
Effect of Methionine Supplementation on Rumen Microbiota, Fermentation, and Amino Acid Metabolism in In Vitro Cultures Containing Nitrate

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates
2.2. Treatments
2.3. In Vitro Batch Culture
2.4. Determination of Total Gas and Methane Production
2.5. Determination of Reducing Equivalents (Expressed as H2)
2.6. Sampling and Determination of In Vitro Fermentation Parameters
2.7. Determination of Amino Acid Concentration
2.8. DNA Extraction and Quantification of Microbial Populations
2.9. Metagenomic Biomarker Identification and Functional Prediction
2.10. Statistical Analysis
3. Results
3.1. Hydrogen Balance, Total Gas, and CH4 Production
3.2. Rumen Fermentation Parameters
3.3. Ruminal Amino Acids
3.4. Rumen Microbial Populations
3.5. Rumen Bacterial Diversity
3.5.1. Alpha and Beta Diversity Parameters
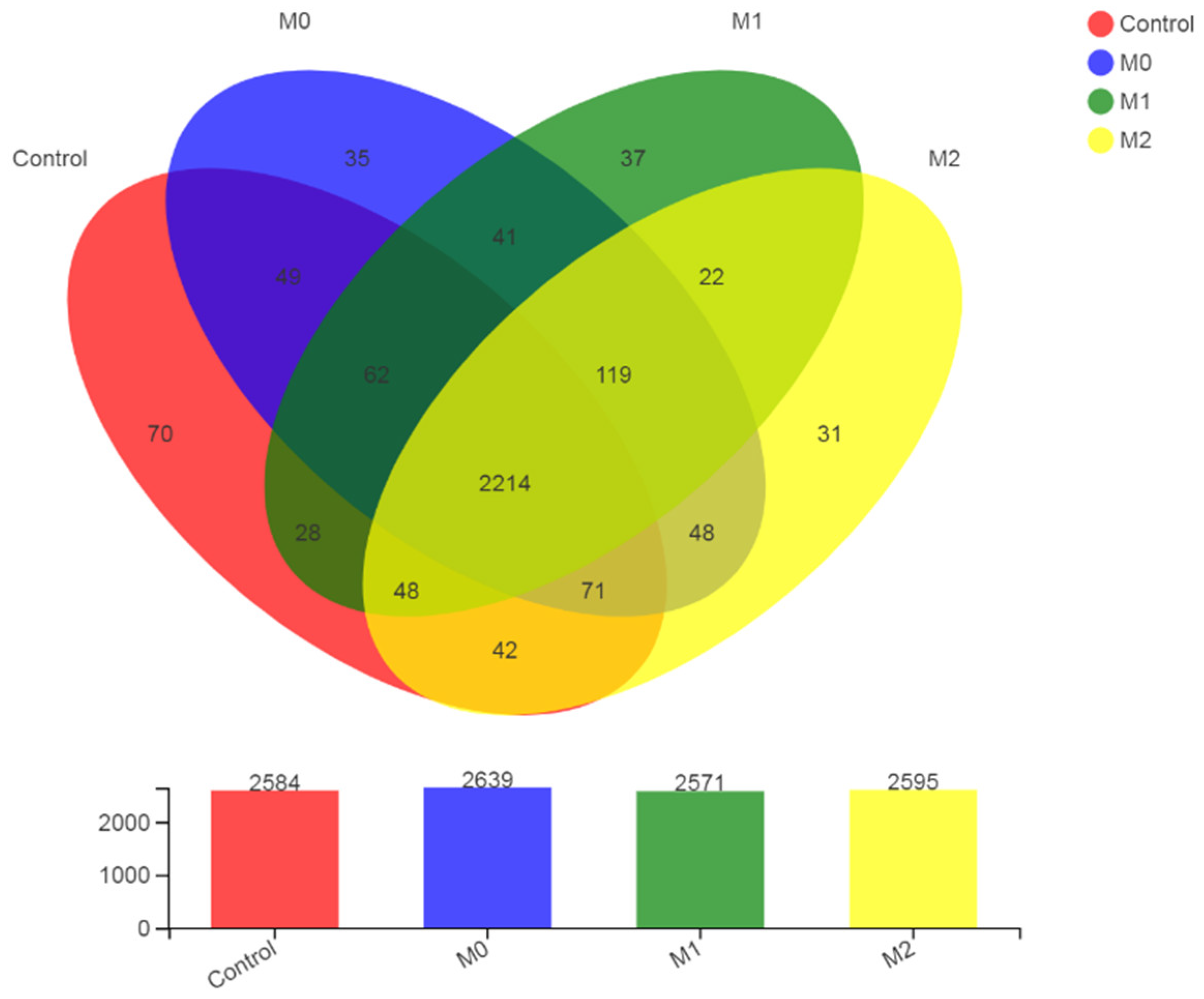
3.5.2. OTU Statistics
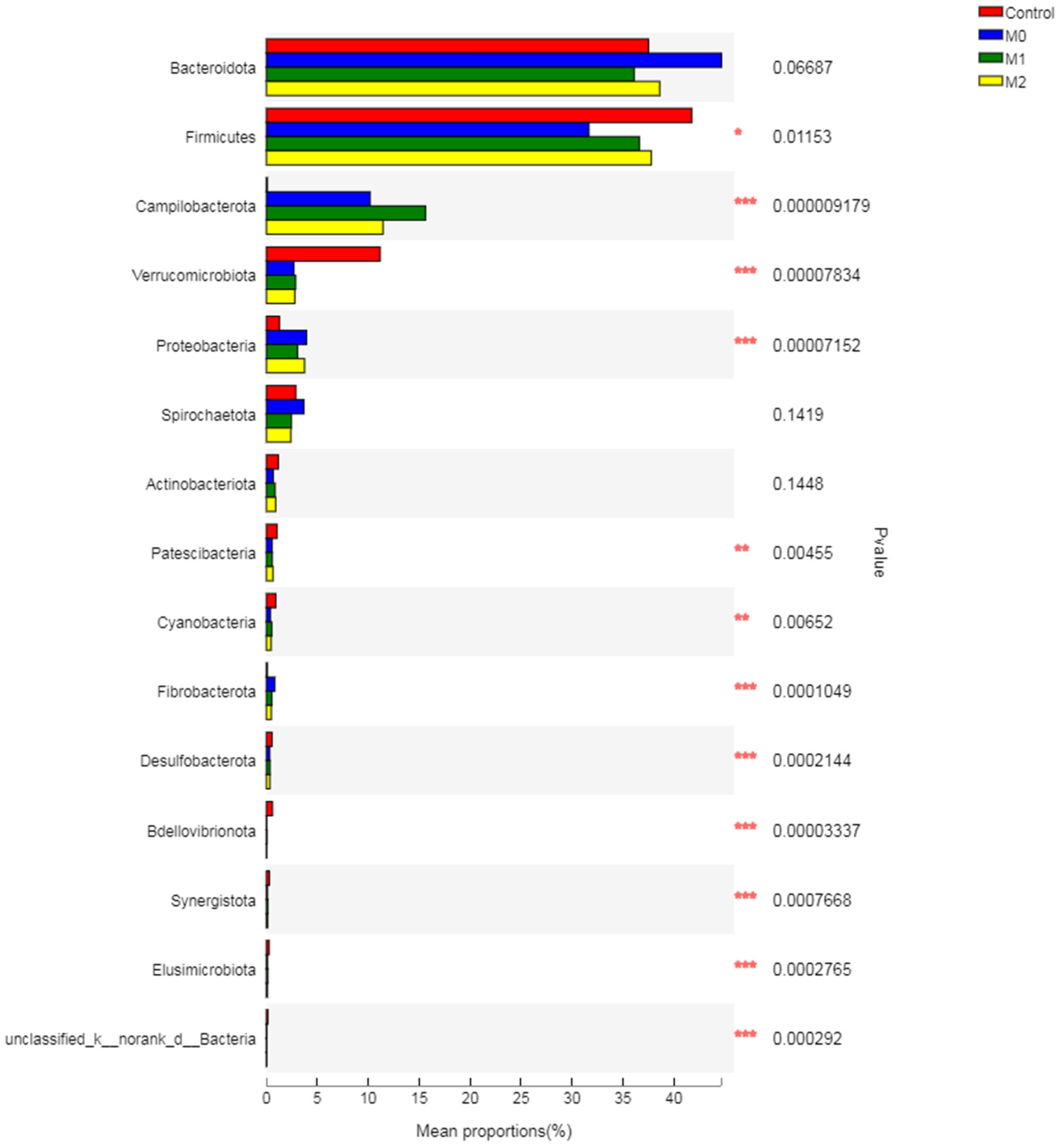
3.5.3. Relative Abundance of Bacterial Phyla
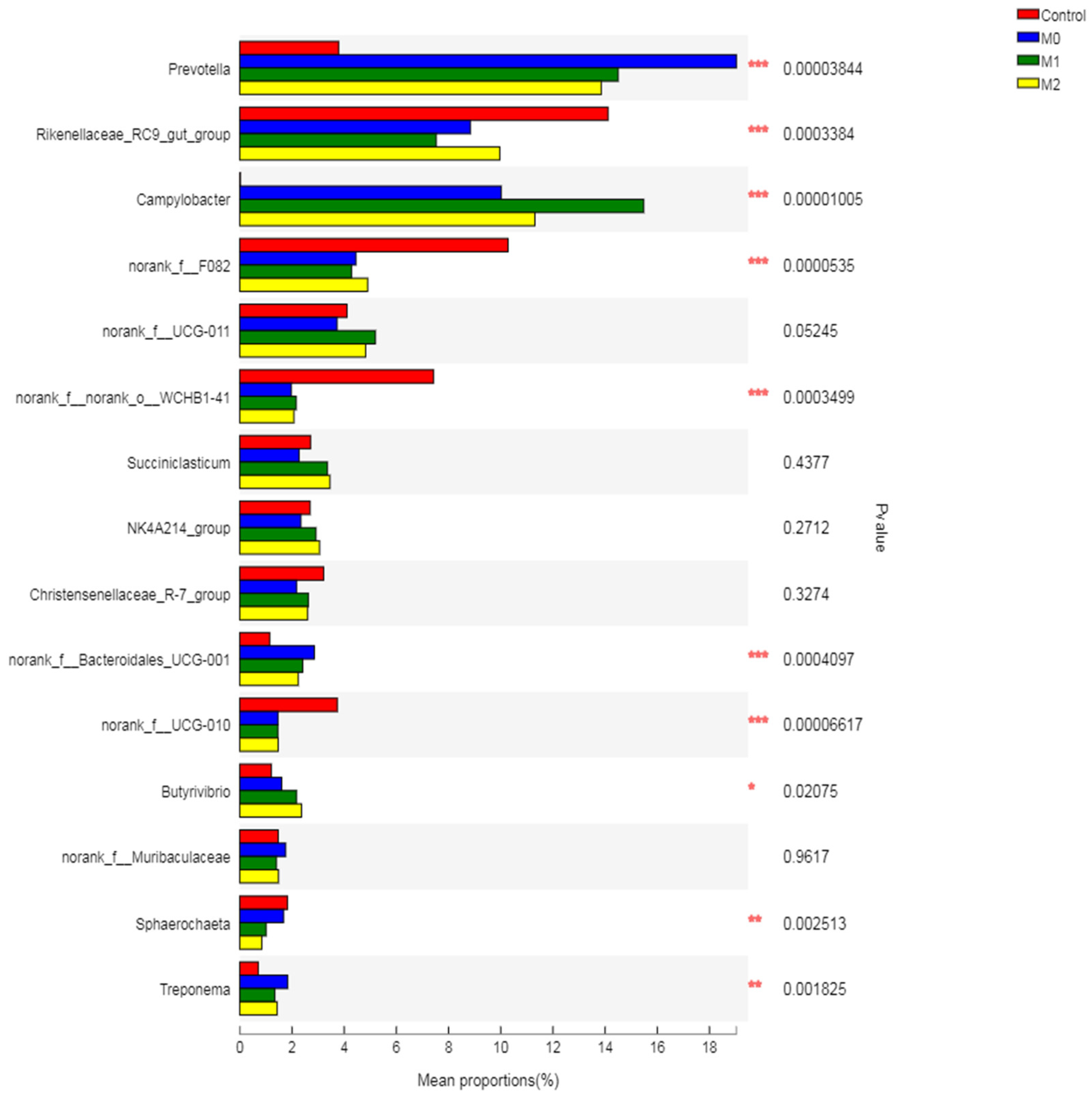
3.5.4. Relative Abundance of Bacterial Genera
3.6. Biomarker Bacteria Taxa and Metagenomic Functional Profile
3.7. Association of Rumen Bacteria with Ruminal Gas, VFA, and Amino Acid Contents
4. Discussion
4.1. Effect of Treatment on H2 Balance, Cumulative Gas, and CH4 Production
4.2. Rumen Fermentation Parameters
4.3. Ruminal Amino Acids
4.4. Rumen Microbial Populations
4.5. Ruminal Bacterial Diversity
4.6. Biomarker Bacterial Genera and Functional Prediction Profile
4.7. Association of Rumen Bacteria with Ruminal Gas, VFA, and Amino Acid Contents
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ungerfeld, E.M. Shifts in metabolic hydrogen sinks in the methanogenesis-inhibited ruminal fermentation: A meta-analysis. Front. Microbiol. 2015, 6, 37. [Google Scholar] [CrossRef]
- Czerkawski, J.W. An Introduction to Rumen Studies; Pergamon Press: Oxford, UK, 1986. [Google Scholar]
- Chalupa, W. Manipulating rumen fermentation. J. Anim. Sci. 1977, 45, 585–599. [Google Scholar] [CrossRef]
- Eckard, R.; Grainger, C.; De Klein, C. Options for the abatement of methane and nitrous oxide from ruminant production: A review. Livestock Sci. 2010, 130, 47–56. [Google Scholar] [CrossRef]
- Guyader, J.; Tavendale, M.; Martin, C.; Muetzel, S. Dose-response effect of nitrate on hydrogen distribution between rumen fermentation end products: An in vitro approach. Anim. Prod. Sci. 2016, 56, 224–230. [Google Scholar] [CrossRef]
- Ungerfeld, E.M.; Aedo, M.F.; Martínez, E.D.; Saldivia, M. Inhibiting methanogenesis in rumen batch cultures did not increase the recovery of metabolic hydrogen in microbial amino acids. Microorganisms 2019, 7, 115. [Google Scholar] [CrossRef] [Green Version]
- Hulshof, R.; Berndt, A.; Gerrits, W.; Dijkstra, J.; Van Zijderveld, S.; Newbold, J.; Perdok, H. Dietary nitrate supplementation reduces methane emission in beef cattle fed sugarcane-based diets. J. Anim. Sci. 2012, 90, 2317–2323. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.G.; Broderick, G.A. A 100-Year Review: Protein and amino acid nutrition in dairy cows. J. Dairy Sci. 2017, 100, 10094–10112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: An overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Schaefer, D.; Guo, X.; Ren, L.; Meng, Q. Use of nitrate-nitrogen as a sole dietary nitrogen source to inhibit ruminal methanogenesis and to improve microbial nitrogen synthesis in vitro. Asian Austr. J. Anim. Sci. 2009, 22, 542–549. [Google Scholar] [CrossRef]
- Yang, C.; Rooke, J.A.; Cabeza, I.; Wallace, R.J. Nitrate and Inhibition of Ruminal Methanogenesis: Microbial Ecology, Obstacles, and Opportunities for Lowering Methane Emissions from Ruminant. Livestock. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Granja-Salcedo, Y.T.; Fernandes, R.M.; Araujo, R.C.; Kishi, L.T.; Berchielli, T.T.; Resende, F.D.; Berndt, A.; Siqueir, G.R. Long-term encapsulated nitrate supplementation modulates rumen microbial diversity and rumen fermentation to reduce methane emission in grazing steers. Front Microbiol. 2019, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- Leng, R.A. The Potential of Feeding Nitrate to Reduce Enteric Methane Production in Ruminants; Report to Department of Climate Change; Commonwealth Government: Canberra, Australia, 2008; p. 82.
- Menke, K.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.W.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Meng, Q.; Yu, Z. Effects of methanogenic inhibitors on methane production and abundances of methanogens and cellulolytic bacteria in in vitro ruminal cultures. Appl. Environ. Microbiol. 2011, 77, 2634–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weilian, H.; Jiakun, W.; Jianmin, L. Rapid gas chromatogram determination of methane, organic acid in vitro ruminal fermentation products. J. Zhejiang Univ. 2006, 32, 217–221. [Google Scholar]
- Anderson, R.C.; Huwe, J.K.; Smith, D.J.; Stanton, T.B.; Krueger, N.A.; Callaway, T.R.; Edrington, T.S.; Harvey, R.B.; Nisbet, D.J. Effect of nitroethane, dimethyl-2-nitroglutarate and 2-nitro-methyl-propionate on ruminal methane production and hydrogen balance in vitro. Bioresour. Technol. 2010, 101, 5345–5349. [Google Scholar] [CrossRef]
- Demeyer, D. Quantitative aspects of microbial metabolism in the rumen and hindgut. In Rumen Microbial Metabolism and Ruminant Digestion; Jouany, J.P., Ed.; INRA Editions: Paris, France, 1991; pp. 217–237. [Google Scholar]
- Weatherburn, M. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Li, X.; Yan, C.; Choi, S.; Long, R.; Jin, G.; Song, M.K. Effects of addition level and chemical type of propionate precursors in dicarboxylic acid pathway on fermentation characteristics and methane production by rumen microbes in vitro. Asian Austr. J. Anim. Sci. 2009, 22, 82–89. [Google Scholar] [CrossRef]
- Qin, W. Determination of rumen volatile fatty acids by means of gas chromatography. J. Nanjing Agric. Coll. 1982, 4, 110–116. [Google Scholar]
- Joyce, R.; Kuziene, V.; Zou, X.; Wang, X.; Pullen, F.; Loo, R.L. Development and validation of an ultra-performance liquid chromatography quadrupole time of flight mass spectrometry method for rapid quantification of free amino acids in human urine. Amino Acids 2016, 48, 219–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef]
- Ebeid, H.M.; Hassan, F.U.; Li, M.; Peng, L.; Peng, K.; Liang, X.; Yang, C. Camelina sativa L. oil mitigates enteric in vitro methane production, modulates ruminal fermentation and ruminal bacterial diversity in buffaloes. Front. Vet. Sci. 2020, 7, 550. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Poulsen, M.; Schwab, C.; Jensen, B.B.; Engberg, R.M.; Spang, A.; Canibe, N.; Højberg, O.; Milinovich, G.; Fragner, L.; Schleper, C. Methylotrophic methanogenic Thermoplasmata implicated in reduced methane emissions from bovine rumen. Nat. Commun. 2013, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- El-Zaiat, H.; Araujo, R.; Soltan, Y.; Morsy, A.; Louvandini, H.; Pires, A.; Patino, H.O.; Correa, P.S.; Abdalla, A. Encapsulated nitrate and cashew nut shell liquid on blood and rumen constituents, methane emission, and growth performance of lambs. J. Anim. Sci. 2014, 92, 2214–2224. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Schaefer, D.; Guo, W.; Ren, L.; Meng, Q. Comparisons of in vitro nitrate reduction, methanogenesis, and fermentation acid profile among rumen bacterial, protozoal and fungal fractions. Asian Australas. J. Anim. Sci. 2011, 24, 471–478. [Google Scholar] [CrossRef]
- Lin, M.; Schaefer, D.; Zhao, G.; Meng, Q. Effects of nitrate adaptation by rumen inocula donors and substrate fiber proportion on in vitro nitrate disappearance, methanogenesis, and rumen fermentation. Acid. Anim. 2013, 7, 1099–1105. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Xu, X.; Cao, Y.; Cai, C.; Cui, H.; Yao, Y. Nitrate decreases methane production also by increasing methane oxidation through stimulating NC10 population in ruminal culture. AMB Express. 2017, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pal, K.; Patra, A.; Sahoo, A.; Mandal, G. Effect of nitrate and fumarate in Prosopis cineraria and Ailanthus excelsa leaves-based diets on methane production and rumen fermentation. Small Rumin. Res. 2014, 121, 168–174. [Google Scholar] [CrossRef]
- Patra, A.K.; Yu, Z. Effects of essential oils on methane production and fermentation by, and abundance and diversity of, rumen microbial populations. Appl. Environ. Microbiol. 2012, 78, 4271–4280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, C.; Meng, Q.; Hou, X.; Ren, L.; Zhou, Z. Response of ruminal fermentation, methane production and dry matter digestibility to microbial source and nitrate addition level in an in vitro incubation with rumen microbes obtained from wethers. J. Anim. Vet. Adv. 2012, 11, 3334–3341. [Google Scholar]
- Zhou, Z.; Yu, Z.; Meng, Q. Effects of nitrate on methane production, fermentation, and microbial populations in in vitro ruminal cultures. Bioresour. Technol. 2012, 103, 173–179. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.; Adesogan, A.T.; Yang, W.; Lee, C.; et al. Special topics—Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef] [Green Version]
- Latham, E.A.; Anderson, R.C.; Pinchak, W.E.; Nisbet, D.J. Insights on alterations to the rumen ecosystem by nitrate and nitrocompounds. Front. Microbiol. 2016, 7, 228. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.; Flint, H.; Bryant, M. The rumen bacteria. In The Rumen Microbial Ecosystem; Springer: Berlin, Germany, 1997; pp. 10–72. [Google Scholar]
- Min, B.R.; Castleberry, L.; Allen, H.; Parker, D.; Waldrop, H.; Brauer, D.; Willis, W. Associative effect of wet distillers’ grains plus solubles and tannin-rich peanut skin supplementation on in vitro rumen fermentation, greenhouse gas emissions, and microbiome changes. J. Anim. Sci. 2019, 97, 4668–4681. [Google Scholar] [CrossRef]
- Patra, A.; Yu, Z. Effects of garlic oil, nitrate, saponin and their combinations supplemented to different substrates on in vitro fermentation, ruminal methanogenesis, and abundance and diversity of microbial populations. J. Appl. Microbiol. 2015, 119, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Van Zijderveld, S.; Gerrits, W.; Apajalahti, J.; Newbold, J.; Dijkstra, J.; Leng, R.; Perdok, H. Nitrate and sulfate: Effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J. Dairy Sci. 2010, 93, 5856–5866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyader, J.; Ungerfeld, E.M.; Beauchemin, K.A. Redirection of metabolic hydrogen by inhibiting methanogenesis in the rumen simulation technique (RUSITEC). Front. Microbiol. 2017, 8, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillman, A.D.; Sheriha, G.; Sirny, R. Nitrate reduction studies with sheep. J. Anim. Sci. 1965, 24, 1140–1146. [Google Scholar] [CrossRef] [Green Version]
- Bach, A.; Stern, M.D. Effects of different levels of methionine and ruminally undegradable protein on the amino acid profile of effluent from continuous culture fermenters. J. Anim. Sci. 1999, 77, 3377–3384. [Google Scholar] [CrossRef]
- Chung, Y.H.; Bateman, H.G., II; Williams, C.C.; Stanley, C.C.; Gantt, D.T.; Braud, T.W.; Southern, L.L.; Ward, J.D.; Hoyt, P.G.; Sod, G.A. Effects of methionine and lysine on fermentation in vitro and in vivo, nutrient flow to the intestine, and milk production. J. Dairy Sci. 2006, 89, 1613–1620. [Google Scholar] [CrossRef]
- Janssen, P.H. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar] [CrossRef]
- Troy, S.; Duthie, C.; Hyslop, J.; Roehe, R.; Ross, D.; Wallace, R.; Waterhouse, A.; Rooke, J. Effectiveness of nitrate addition and increased oil content as methane mitigation strategies for beef cattle fed two contrasting basal diets. J. Anim. Sci. 2015, 93, 1815–1823. [Google Scholar] [CrossRef]
- Ballard, F. Supply and utilization of acetate in mammals. Am. J. Clin. Nutr. 1972, 25, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Meng, Q.; Ren, L.; Liu, W.; Zhang, X.; Huo, Y.; Zhou, Z. Effects of nitrate addition on rumen fermentation, bacterial biodiversity and abundance. Asian Australas. J. Anim. Sci. 2015, 28, 1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Hellwing, A.; Cao, B.; Weisbjerg, M. The acute effect of addition of nitrate on in vitro and in vivo methane emission in dairy cows. Anim. Prod. Sci. 2014, 54, 1432–1435. [Google Scholar]
- Takahashi, J.; Young, B. Prophylactic effect of L-cysteine on nitrate-induced alterations in respiratory exchange and metabolic rate in sheep. Anim. Feed Sci. Technol. 1991, 35, 105–113. [Google Scholar] [CrossRef]
- Carver, L.; Pfander, W. Some metabolic aspects of urea and/or potassium nitrate utilization by sheep. J. Anim. Sci. 1974, 38, 410–416. [Google Scholar] [CrossRef]
- Li, L.; Davis, J.; Nolan, J.; Hegarty, R. An initial investigation on rumen fermentation pattern and methane emission of sheep offered diets containing urea or nitrate as the nitrogen source. Anim. Prod. Sci. 2012, 52, 653–658. [Google Scholar] [CrossRef]
- Silivong, P.; Preston, T.; Van Man, N. Effect of supplements of potassium nitrate or urea as sources of NPN on methane production in an in vitro system using molasses and Paper mulberry or Muntingia foliages as the substrate. Gas 2012, 17, 12–14. [Google Scholar]
- Sophea, I.; Preston, T. Effect of different levels of supplementary potassium nitrate replacing urea on growth rates and methane production in goats fed rice straw, mimosa foliage and water spinach. Livest. Res. Rural Dev. 2011, 23, 71. [Google Scholar]
- Thanh, V.D.; Thu, N.; Preston, T. Effect of potassium nitrate or urea as NPN source and levels of Mangosteen peel on in vitro gas and methane production using molasses, Operculina turpethum and Brachiaria mutica as substrate. Livest. Res. Rural Dev. 2012, 24, 63. [Google Scholar]
- Russell, J.; Wallace, R. Energy-yielding and energy-consuming reactions. In the Rumen Microbial Ecosystem; Springer: Berlin/Heidelberg, Germany, 1997; pp. 246–282. [Google Scholar]
- Wang, R.; Wang, M.; Ungerfeld, E.M.; Zhang, X.M.; Long, D.L.; Mao, H.X.; Deng, J.P.; Bannink, A.; Tan, Z.L. Nitrate improves ammonia incorporation into rumen microbial protein in lactating dairy cows fed a low-protein diet. J. Dairy Sci. 2018, 101, 9789–9799. [Google Scholar] [CrossRef] [Green Version]
- Allison, M.; Reddy, C. Adaptations of Gastrointestinal Bacteria in Response to Changes in Dietary Oxalate and Nitrate; American Society for Microbiology: Washington, DC, USA, 1984. [Google Scholar]
- Hasan, S.; Hall, J. The physiological function of nitrate reduction in Clostridium perfringens. Microbiology 1975, 87, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Madigan, M.T.; Martinko, J.M.; Parker, J. Brock Biology of Microorganisms; Prentice hall Upper Saddle River: Upper Saddle River, NJ, USA, 1997; Volume 11. [Google Scholar]
- Argyle, J.; Baldwin, R. Effects of amino acids and peptides on rumen microbial growth yields. J. Dairy Sci. 1989, 72, 2017–2027. [Google Scholar] [CrossRef]
- Cotta, M.A.; Russell, J.B. Effect of peptides and amino acids on efficiency of rumen bacterial protein synthesis in continuous culture. J. Dairy Sci. 1982, 65, 226–234. [Google Scholar] [CrossRef]
- Van Kessel, J.S.; Russell, J.B. The effect of amino nitrogen on the energetic of ruminal bacteria and its impact on energy spilling. J. Dairy Sci. 1996, 79, 1237–1243. [Google Scholar] [CrossRef]
- Firkins, J.L.; Yu, Z.; Morrison, M. Ruminal Nitrogen Metabolism: Perspectives for Integration of Microbiology and Nutrition for Dairy. J. Dairy Sci. 2007, 90, E1–E16. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B.; Strobell, H.J. Microbial energetics. In Quantitative Aspects of Ruminant Digestion and Metabolism; Dijkstra, J., Forbes, J.M., France, J., Eds.; CAB International: Wallingford, UK, 1993; pp. 165–186. [Google Scholar]
- Atasoglu, C.E.N.G.İ.Z.; Guliye, A.Y. Use of stable isotopes to measure de novo synthesis and turnover of amino acid-C and—N in mixed micro-organisms from the sheep rumen in vitro. Br. J. Nutr. 2004, 91, 253–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carro, M.; Miller, E. Effect of supplementing a fibre basal diet with different nitrogen forms on ruminal fermentation and microbial growth in an in vitro semi-continuous culture system (RUSITEC). Br. J. Nutr. 1999, 82, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Chikunya, S.; Newbold, C.; Rode, L.; Chen, X.; Wallace, R. Influence of dietary rumen-degradable protein on bacterial growth in the rumen of sheep receiving different energy sources. Anim. Feed Sci. Technol. 1996, 63, 333–340. [Google Scholar] [CrossRef]
- Zhang, H.L.; Chen, Y.; Xu, X.L.; Yang, Y.X. Effects of branched-chain amino acids on in vitro ruminal fermentation of wheat straw. Asian Austr. J. Anim. Sci. 2013, 26, 523. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Yu, Z. Effective reduction of enteric methane production by a combination of nitrate and saponin without adverse effect on feed degradability, fermentation, or bacterial and archaeal communities of the rumen. Bioresour. Technol. 2013, 148, 352–360. [Google Scholar] [CrossRef]
- Sharma, B.K.; Erdman, R.A. Effects of high amounts of dietary choline supplementation on duodenal choline flow and production response of dairy cows. J. Dairy Sci. 1988, 71, 2670–2676. [Google Scholar] [CrossRef]
- Marais, J.P.; Therion, J.J.; Mackie, R.; Kistner, A.; Dennison, C. Effect of nitrate and its reduction products on the growth and activity of the rumen microbial population. Br. J. Nutr. 1988, 59, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Patton, R.; McCarthy, R.; Keske, L.; Griel, L.J.; Baumgardt, B. Effect of Feeding Methionine Hydroxy Analog on the Concentration of Protozoa in Rumen of Sheep. J. Dairy Sci. 1970, 53, 933–935. [Google Scholar] [CrossRef]
- Patton, R.A.; McCarthy, R.D.; Griel, L.C.J. Lipid synthesis by rumen microorganisms. II. Further characterization of the effects of methionine. J. Dairy Sci. 1970, 53, 460. [Google Scholar] [CrossRef]
- Yao, Z.; Vance, D.E. The active synthesis of phosphatidylcholine is required for very low density lipoprotein secretion from rat hepatocytes. J. Biol. Chem. 1988, 263, 2998–3004. [Google Scholar] [CrossRef]
- Williams, A.G.; Coleman, G.S. The Rumen Protozoa; Springer: New York, NY, USA, 1992. [Google Scholar]
- Eugène, M.; Archimède, H.; Sauvant, D. Quantitative meta-analysis on the effects of defaunation of the rumen on growth, intake and digestion in ruminants. Livest. Prod. Sci. 2004, 85, 81–97. [Google Scholar] [CrossRef]
- Hillman, K.; Williams, A.G.; Lloyd, D. Postprandial variations in endogenous metabolic activities of ovine rumen ciliate protozoa. Anim. Feed. Sci. Technol. 1995, 52, 237–247. [Google Scholar] [CrossRef]
- Iwamoto, M.; Asanuma, N.; Hino, T. Effects of nitrate combined with fumarate on methanogenesis, fermentation, and cellulose digestion by mixed ruminal microbes in vitro. Nihon Chikusan Gakkaiho 1999, 70, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, M.; Asanuma, N.; Hino, T. Effects of pH and electron donors on nitrate and nitrite reduction in ruminal microbiota. Nihon Chikusan Gakkaiho 2001, 72, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.; Fernandez, G.; Shinkai, T.; Mitsumori, M.; Mc-Sweeney, C.S. Metagenomic analysis of the rumen microbial community following inhibition of methane formation by a halogenated methane analog. Front. Microbiol. 2015, 6, 1087. [Google Scholar] [CrossRef]
- Miller, W.G.; On, S.L.; Wang, G.; Fontanoz, S.; Lastovica, A.J.; Mandrell, R.E. Extended multilocus sequence typing system for Campylobacter coli, C. lari, C. upsaliensis, and C. helveticus. J. Clin. Microbiol. 2005, 43, 2315–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkhill, J.; Wren, B.; Mungall, K.; Ketley, J.; Churcher, C.; Basham, D.; Holroyd, S.; Chillingworth, T.; Davies, R.M.; Feltwell, T.; et al. The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 2000, 403, 665–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, E.; Frías, J.E.; Rubio, L.M.; Herrero, A. Photosynthetic nitrate assimilation in cyanobacteria. Photosyn. Res. 2005, 83, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, R.; Schnürer, A.; Arthurson, V.; Bertilsson, J. Methanogenic population and CH4 production in Swedish dairy cows fed different levels of forage. App. Environ. Microbiol. 2012, 78, 6172–6179. [Google Scholar] [CrossRef] [Green Version]
- Tapio, I.; Snelling, T.J.; Strozzi, F.; Wallace, R.J. The ruminal microbiome associated with methane emissions from ruminant livestock. J. Anim. Sci. Biotechnol. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Wallace, R.J.; Rooke, J.A.; McKain, N.; Duthie, C.-A.; Hyslop, J.J.; Ross, D.W.; Waterhouse, A.; Watson, M.; Roehe, R. The rumen microbial metagenome associated with high methane production in cattle. BMC Genom. 2015, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kamke, J.; Kittelmann, S.; Soni, P.; Li, Y.; Tavendale, M.; Ganesh, S.; Janssen, P.H.; Shi, W.; Froula, J.; Rubin, E.M. Rumen metagenome and metatranscriptome analyses of low methane yield sheep reveals a Sharpea-enriched microbiome characterised by lactic acid formation and utilisation. Microbiome 2016, 4, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Álvaro, M.; Auffret, M.D.; Stewart, R.D.; Dewhurst, R.J.; Duthie, C.-A.; Rooke, J.A.; Wallace, R.J.; Shih, B.; Freeman, T.C.; Watson, M. Identification of complex rumen microbiome interaction within diverse functional niches as mechanisms affecting the variation of methane emissions in bovine. Front. Microbiol. 2020, 11, 659. [Google Scholar] [CrossRef]
- Strobel, H.J. Vitamin B12-dependent propionate production by the ruminal bacterium Prevotella ruminicola 23. Appl. Environ. Microbiol. 1992, 58, 2331–2333. [Google Scholar] [CrossRef] [Green Version]
- Purushe, J.; Fouts, D.E.; Morrison, M.; White, B.A.; Mackie, R.I.; Coutinho, P.M.; Henrissat, B.; Nelson, K.E. Comparative genome analysis of Prevotella ruminicola and Prevotella bryantii: Insights into their environmental niche. Microb. Ecol. 2010, 60, 721–729. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Forano, E.; Martin, C.; Newbold, C.J. Microbial ecosystem and methanogenesis in ruminants. Animal 2010, 4, 1024–1036. [Google Scholar] [CrossRef] [Green Version]
- Mitsumori, M.; Shinkai, T.; Takenaka, A.; Enishi, O.; Higuchi, K.; Kobayashi, Y.; Nonaka, I.; Asanuma, N.; Denman, S.E.; McSweeney, C.S. Responses in digestion, rumen fermentation and microbial populations to inhibition of methane formation by a halogenated methane analogue. Br. J. Nutr. 2012, 108, 482–491. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content |
|---|---|
| Elephant grass (%) | 90 |
| Concentrate mixture (%) * | 10 |
| Chemical Composition | |
| Dry matter (%) | 20 |
| Crude protein (%) | 9.03 |
| Neutral detergent fiber (%) | 75.34 |
| Acid detergent fiber (%) | 46.01 |
| Ash (%) | 9.72 |
| Gross energy (kcal/kg DM) | 4.69 |
| Control | M0 (0% Methionine) | M1 (0.28% Methionine) | M2 (1.12% Methionine) | |
|---|---|---|---|---|
| Substrate | 0.5 g | 0.5 g | 0.5 g | 0.5 g |
| NaNO3 | 0 | 0.06 g | 0.06 g | 0.06 g |
| Methionine | 0 | 0 | 0.0014 g | 0.0056 g |
| Parameter | Control | M0 (0%) | M1 (0.28%) | M2 (1.12%) | SEM | p Value |
|---|---|---|---|---|---|---|
| CH4 (mL/g DM) | 25.69 a | 5.88 b | 5.99 b | 6.31 b | 0.601 | 0.001 |
| Total gas (mL/g DM) | 205.2 a | 110.8 b | 117.2 b | 113.2 b | 4.001 | 0.001 |
| Reductive hydrogen | ||||||
| H2 produced (mmol) | 7.72 a | 6.26 b | 6.40 b | 6.25 b | 0.141 | 0.001 |
| H2 utilized (mmol) | 5.65 a | 2.99 b | 3.01 b | 3.05 b | 1.093 | 0.001 |
| H2 recovery (%) | 73.40 a | 47.88 c | 47.10 c | 48.88 c | 0.085 | 0.001 |
| Control | M0 | M1 | M2 | SEM | p Value | |
|---|---|---|---|---|---|---|
| pH | 6.70 c | 6.84 a | 6.78 b | 6.80 a | 0.023 | 0.001 |
| Acetate (mmole/L) | 32.32 | 32.21 | 32.81 | 31.94 | 0.547 | 0.748 |
| Propionate (mmole/L) | 17.54 a | 15.01 b | 14.67 b | 15.03 b | 0.336 | 0.001 |
| Isobutyrate (mmole/L) | 1.10 a | 0.59 b | 0.58 b | 0.56 b | 0.021 | 0.001 |
| Butyrate (mmole/L) | 9.81 a | 5.17 b | 5.58 b | 5.27 b | 0.238 | 0.001 |
| Isovalerate (mmole/L) | 2.31 a | 1.35 b | 1.19 b | 1.30 b | 0.066 | 0.001 |
| Valerate (mmole/L) | 1.31 a | 0.80 b | 0.81 b | 0.77 b | 0.025 | 0.001 |
| MCP (mg/mL) | 3.43 a | 4.33 b | 4.49 b | 4.47 b | 0.093 | 0.001 |
| NH3-N (mg/mL) | 18.32 | 17.81 | 18.45 | 18.78 | 0.318 | 0.272 |
| TVFA (mmole/L) | 64.38 a | 55.13 b | 55.64 b | 54.87 b | 0.957 | 0.001 |
| A/P | 1.84 a | 2.17 b | 2.24 b | 2.13 b | 0.047 | 0.001 |
| Amino Acid | Control | M0 | M1 | M2 | SEM | p Value |
|---|---|---|---|---|---|---|
| Alanine | 327.72 b | 753.56 a | 615.46 a | 296.67 b | 82.01 | 0.001 |
| Valine | 62.62 b | 121.60 ab | 141.98 a | 65.68 b | 19.73 | 0.014 |
| Histidine | 52.19 bc | 76.62 a | 60.15 ab | 35.86 c | 7.69 | 0.006 |
| Arginine | 30.89 b | 53.31 a | 49.89 a | 26.99 b | 5.57 | 0.003 |
| Glycine | 184.32 bc | 363.54 a | 303.09 ab | 136.81 c | 42.16 | 0.002 |
| Glutamine | 409.13 b | 822.82 a | 681.91 a | 370.95 b | 78.99 | 0.001 |
| Glutamate | 786.52 b | 1402.61 a | 1212.22 a | 652.38 b | 134.52 | 0.001 |
| Proline | 36.54 bc | 67.70 a | 61.17 ab | 23.97 c | 8.65 | 0.003 |
| Leucine | 151.82 b | 359.94 a | 296.21 a | 131.80 b | 43.08 | 0.001 |
| Lysine | 472.00 b | 970.89 a | 807.53 a | 431.42 b | 93.51 | 0.001 |
| Methionine | 96.22 b | 222.14 a | 192.11 a | 99.98 b | 23.19 | 0.001 |
| Tryptophan | 71.19 b | 127.54 a | 109.68 a | 61.12 b | 13.26 | 0.003 |
| Phenylalanine | 170.84 b | 355.42 a | 313.22 a | 153.91 b | 39.72 | 0.001 |
| Threonine | 166.87 bc | 358.22 a | 289.51 ab | 131.18 c | 43.19 | 0.002 |
| Isoleucine | 152.68 b | 357.16 a | 298.67 a | 133.66 b | 42.63 | 0.001 |
| Tyrosine | 45.99 b | 88.84 a | 84.05 a | 37.78 b | 10.24 | 0.001 |
| Serine | 188.18 b | 379.51 a | 322.97 a | 155.95 b | 41.06 | 0.001 |
| Aspartic acid | 141.08 b | 306.21 a | 254.98 a | 124.40 b | 35.05 | 0.002 |
| Asparagine | 8.187 b | 7.507 b | 19.431 a | 17.181 a | 2.553 | 0.003 |
| Cysteine | 57.86 | 69.48 | 70.70 | 65.89 | 3.90 | 0.105 |
| Essential Amino acids 1 | 1396.44 bc | 2949.53 a | 2509.06 ab | 1244.60 c | 314.96 | 0.001 |
| Non-Essential Amino acids 2 | 2216.41 bc | 4315.09 a | 3675.87 ab | 1908.97 c | 432.85 | 0.001 |
| Total Amino Acids 3 | 3612.85 bc | 7264.62 a | 6184.93 ab | 3153.57 c | 747.33 | 0.001 |
| Parameter | Control | M0 | M1 | M2 | SEM | p Value |
|---|---|---|---|---|---|---|
| Bacteria | 11.13 b | 11.44 a | 11.49 a | 11.47 a | 0.040 | 0.001 |
| Fungi | 8.40 | 8.64 | 8.70 | 8.57 | 0.087 | 0.109 |
| Protozoa | 7.31 b | 7.54 ab | 7.89 a | 7.66 ab | 0.125 | 0.020 |
| Methanogens | 9.32 b | 9.56 a | 9.60 a | 9.56 a | 0.046 | 0.001 |
| Parameter | Control | M0 | M1 | M2 | SEM | p Value |
|---|---|---|---|---|---|---|
| Sobs | 1740.20 | 1686.70 | 1638.90 | 1645.90 | 26.68 | 0.051 |
| Shannon | 6.08 a | 5.59 b | 5.36 b | 5.55 b | 0.07 | 0.001 |
| Simpson | 0.006 c | 0.021 b | 0.032 a | 0.022 ab | 0.002 | 0.001 |
| ACE | 2035.80 | 2043.20 | 2013.00 | 2018.50 | 25.93 | 0.841 |
| Chao | 2068.60 | 2063.10 | 2041.40 | 2053.60 | 29.78 | 0.927 |
| Shannon evenness | 0.82 a | 0.75 b | 0.72 b | 0.75 b | 0.01 | 0.001 |
| Simpson evenness | 0.10 a | 0.04 b | 0.02 b | 0.04 a | 0.01 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, F.-u.; Guo, Y.; Li, M.; Tang, Z.; Peng, L.; Liang, X.; Yang, C. Effect of Methionine Supplementation on Rumen Microbiota, Fermentation, and Amino Acid Metabolism in In Vitro Cultures Containing Nitrate. Microorganisms 2021, 9, 1717. https://doi.org/10.3390/microorganisms9081717
Hassan F-u, Guo Y, Li M, Tang Z, Peng L, Liang X, Yang C. Effect of Methionine Supplementation on Rumen Microbiota, Fermentation, and Amino Acid Metabolism in In Vitro Cultures Containing Nitrate. Microorganisms. 2021; 9(8):1717. https://doi.org/10.3390/microorganisms9081717
Chicago/Turabian StyleHassan, Faiz-ul, Yanxia Guo, Mengwei Li, Zhenhua Tang, Lijuan Peng, Xin Liang, and Chengjian Yang. 2021. "Effect of Methionine Supplementation on Rumen Microbiota, Fermentation, and Amino Acid Metabolism in In Vitro Cultures Containing Nitrate" Microorganisms 9, no. 8: 1717. https://doi.org/10.3390/microorganisms9081717
APA StyleHassan, F. -u., Guo, Y., Li, M., Tang, Z., Peng, L., Liang, X., & Yang, C. (2021). Effect of Methionine Supplementation on Rumen Microbiota, Fermentation, and Amino Acid Metabolism in In Vitro Cultures Containing Nitrate. Microorganisms, 9(8), 1717. https://doi.org/10.3390/microorganisms9081717