Metacestodes of Elasmobranch Tapeworms in Octopus vulgaris (Mollusca, Cephalopoda) from Central Mediterranean—SEM and Molecular Data


 ,
, 
Abstract
:Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preservation
2.2. Scanning Electron Microscopy (SEM)
2.3. Molecular Analysis
3. Results
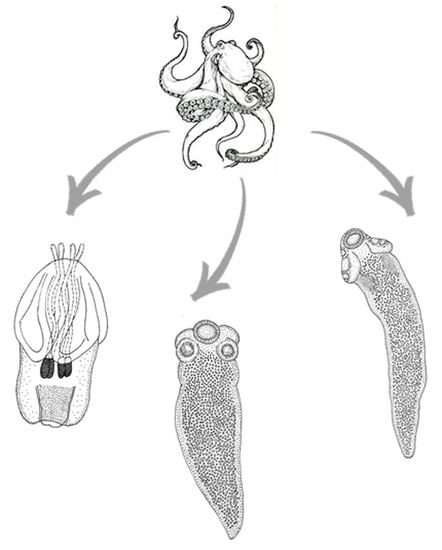
3.1. Morphological Description
3.1.1. Acanthobothrium sp. (Onchoproteocephalidea: Onchobothriidae) plerocercoids
3.1.2. Anthobothrium sp. (“Tetraphyllidea”: Clade 2) plerocercoids
3.1.3. Nybelinia sp. (Trypanorhyncha: Tentaculariidae) plerocercoid
3.2. Molecular Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gestal, C.; Abollo, E.; Arias, C.; Pascual, S.S.E.M. study of the plerocercoid larval Phyllobothrium sp. (Tetraphyllidea, Phyllobothriidae) and Nybelinia lingualis (Trypanothyncha, Tentaculariidae), cestode parasite in Octopus vulgaris (Mollusca, Cephalopoda) off Vigo estuary. Iberus 1998, 16, 125–132. [Google Scholar]
- Brickle, P.; Olson, P.D.; Littlewood, D.T.J.; Bishop, A.; Arkhipkin, A.I. Parasites of Loligo gahi from waters off the Falkland Islands, with a phylogenetically based identification of their cestode larvae. Can J. Zool. 2001, 79, 2289–2296. [Google Scholar] [CrossRef] [Green Version]
- Shukhgalter, O.A.; Nigmatullin, C.M. Parasitic helminths of jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) in open waters of the central east Pacific. Fish Res. 2001, 54, 95–110. [Google Scholar] [CrossRef]
- Pardo-Gandarillas, M.C.; Lohrmann, K.B.; Valdivia, A.L.; Ibáñez, C.M. First record of parasites of Dosidicus gigas (d’Orbigny, 1835) (Cephalopoda: Ommastrephidae) from the Humboldt Current system off Chile. Rev. Biol. Mar. Oceanogr 2009, 44, 397–408. [Google Scholar]
- Naidenova, N.N.; Nigmatullin, C.M.; Gaevskaya, A.V. The helminthofauna and parasite-host relationships of the squid Sthenoteuthis oualaniensis in the Indian Ocean and Red Sea. In Proceedings of the Symposium on Parasitology and Pathology of Marine Organisms, Leningrad, Russia, 13–16 October 1981; pp. 69–74. (In Russian). [Google Scholar]
- Naidenova, N.N.; Nigmatullin, C.M.; Gaevskaya, A.V. The helminth fauna and host-parasite relations of squids Sthenoteuthis oualaniensis (Lesson) (Cephalopoda, Ommastrephidae) in the Indian Ocean and the Red Sea. In Parasitology and Pathology of Marine Organisms of the World Ocean; Hargis, W.J., Ed.; NOAA Scientific Publications Office: Seattle, WA, USA, 1985; Volume 25, pp. 113–116. [Google Scholar]
- Williams, H.; Jones, A. Parasitic Worms of Fish; Taylor & Francis: London, UK, 1994. [Google Scholar]
- Jensen, K.; Bullard, S.A. Characterization of a diversity of tetraphyllidean and rhinebothriidean cestode larval types, with comments on host associations and life-cycles. Int. J. Parasitol. 2010, 40, 889–910. [Google Scholar] [CrossRef] [PubMed]
- Caira, J.N.; Reyda, F.B. Eucestoda (true tapeworms). In Marine Parasitology; Rhode, K., Ed.; CABI Publishing: New York, NY, USA, 2005; pp. 92–104. [Google Scholar]
- Palm, H.W. The Trypanorhyncha Diesing, 1863; PKSPL-IPB Press: Bogor, Indonesia, 2004. [Google Scholar]
- Tedesco, P.; Bevilacqua, S.; Fiorito, G.; Terlizzi, A. Global patterns of parasite diversity in cephalopods. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pascual, S.; Abollo, E.; Mladineo, I.; Gestal, C. Metazoa and Related Diseases. In Handbook of Pathogens and Diseases in Cephalopods; Gestal, C., Pascual, S., Guerra, A., Fiorito, G., Vieites, J.M., Eds.; Springer: Cham, Switzerland, 2019; pp. 169–179. [Google Scholar]
- Ruhnke, T.R. Tapeworms of elasmobranchs (Part III) A Monograph on the Phyllobothriidae (Platyhelminthes, Cestoda). Bull Univ. Nebr. State Mus. 2011, 25, 1–208. [Google Scholar]
- Hochberg, F.G. Diseases of Mollusca Cephalopoda-Diseases caused by Protistans and Metazoans. In Diseases of Marine Animals; Kinne, O., Ed.; Biologische Anstalt Helgoland: Hamburg, Germany, 1990; Volume 3, pp. 47–227. [Google Scholar]
- Riser, N.W. Studies on the Tetraphyllidea. Ph.D. Thesis, Stanford University, Palo Alto, CA, USA, 1949. [Google Scholar]
- Cake, E.W. A key to larval cestodes of shallow-water, benthic mollusks of the northern Gulf of Mexico. Proc. Helminthol. Soc. Wash. 1976, 43, 160–171. [Google Scholar]
- González, A.F.; Pascual, S.; Gestal, C.; Abollo, E.; Guerra, A. What makes a cephalopod a suitable host for a parasite? The case of Galician waters. Fish. Res. 2003, 60, 177–183. [Google Scholar] [CrossRef]
- Dollfus, R.P. Cestodes des invertébrés marins et thalassoides. In Faune de France: Cestodes; Joyeux, C., Baer, J.G., Eds.; Paul Lechevalier et Fils: Paris, France, 1936; Volume 30, pp. 509–539. [Google Scholar]
- Dollfus, R.P. Copépodes, Isopodes et Helminths parasites de Cephalopodes de la Méditerranée et de l’Atlantique Européen. Faune Mar Pyrénées-Orientales 1958, 1, 61–72. [Google Scholar]
- Dollfus, R.P. Enumération des cestodes du plancton et des invertébrés marins. (6econtribution). Ann. Parasit. Hum. Comp. 1964, 39, 329–379. [Google Scholar] [CrossRef]
- Chervy, L. The terminology of larval cestodes or metacestodes. Syst. Parasitol. 2002, 52, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Chervy, L. Unified terminology for cestode microtriches: A proposal from the International Workshops on Cestode Systematics in 2002–2008. Folia. Parasit. 2009, 56, 199–230. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, A.E.; Olson, P.D.; Littlewood, D.T.J. Utility of complete large and small subunit rRNA genes in resolving the phylogeny of the Neodermata (Platyhelminthes): Implications and a review of the cercomer theory. Biol. J. Linn. Soc. 2003, 78, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 5–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 547–1549. [Google Scholar] [CrossRef]
- Dollfus, R.P. Énumération des cestodes du plancton et des invertébrés marins. II. Mollusques céphalopodes et Crustacés. Ann. Parasit. Hum. Comp. 1923, 1, 363–394. [Google Scholar] [CrossRef]
- Dollfus, R.P. Addendum à mon «Énumération des cestodes du plancton et des invertébrés marins». Ann. Parasit. Hum. Comp. 1929, 7, 325–347. [Google Scholar] [CrossRef]
- Young, R.T. Cestodes of sharks and rays in Southern California. Proc. Helminthol. Soc. Wash. 1954, 21, 106–112. [Google Scholar]
- Goldstein, R.J. The genus Acanthobothrium van Beneden, 1849 (Cestoda: Tetraphyllidea). J. Parasitol. 1967, 53, 455–483. [Google Scholar] [CrossRef]
- Holland, N.D.; Wilson, N.G. Molecular identification of larvae of a tetraphyllidean tapeworm (Platyhelminthes: Eucestoda) in a razor clam as an alternative intermediate host in the life cycle of Acanthobothrium brevissime. J. Parasitol. 2009, 95, 1215–1218. [Google Scholar] [CrossRef]
- Zaragoza-Tapia, F.; Pulido-Flores, G.; Gardner, S.L.; Monks, S. Host relationships and geographic distribution of species of Acanthobothrium Blanchard, 1848 (Onchoproteocephalidea, Onchobothriidae) in elasmobranchs: A metadata analysis. ZooKeys 2020, 940, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, H.S.; Poulin, R. Evolution of interspecific variation in size of attachment structures in the large tapeworm genus Acanthobothrium (Tetraphyllidea: Onchobothriidae). Parasitology 2010, 137, 1707–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caira, J.N.; Jensen, K. Planetary Biodiversity Inventory (2008–2017): Tapeworms from Vertebrate Bowels of the Earth; Natural History Museum, The University of Kansas: Lawrence, KS, USA, 2017. [Google Scholar]
- Zschokke, F. Recherches sur la structure anatomique des cestodes. Mem. Inst. Nat. Genevois. 1888, 17, 1–396. [Google Scholar]
- Baer, J.G. Contributions à l’étude des cestodes de sélaciens. I-IV. Bull. Soc. Neuchâtel Sci. Nat. 1948, 71, 63–122. [Google Scholar]
- Yamaguti, S. Systema Helminthum. vol. II. The Cestodes of Vertebrates; Interscience Publishers: New York, NY, USA, 1959. [Google Scholar]
- Healy, C.J.; Caira, J.N.; Jensen, K.; Webster, B.L.; Littlewood, D.T.J. Proposal for a new tapeworm order, Rhinebothriidea. Int. J. Parasitol. 2009, 39, 497–511. [Google Scholar] [CrossRef]
- Caira, J.N.; Jensen, K.; Waeschenbach, A.; Olson, P.D.; Littlewood, D.T.J. Orders out of chaos–molecular phylogenetics reveals the complexity of shark and stingray tapeworm relationships. Int. J. Parasitol. 2014, 44, 55–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avdeeva, N.V.; Avdeev, V.V. Peculiarities of morphogenesis of adhesive organs of some plerocercoids of the collective genus Scolex (Tetraphyllidea) and their identification. Parazitologiya 1980, 14, 242–250. [Google Scholar]
- Di Cave, D.; Orecchia, P.; Ortis, M.; Paggi, L. Metazoi parassiti di alcuni elasmobranchi del mar Tirreno. Biol. Mar. Mediterr. 2003, 10, 249–252. [Google Scholar]
- Orecchia, P.; Paggi, L.; Minervini, R.; Di Cave, D. La parassitofauna delle specie ittiche strascicabili presenti alla foce del fiume Tevere. Oebalia 1985, 11, 623–632. [Google Scholar]
- Bona, F.; Buriola, E.; Cerioni, S.; di Cave, D.; Orecchia, P.; Paggi, L. Cestoda, Monogenea. In Checklist Delle Specie Della Fauna Italiana; Minelli, A., Ruffo, S., La Posta, S., Eds.; Calderini: Bologna, Italy, 1995. [Google Scholar]
- Paggi, L. Cestoda. Biol. Mar. Mediterr. 2008, 15 (Suppl. 1), 150–154. [Google Scholar]
- Capapé, C.; Crouzet, S.; Clement, C.; Vergne, Y.; Guelorget, O. Diet of the marbled electric ray Torpedo marmorata (Chondrichthyes: Torpedinidae) off the Languedocian coast (Southern France, Northern Mediterranean). Ann. Ser. Hist. Nat. 2007, 17, 17–22. [Google Scholar]
- Miller, M.H. Status Review Report of 3 Species of Angelsharks: Squatina aculeata, S. oculata and S. squatina. Report to National Marine Fisheries Service, Office of Protected Resources; 2016; pp. 6–74. Available online: repository.library.noaa.gov/view/noaa/16283 (accessed on 1 September 2020).
- Vaullegeard, A.C. Recherches sur les tétrarhynqués. Mém. Soc. Linn. Normandie 1899, 19, 187–376. [Google Scholar]
- Vaullegeard, A.C. Sur les Tétrarhynqués de la collection helminthologique du Professeur Corrado Parona, de Gênes. Atti. Soc. Ligust. Sci. Nat. Geogr. 1901, 12, 43–49. [Google Scholar]
- Pintner, T. Wenig Bekanntes und Unbekanntes von Rüssel bandwürmer. Sitzungsber Akademie d Wissenschaften Wien Math–Naturw Classe 1 1930, 139, 445–537. [Google Scholar]
- Pascual, S.; Arias, C.; Guerra, Á. First record of the trypanorhynchidean parasite Nybelinia lingualis Cuvier, 1817 in the squid Todaropsis eblanae (Cephalopoda: Ommastrephidae). Sci. Mar. 1996, 60, 553–555. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tedesco, P.; Caffara, M.; Gustinelli, A.; Fiorito, G.; Fioravanti, M.L. Metacestodes of Elasmobranch Tapeworms in Octopus vulgaris (Mollusca, Cephalopoda) from Central Mediterranean—SEM and Molecular Data. Animals 2020, 10, 2038. https://doi.org/10.3390/ani10112038
Tedesco P, Caffara M, Gustinelli A, Fiorito G, Fioravanti ML. Metacestodes of Elasmobranch Tapeworms in Octopus vulgaris (Mollusca, Cephalopoda) from Central Mediterranean—SEM and Molecular Data. Animals. 2020; 10(11):2038. https://doi.org/10.3390/ani10112038
Chicago/Turabian StyleTedesco, Perla, Monica Caffara, Andrea Gustinelli, Graziano Fiorito, and Maria Letizia Fioravanti. 2020. "Metacestodes of Elasmobranch Tapeworms in Octopus vulgaris (Mollusca, Cephalopoda) from Central Mediterranean—SEM and Molecular Data" Animals 10, no. 11: 2038. https://doi.org/10.3390/ani10112038
APA StyleTedesco, P., Caffara, M., Gustinelli, A., Fiorito, G., & Fioravanti, M. L. (2020). Metacestodes of Elasmobranch Tapeworms in Octopus vulgaris (Mollusca, Cephalopoda) from Central Mediterranean—SEM and Molecular Data. Animals, 10(11), 2038. https://doi.org/10.3390/ani10112038