection and prevalence of protozoan parasites in ready-to-eat packaged salads on sale in Italy. Food Microbiol. 2017, 67, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Angelici, M.C.; Nardis, C.; Scarpelli, R.; Ade, P. Blastocystis hominis transmission by non-potable water: A case report in Italy. N. Microbiol. 2018, 41, 173–177. [Google Scholar]
- Masucci, L.; Graffeo, R.; Bani, S.; Bugli, F.; Boccia, S.; Nicolotti, N.; Fiori, B.; Fadda, G.; Spanu, T. Intestinal parasites isolated in a large teaching hospital, Italy, 1 May 2006 to 31 December 2008. Eurosurveillance 2011, 16, 19891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capasso, M.; Maurelli, M.P.; Ianniello, D.; Alves, L.C.; Amadesi, A.; Laricchiuta, P.; Silvestre, P.; Campolo, M.; Cringoli, G.; Rinaldi, L. Use of Mini-FLOTAC and Fill-FLOTAC for rapidly diagnosing parasitic infections in zoo mammals. Rev. Bras. Parasitol. Veterinária 2019, 28, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Gazzonis, A.L.; Marangi, M.; Zanzani, S.A.; Villa, L.; Giangaspero, A.; Manfredi, M.T. Molecular epidemiology of blastocystis sp. in dogs housed in italian rescue shelters. Parasitol. Res. 2019, 118, 3011–3017. [Google Scholar] [CrossRef]
- Zanzani, S.A.; Gazzonis, A.L.; Epis, S.; Manfredi, M.T. Study of the gastrointestinal parasitic fauna of captive non-human primates (Macaca fascicularis). Parasitol. Res. 2016, 115, 307–312. [Google Scholar] [CrossRef]
- Russini, V.; Di Filippo, M.M.; Fanelli, R.; Polidori, M.; Berrilli, F.; Di Cave, D.; Novelletto, A.; Calderini, P. Characterization of prevalence and genetic subtypes of blastocystis sp. in wild and domestic Suidae of central Italy aided by amplicon NGS. Veter- Parasitol. Reg. Stud. Rep. 2020, 22, 100472. [Google Scholar] [CrossRef]
- Santín, M.; Gomez_Munoz, M.T.; Solano-Aguilar, G.; Fayer, R. Development of a new PCR protocol to detect and subtype blastocystis spp. from humans and animals. Parasitol. Res. 2011, 109, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Scicluna, S.M.; Tawari, B.; Clark, C.G. DNA barcoding of blastocystis. Protist 2006, 157, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Suresh, G.K.; Tan, K.S.; Thompson, R.A.; Traub, R.; Viscogliosi, E.; Yoshikawa, H.; Clark, C.G. Terminology for blastocystis subtypes—A consensus. Trends Parasitol. 2007, 23, 93–96. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Wylezich, C.; Belka, A.; Hanke, D.; Beer, M.; Blome, S.; Hoeper, D. Metagenomics for broad and improved parasite detection: A proof-of-concept study using swine faecal samples. Int. J. Parasitol. 2019, 49, 769–777. [Google Scholar] [CrossRef]
- Eroglu, F.; Koltas, I.S. Evaluation of the transmission mode of B. hominis by using PCR method. Parasitol. Res. 2010, 107, 841–845. [Google Scholar] [CrossRef]
- Richard, R.L.; Ithoi, I.; Majid, M.A.A.; Sulaiman, W.Y.W.; Tan, T.C.; Nissapatorn, V.; Lim, Y.A.L. Monitoring of Waterborne Parasites in Two Drinking Water Treatment Plants: A Study in Sarawak, Malaysia. Int. J. Environ. Res. Public Health 2016, 13, 641. [Google Scholar] [CrossRef] [Green Version]
- Stensvold, C.R.; Lewis, H.C.; Hammerum, A.M.; Porsbo, L.J.; Nielsen, S.S.; Olsen, K.E.P.; Arendrup, M.C.; Nielsen, H.V.; Mølbak, K. Blastocystis: Unravelling potential risk factors and clinical significance of a common but neglected parasite. Epidemiol. Infect. 2009, 137, 1655–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Mirza, H.; Tan, K.S.W. Intra-subtype variation in enteroadhesion accounts for differences in epithelial barrier disruption and is associated with metronidazole resistance in blastocystis subtype-7. PLoS Negl. Trop. Dis. 2014, 8, e2885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Márquez, M.V.; Guna, R.; Munoz, C.; Gómez-Muñoz, M.T.; Borras, R. High prevalence of subtype 4 among isolates of blastocystis hominis from symptomatic patients of a health district of Valencia (Spain). Parasitol. Res. 2009, 105, 949–955. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Alfellani, M.; Clark, C.G. Levels of genetic diversity vary dramatically between blastocystis subtypes. Infect. Genet. Evol. 2012, 12, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Nieves-Ramírez, M.E.; Partida-Rodríguez, O.; Laforest-Lapointe, I.; Reynolds, L.A.; Brown, E.M.; Valdez-Salazar, A.; Morán-Silva, P.; Rojas-Velázquez, L.; Morien, E.; Parfrey, L.W.; et al. Asymptomatic intestinal colonization with protist blastocystis is strongly associated with distinct microbiome ecological patterns. mSystems 2018, 3, e00007-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Host | Scientific Name | Collected Samples (N) | Positive Samples (%) | STs Identified | Alleles |
|---|---|---|---|---|---|
| Cow | Bos taurus | 13 | 1 (7.69) | untypable | - |
| Donkey | Equus asinus | 2 | 0 | - | - |
| Duck | Anas platyrhynchos domesticus | 2 | 0 | - | - |
| Fallow deer | Dama dama | 1 | 1 (100) | 5 | 17 |
| Goat | Capra aegagrus hircus | 9 | 4 (44.4) | 5 | - |
| Chicken | Gallus gallus domesticus | 17 | 8 (47) | 7 | 41 |
| Horse | Equus caballus | 3 | 0 | - | - |
| Ostrich | Struthio camelus | 2 | 0 | - | - |
| Peacock | Pavo sp. | 7 | 3 (42.85) | untypable | - |
| Pheasant | Phasianus colchicus | 11 | 6 (54.5) | 6 | 122 |
| Pig | Sus scrofa domesticus | 13 | 10 (76.92) | 5 | 17, 115, 153 |
| Sheep | Ovis aries | 11 | 9 (81.8) | 10 | 152 |
| Turkey | Meleagris gallopavo | 1 | 0 | - | - |
| Water buffalo | Bubalus bubalis | 101 | 18 (17.82) | 14 | 157 |
| Total | 193 | 60 (31.08%) |
| Sample ID/Isolate | Blastocystis Subtype (GenBank Accession Number) | Host | Locality | References | Similarity (%) |
|---|---|---|---|---|---|
| Phaesant91 | ST6 (MW713074) | Tibetan goat | China | Chang et al., unpublished | 99.63 |
| Phaesant89 | ST6 (MW713074) | Tibetan goat | China | Chang et al., unpublished | 99 |
| Chicken31 | ST7 (KY488585) | - | Southwest of Iran | Salehi et al., unpublished | 100 |
| Chicken32 | ST7 (KY488585) | - | Southwest of Iran | Salehi et al., unpublished | 100 |
| Chicken33 | ST7 (KY488585) | - | Southwest of Iran | Salehi et al., unpublished | 100 |
| Chicken34 | ST7 (KY488585) | - | Southwest of Iran | Salehi et al., unpublished | 100 |
| Chicken35 | ST7 (KY488585) | - | Southwest of Iran | Salehi et al., unpublished | 100 |
| Chicken37 | ST7 (KY488585) | - | Southwest of Iran | Salehi et al., unpublished | 100 |
| Chicken38 | ST7 (KY488585) | - | Southwest of Iran | Salehi et al., unpublished | 100 |
| Chicken39 | ST7 (KY488585) | - | Southwest of Iran | Salehi et al., unpublished | 100 |
| Sheep140 | ST10 (MF186708) | Wallaby | - | Betts et al., unpublished | 99.80 |
| Sheep145 | ST10 (MF186708) | Wallaby | - | Betts et al., unpublished | 99.80 |
| Sheep146 | ST10 (MF186708) | Wallaby | - | Betts et al., unpublished | 99.80 |
| Sheep147 | ST10 (MF186708) | Wallaby | - | Betts et al., unpublished | 99.80 |
| Sheep148 | ST10 (MF186708) | Wallaby | - | Betts et al., unpublished | 99.80 |
| Sheep149 | ST10 (MF186708) | Wallaby | - | Betts et al., unpublished | 99.80 |
| Buffalo10_2 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 99.08 |
| Buffalo10_4 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 99.08 |
| Buffalo10_5 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 99.08 |
| Buffalo7_2 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 98.90 |
| Buffalo7_3 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 98.72 |
| Buffalo6_10 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 98.72 |
| Buffalo10_3 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 98.75 |
| Buffalo1_5 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 99.08 |
| Buffalo6_6 | ST14 (MW682196) | Goat | Poland | Rudzinska, unpublished | 99.08 |
| Pig40 | ST5 (MK801418) | Pig | Romania | Wylezichet al., 2019 [29] | 100 |
| Pig42 | ST5 (MN493729) | Wild boar | South Korea | Lee and Kwak, unpublished | 100 |
| Pig43 | ST5 (MN493729) | Wild boar | South Korea | Lee and Kwak, unpublished | 100 |
| Pig45 | ST5 (MN493729) | Wild boar | South Korea | Lee and Kwak, unpublished | 100 |
| Pig46 | ST5 (MN493729) | Wild boar | South Korea | Lee and Kwak, unpublished | 100 |
| Pig50 | ST5 (MN493729) | Wild boar | South Korea | Lee and Kwak, unpublished | 100 |
| Goat51 | ST5 (MN493729) | Wild boar | South Korea | Lee and Kwak, unpublished | 100 |
| Deer55 | ST5 (MK801418) | Pig | Romania | Wylezichet al., 2019 [29] | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabrielli, S.; Palomba, M.; Furzi, F.; Brianti, E.; Gaglio, G.; Napoli, E.; Rinaldi, L.; Alburqueque, R.A.; Mattiucci, S. Molecular Subtyping of Blastocystis sp. Isolated from Farmed Animals in Southern Italy. Microorganisms 2021, 9, 1656. https://doi.org/10.3390/microorganisms9081656
Gabrielli S, Palomba M, Furzi F, Brianti E, Gaglio G, Napoli E, Rinaldi L, Alburqueque RA, Mattiucci S. Molecular Subtyping of Blastocystis sp. Isolated from Farmed Animals in Southern Italy. Microorganisms. 2021; 9(8):1656. https://doi.org/10.3390/microorganisms9081656
Chicago/Turabian StyleGabrielli, Simona, Marialetizia Palomba, Federica Furzi, Emanuele Brianti, Gabriella Gaglio, Ettore Napoli, Laura Rinaldi, Renato Aco Alburqueque, and Simonetta Mattiucci. 2021. "Molecular Subtyping of Blastocystis sp. Isolated from Farmed Animals in Southern Italy" Microorganisms 9, no. 8: 1656. https://doi.org/10.3390/microorganisms9081656
APA StyleGabrielli, S., Palomba, M., Furzi, F., Brianti, E., Gaglio, G., Napoli, E., Rinaldi, L., Alburqueque, R. A., & Mattiucci, S. (2021). Molecular Subtyping of Blastocystis sp. Isolated from Farmed Animals in Southern Italy. Microorganisms, 9(8), 1656. https://doi.org/10.3390/microorganisms9081656