
Development of a Novel Method for Identification of Alaria alata Mesocercariae by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Molecular Species Identification of the Samples
2.3. Development of a Protein Extraction Protocol for MALDI-TOF Mass Spectrometry
2.4. Generation of an AM-Specific MSP Library
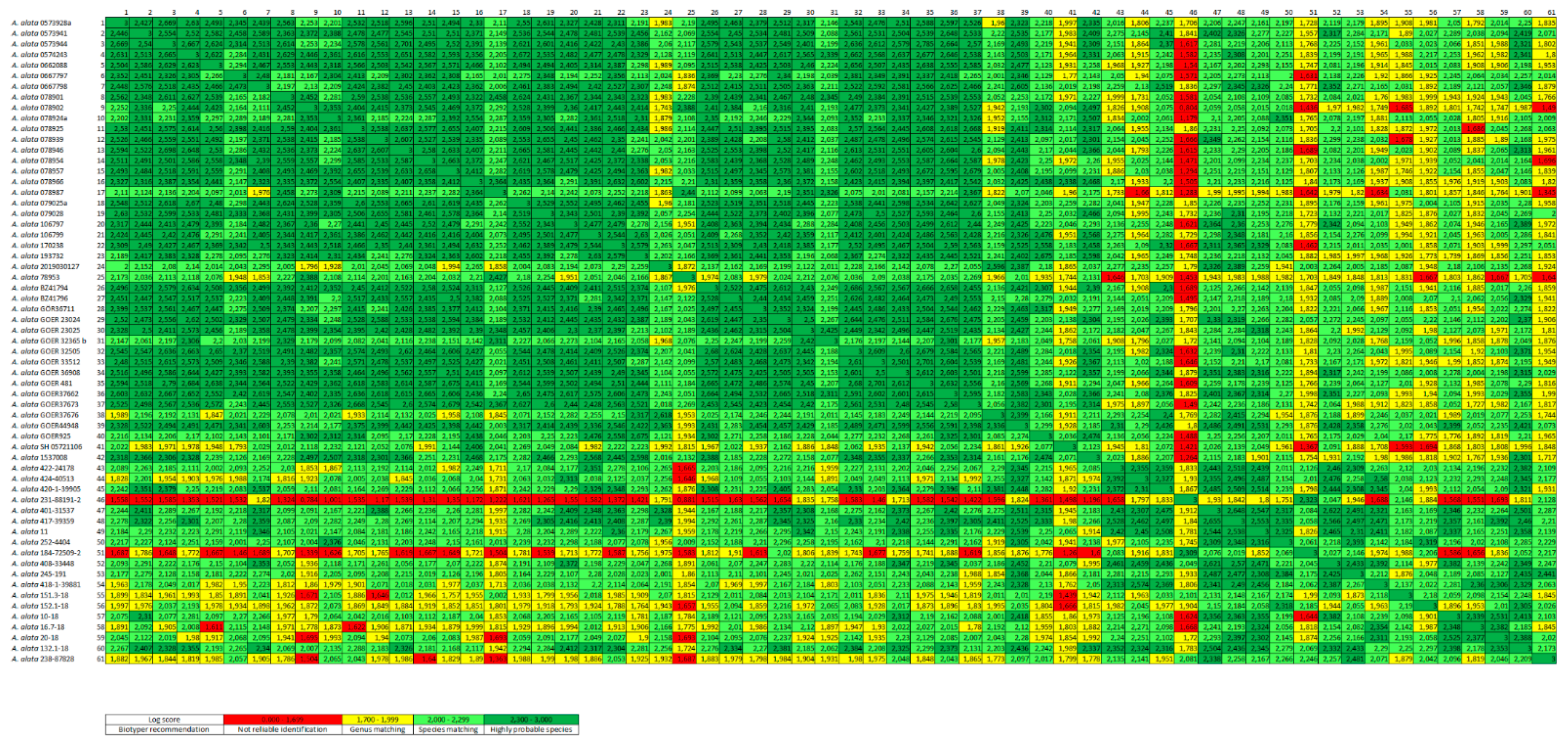
2.5. Evaluation of Generated Protein Spectra by Log Score Values
2.6. Modification of the MSP Identification Method in Biotyper 3 Software
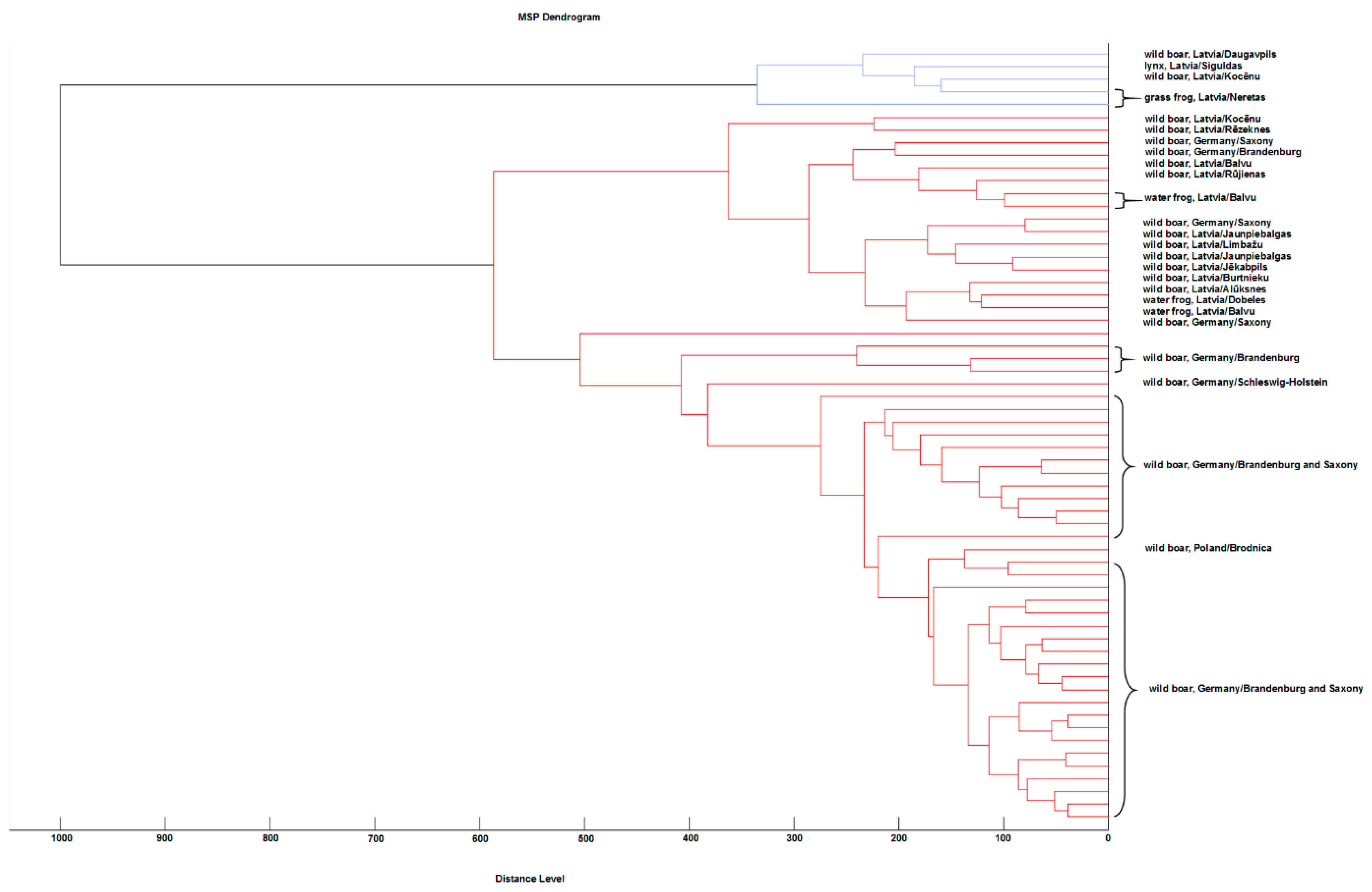
2.7. Cluster Analysis
2.8. Validation of the Developed Protocol Based on 10 AM
3. Results
3.1. Origin of the Samples
3.2. Molecular Analysis of the Samples
3.3. Creation of Main Spectra Profiles (MSPs)
3.4. Cluster Analysis of the Created MSPs
3.5. Sensitivity, Reproducibility and Repeatability of this MALDI-TOF Technique
4. Discussion
4.1. Background and Context to Previous Studies
4.2. Interpretation of the Cluster Analysis
4.3. Future Potential of the MALDI-TOF Technique
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- European Commission. Commission Implementing Regulation (EU) 2015/1375 of 10 August 2015 Laying down Specific Rules on Official Controls for Trichinella in Meat. Off. J. Eur. Union 2015, 58, 7–34. [Google Scholar]
- European Commission. Commission Implementing Regulation (EU) 2020/1478 of 14 October 2020 amending Implementing Regulation (EU) 2015/1375 as regards sampling, the reference method for detection and import conditions related to Trichinella control C.I.R. Off. J. Eur. Union 2020, 3, 1–3. [Google Scholar]
- Karadjian, G.; Kaestner, C.; Laboutière, L.; Adicéam, E.; Wagner, T.; Johne, A.; Thomas, M.; Polack, B.; Mayer-Scholl, A.; Vallée, I. A two-step morphology-PCR strategy for the identification of nematode larvae recovered from muscles after artificial digestion at meat inspection. Parasitol. Res. 2020, 119, 4113–4122. [Google Scholar] [CrossRef] [PubMed]
- Riehn, K.; Hamedy, A.; Grosse, K.; Wüste, T.; Lücker, E. Alaria alata in wild boars (Sus scrofa, Linnaeus, 1758) in the eastern parts of Germany. Parasitol. Res. 2012, 111, 1857–1861. [Google Scholar] [CrossRef]
- Sailer, A.; Glawischnig, W.; Irschik, I.; Lücker, E.; Riehn, K.; Paulsen, P. Findings of Alaria alata mesocercariae in wild boar in Austria: Current knowledge, identification of risk factors and discussion of risk management options. Wien. Tierärztliche Mon. 2012, 99, 346–352. [Google Scholar]
- Paulsen, P.; Forejtek, P.; Hutarova, Z.; Vodnansky, M. Alaria alata mesocercariae in wild boar (Sus scrofa, Linnaeus, 1758) in south regions of the Czech Republic. Vet. Parasitol. 2013, 197, 384–387. [Google Scholar] [CrossRef]
- Portier, J.; Vallée, I.; Lacour, S.A.; Martin-Schaller, R.; Ferté, H.; Durand, B. Increasing circulation of Alaria alata mesocercaria in wild boar populations of the Rhine valley, France, 2007–2011. Vet. Parasitol. 2014, 199, 153–159. [Google Scholar] [CrossRef]
- Riehn, K.; Lalkovski, N.; Hamedy, A.; Lücker, E. First detection of Alaria alata mesocercariae in wild boars (Sus scrofa Linnaeus, 1758) from Bulgaria. J. Helminthol. 2014, 88, 247–249. [Google Scholar] [CrossRef]
- Maleševic, M.; Smulders, F.J.M.; Petrovic, J.; Mirceta, J.; Paulsen, P. Alaria alata mesocercariae in wild boars (Sus scrofa) in northern Serbia after the flood disaster of 2014. Wien. Tierarztl. Mon. 2016, 103, 345–349. [Google Scholar]
- Ozoliņa, Z.; Mateusa, M.; Šuksta, L.; Liepiņa, L.; Deksne, G. The wild boar (Sus scrofa, Linnaeus1758) as an important reservoir host for Alaria alata in the Baltic region and potential risk of infection in humans. Vet. Parasitol. Reg. Stud. Rep. 2020, 22, 100485. [Google Scholar]
- Kästner, C.; Bier, N.S.; Mayer-Scholl, A.; Nöckler, K.; Richter, M.H.; Johne, A. Prevalence of Alaria alata mesocercariae in wild boars from Brandenburg, Germany. Parasitol. Res. 2021, 120, 2103–2108. [Google Scholar] [CrossRef]
- Strokowska, N.; Nowicki, M.; Klich, D.; Bełkot, Z.; Wiśniewski, J.; Didkowska, A.; Chyla, P.; Anusz, K. The occurrence of Alaria alata mesocercariae in wild boars (Sus scrofa) in north-eastern Poland. Int. J. Parasitol. Parasites Wildl. 2020, 12, 25–28. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Villa, L.; Riehn, K.; Hamedy, A.; Minazzi, S.; Olivieri, E.; Zanzani, S.A.; Manfredi, M.T. Occurrence of selected zoonotic food-borne parasites and first molecular identification of Alaria alata in wild boars (Sus scrofa) in Italy. Parasitol. Res. 2018, 117, 2207–2215. [Google Scholar] [CrossRef]
- Odening, K. Der “Dunckersche Muskelegel” kann experimentell auf Affen übertragen werden. Mon. Für Veterinärmedizin 1961, 16, 395–399. [Google Scholar]
- Möhl, K.; Große, K.; Hamedy, A.; Wüste, T.; Kabelitz, P.; Lücker, E. Biology of Alaria spp. and human exposition risk to Alaria mesocercariae-a review. Parasitol. Res. 2009, 105, 1–15. [Google Scholar] [CrossRef]
- Gottstein, B. Einstufung von Organismen. Modul 3: Parasiten. Stand Januar 2013. Herausgegeben vom Bundesamt für Umwelt BAFU, Bundesamt für Gesundheit. BAG: Bern, Switzerland, 2013. Available online: http://www.bafu.admin.ch/uv-1114-d (accessed on 19 February 2021).
- Federal Ministry of Labour and Social Affairs, Technische Regeln für Biologische Arbeitsstoffe (TRBA) 464 “Einstufung von Parasiten in Risikogruppen”, Ausgabe Juli 2013, 1. Änderung vom 10.11.2020, GMBI Nr. 45. 2020. Available online: https://www.baua.de/DE/Angebote/Rechtstexte-und-Technische-Regeln/Regelwerk/TRBA/pdf/TRBA464.pdf?__blob=publicationFile&v=3 (accessed on 26 April 2021).
- BVL, Zoonosen-Monitoring 2015. Gemeinsamer Bericht des Bundes und der Länder. Berichte zur Lebensmittelsicherheit, BVL-Report 11.2: 30-31. 2016. Available online: https://www.bvl.bund.de/SharedDocs/Downloads/01_Lebensmittel/04_Zoonosen_Monitoring/Zoonosen_Monitoring_Bericht_2015.pdf%3F__blob%3DpublicationFile%26v%3D6 (accessed on 24 August 2020).
- Gavrilović, P.; Pavlović, I.; Todorović, I. Alaria alata mesocercariae in domestic pigs and wild boars in South Banat, northern Serbia. Comp. Immunol. Microbiol. Infect. Dis. 2019, 63, 142–144. [Google Scholar] [CrossRef]
- Paulsen, P.; Ehebruster, J.; Irschik, I.; Lücker, E.; Riehn, K.; Winkelmayer, R.; Smulders, F.J.M. Findings of Alaria alata mesocercariae in wild boars (Sus scrofa) in eastern Austria. Eur. J. Wildl. Res. 2012, 58, 991–995. [Google Scholar] [CrossRef]
- Riehn, K.; Hamedy, A.; Große, K.; Zeitler, L.; Lücker, E. A novel detection method for Alaria alata mesocercariae in meat. Parasitol. Res. 2010, 107, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Riehn, K.; Hamedy, A.; Alter, T.; Lücker, E. Development of a PCR approach for differentiation of Alaria spp. mesocercariae. Parasitol. Res. 2011, 108, 1327–1332. [Google Scholar] [CrossRef]
- Bilska-Zając, E.; Marucci, G.; Piróg-Komorowska, A.; Cichocka, M.; Różycki, M.; Karamon, J.; Sroka, J.; Bełcik, A.; Mizak, I.; Cencek, T. Occurrence of Alaria alata in wild boars (Sus scrofa) in Poland and detection of genetic variability between isolates. Parasitol. Res. 2020, 20, 83–91. [Google Scholar] [CrossRef]
- Mayer-Scholl, A.; Murugaiyan, J.; Neumann, J.; Bahn, P.; Reckinger, S.; Nöckler, K. Rapid Identification of the Foodborne Pathogen Trichinella spp. by Matrix-Assisted Laser Desorption/Ionization Mass Spectrometry. PLoS ONE 2016, 11, e0152062. [Google Scholar] [CrossRef] [PubMed]
- Wieser, A.; Schneider, L.; Jung, J.; Schubert, S. MALDI-TOF MS in microbiological diagnostics-identification of microorganisms and beyond (mini review). Appl. Microbiol. Biotechnol. 2012, 93, 965–974. [Google Scholar] [CrossRef]
- Shannon, S.; Kronemann, D.; Patel, R.; Schuetz, A.N. Routine use of MALDI-TOF MS for anaerobic bacterial identification in clinical microbiology. Anaerobe 2018, 54, 191–196. [Google Scholar] [CrossRef]
- Croxatto, A.; Prod’hom, G.; Greub, G. Applications of MALDI-TOF mass spectrometry in clinical diagnostic microbiology. FEMS Microbiol. Rev. 2012, 36, 380–407. [Google Scholar] [CrossRef]
- Bredtmann, C.M.; Krücken, J.; Murugaiyan, J.; Kuzmina, T.; von Samson-Himmelstjerna, G. Nematode species identification—current status, challenges and future perspectives for Cyathostomins. Front. Cell. Infect. Microbiol. 2017, 7, 283. [Google Scholar] [CrossRef] [PubMed]
- Marzano, V.; Pane, S.; Foglietta, G.; Levi Mortera, S.; Vernocchi, P.; Onetti Muda, A.; Putignani, L. Mass Spectrometry Based-Proteomic Analysis of Anisakis spp.: A Preliminary Study towards a New Diagnostic Tool. Genes 2020, 11, 693. [Google Scholar] [CrossRef]
- Sy, I.; Margardt, L.; Ngbede, E.O.; Adah, M.I.; Yusuf, S.T.; Keiser, J.; Utzinger, J.; Poppert, S.; Becker, S.L. Identification of Adult Fasciola spp. Using Matrix-Assisted Laser/Desorption Ionization Time-of-Flight (MALDI-TOF) Mass Spectrometry. Microorganisms 2021, 9, 82. [Google Scholar] [CrossRef]
- Huguenin, A.; Depaquit, J.; Villena, I.; Ferté, H. MALDI-TOF mass spectrometry: A new tool for rapid identification of cercariae (Trematoda, Digenea). Parasite 2019, 26, 11. [Google Scholar] [CrossRef]
- Karadjian, G.; Bilska-Zając, E.; Bahn, P.; Py, J.-S.; Johne, A.; Gassilloud, B.; Różycki, M.; Cencek, T.; Mayer-Scholl, A.; Vallée, I. Species identification of Trichinella originated from various host and different geographical location by MALDI-TOF. Exp. Parasitol. 2020, 213, 107890. [Google Scholar] [CrossRef] [PubMed]
- Ozoliņa, Z.; Bagrade, G.; Deksne, G. First confirmed case of Alaria alata mesocercaria in Eurasian lynx (Lynx lynx) hunted in Latvia. Parasitol. Res. 2020, 119, 759–762. [Google Scholar] [CrossRef] [PubMed]
- Ozoliņa, Z.; Deksne, G.; Pupins, M.; Gravele, E.; Gavarane, I.; Kirjušina, M. Alaria alata mesocercariae prevalence and predilection sites in amphibians in Latvia. Parasitol. Res. 2020, 120, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Guillen, J. FELASA guidelines and recommendations. J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 311–321. [Google Scholar] [PubMed]
- Justine, J.-L.; Briand, M.J.; Bray, R.A. A quick and simple method, usable in the field, for collecting parasites in suitable condition for both morphological and molecular studies. Parasitol. Res. 2012, 111, 341–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, M.I.; El-Shahawy, I.S.; Abdelkader, H.S. Studies on some fish parasites of public health importance in the southern area of Saudi Arabia. Rev. Bras. Parasitol. Veterinária 2014, 23, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, P.D.; Cribb, T.; Tkach, V.; Bray, R.; Littlewood, D. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 2003, 33, 733–755. [Google Scholar] [CrossRef]
- Portier, J.; Jouet, D.; Vallee, I.; Ferté, H. Detection of Planorbis planorbis and Anisus vortex as first intermediate hosts of Alaria alata (Goeze, 1792) in natural conditions in France: Molecular evidence. Vet. Parasitol. 2012, 190, 151–158. [Google Scholar] [CrossRef]
- Marucci, G.; Interisano, M.M.; La Rosa, G.; Pozio, E. Molecular identification of nematode larvae different from those of the Trichinella genus detected by muscle digestion. Vet. Parasitol. 2013, 194, 117–120. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. | Sample Number | Year of Sampling | Host Species | Origin of the Host | Host Tissue Used for AM Isolation | Number of MSPs | Log Scores for MSPs from the Same Host |
|---|---|---|---|---|---|---|---|
| 1 | A. alata 0573928a | 2018 | wild boar | Germany, Brandenburg | tongue | 2 | 2.474 |
| 2 | A. alata 0573941 | 2018 | wild boar | Germany, Brandenburg | tongue | 1 | no value |
| 3 | A. alata 0573944 | 2018 | wild boar | Germany, Brandenburg | tongue | 1 | no value |
| 4 | A. alata 0574243 | 2018 | wild boar | Germany, Brandenburg | tongue | 2 | 2.592–2.593 |
| 5 | A. alata 0662088 | 2019 | wild boar | Germany, Brandenburg | foreleg muscles | 1 | no value |
| 6 | A. alata 0667797 | 2019 | wild boar | Germany, Brandenburg | tongue | 1 | no value |
| 7 | A. alata 0667798 | 2019 | wild boar | Germany, Brandenburg | tongue | 3 | 2.062–2.599 |
| 8 | A. alata 078901 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 5 | 2.383–2.660 |
| 9 | A. alata 078902 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 5 | 2.316–2.638 |
| 10 | A. alata 078924a | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 5 | 2.011–2.562 |
| 11 | A. alata 078925 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 1 | no value |
| 12 | A. alata 078939 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 4 | 2.308–2.550 |
| 13 | A. alata 078946 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 1 | no value |
| 14 | A. alata 078954 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 5 | 2.318–2.623 |
| 15 | A. alata 078957 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 4 | 2.127–2.430 |
| 16 | A. alata 078966 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 1 | no value |
| 17 | A. alata 078987 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 5 | 2.341–2.767 |
| 18 | A. alata 079025a | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 2 | 2.625–2.632 |
| 19 | A. alata 079028 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 4 | 2.337–2.647 |
| 20 | A. alata 106797 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 5 | 2.013–2.597 |
| 21 | A. alata 106799 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 1 | no value |
| 22 | A. alata 170238 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 1 | no value |
| 23 | A. alata 193732 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 1 | no value |
| 24 | A. alata 2019030127 | 2020 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue, diaphragm, chewing muscles | 3 | 2.488–2.681 |
| 25 | A. alata 78953 | 2019 | wild boar | Germany, Brandenburg | tongue, abdominal fat tissue | 5 | 2.108–2.675 |
| 26 | A. alata BZ41794 | 2019 | wild boar | Germany, Saxony | tongue, foreleg muscles | 1 | no value |
| 27 | A. alata BZ41796 | 2019 | wild boar | Germany, Saxony | tongue, foreleg muscles | 1 | no value |
| 28 | A. alata GOER36711 | 2018 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 1 | no value |
| 29 | A. alata GOER23024 | 2019 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 5 | 2.012–2.491 |
| 30 | A. alata GOER23025 | 2019 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 5 | 2.368–2.545 |
| 31 | A. alata GOER32365b | 2019 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 3 | 2.492–2.707 |
| 32 | A. alata GOER32505 | 2018 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 1 | no value |
| 33 | A. alata GOER33512 | 2019 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 4 | 2.468–2.575 |
| 34 | A. alata GOER36908 | 2019 | wild boar | Germany, Saxony | diaphragm, chewing muscles, abdominal fat tissue | 5 | 2.167–2.497 |
| 35 | A. alata GOER481 | 2018 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 1 | no value |
| 36 | A. alata GOER37662 | 2019 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 4 | 2.455–2.619 |
| 37 | A. alata GOER37673 | 2020 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 4 | 2.305–2.587 |
| 38 | A. alata GOER37676 | 2020 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 1 | no value |
| 39 | A. alata GOER44948 | 2020 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 3 | 2.668–2.747 |
| 40 | A. alata GOER925 | 2020 | wild boar | Germany, Saxony | diaphragm, stomach, chewing muscles | 5 | 2.000–2.508 |
| 41 | A. alata SH05721106 | 2019 | wild boar | Germany, Schleswig-Holstein | diaphragm | 1 | no value |
| 42 | A. alata 1537008 | 2019 | wild boar | Poland, Brodnica | diaphragm | 5 | 2.175–2.579 |
| 43 | A. alata 422-24178 | 2020 | wild boar | Latvia, Alūksnes | diaphragm | 1 | no value |
| 44 | A. alata 424-40513 | 2020 | wild boar | Latvia, Balvu | diaphragm | 1 | no value |
| 45 | A. alata 420-1-39905 | 2020 | wild boar | Latvia, Burtnieku | diaphragm | 2 | 2.428–2.444 |
| 46 | A. alata 231-88191-2 | 2018 | wild boar | Latvia, Daugavpils | diaphragm, tongue | 1 | no value |
| 47 | A. alata 401-31537 | 2020 | wild boar | Latvia, Jaunpiebalgas | diaphragm | 1 | no value |
| 48 | A. alata 417-39359 | 2020 | wild boar | Latvia, Jaunpiebalgas | diaphragm | 1 | no value |
| 49 | A. alata 11 | 2020 | wild boar | Latvia, Jēkabpils | diaphragm | 2 | 2.520–2.523 |
| 50 | A. alata 252-4404 | 2019 | wild boar | Latvia, Kocēnu | diaphragm, tongue | 1 | no value |
| 51 | A. alata 184-72509-2 | 2018 | wild boar | Latvia, Kocēnu | diaphragm | 1 | no value |
| 52 | A. alata 408-33448 | 2020 | wild boar | Riga, Limbažu | diaphragm | 1 | no value |
| 53 | A. alata 245-191 | 2019 | wild boar | Latvia, Rēzeknes | diaphragm, tongue | 1 | no value |
| 54 | A. alata 418-1-39881 | 2020 | wild boar | Latvia, Rūjienas | diaphragm | 9 | 2.181–2.745 |
| 55 | A. alata 151.3-18 | 2018 | common frog | Latvia, Neretas | serous coat of the internal organs | 1 | no value |
| 56 | A. alata 152.1-18 | 2018 | common frog | Latvia, Neretas | head | 1 | no value |
| 57 | A. alata 10-18 | 2018 | water frog | Latvia, Balvu | serous coat of the internal organs | 1 | no value |
| 58 | A. alata 16.7-18 | 2018 | water frog | Latvia, Balvu | inner organs | 1 | no value |
| 59 | A. alata 20-18 | 2018 | water frog | Latvia, Balvu | serous coat of the internal organs | 1 | no value |
| 60 | A. alata 132.1-18 | 2018 | water frog | Latvia, Dobeles | head | 1 | no value |
| 61 | A. alata 238-87828 | 2018 | lynx | Latvia, Siguldas | tongue, different muscles | 1 | no value |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kästner, C.; Bahn, P.; Schönfelder, R.; Ozoliņa, Z.; Alksne, L.; Richter, M.H.; Deksne, G.; Mayer-Scholl, A.; Johne, A. Development of a Novel Method for Identification of Alaria alata Mesocercariae by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Microorganisms 2021, 9, 1664. https://doi.org/10.3390/microorganisms9081664
Kästner C, Bahn P, Schönfelder R, Ozoliņa Z, Alksne L, Richter MH, Deksne G, Mayer-Scholl A, Johne A. Development of a Novel Method for Identification of Alaria alata Mesocercariae by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Microorganisms. 2021; 9(8):1664. https://doi.org/10.3390/microorganisms9081664
Chicago/Turabian StyleKästner, Carolyn, Peter Bahn, Ralph Schönfelder, Zanda Ozoliņa, Laura Alksne, Martin Heinrich Richter, Gunita Deksne, Anne Mayer-Scholl, and Annette Johne. 2021. "Development of a Novel Method for Identification of Alaria alata Mesocercariae by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry" Microorganisms 9, no. 8: 1664. https://doi.org/10.3390/microorganisms9081664
APA StyleKästner, C., Bahn, P., Schönfelder, R., Ozoliņa, Z., Alksne, L., Richter, M. H., Deksne, G., Mayer-Scholl, A., & Johne, A. (2021). Development of a Novel Method for Identification of Alaria alata Mesocercariae by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Microorganisms, 9(8), 1664. https://doi.org/10.3390/microorganisms9081664