GC-TOF-MS-Based Metabolomics Analyses of Liver and Intestinal Contents in the Overfed vs. Normally-Fed Geese


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiment
2.2. Treatment of Goose Primary Cells with Metabolites
2.3. Sample Preparation for Metabolomics Analysis
2.4. GC-TOF-MS Analysis
2.5. Real-Time PCR Analysis
2.6. Data Processing and Analyses
3. Results
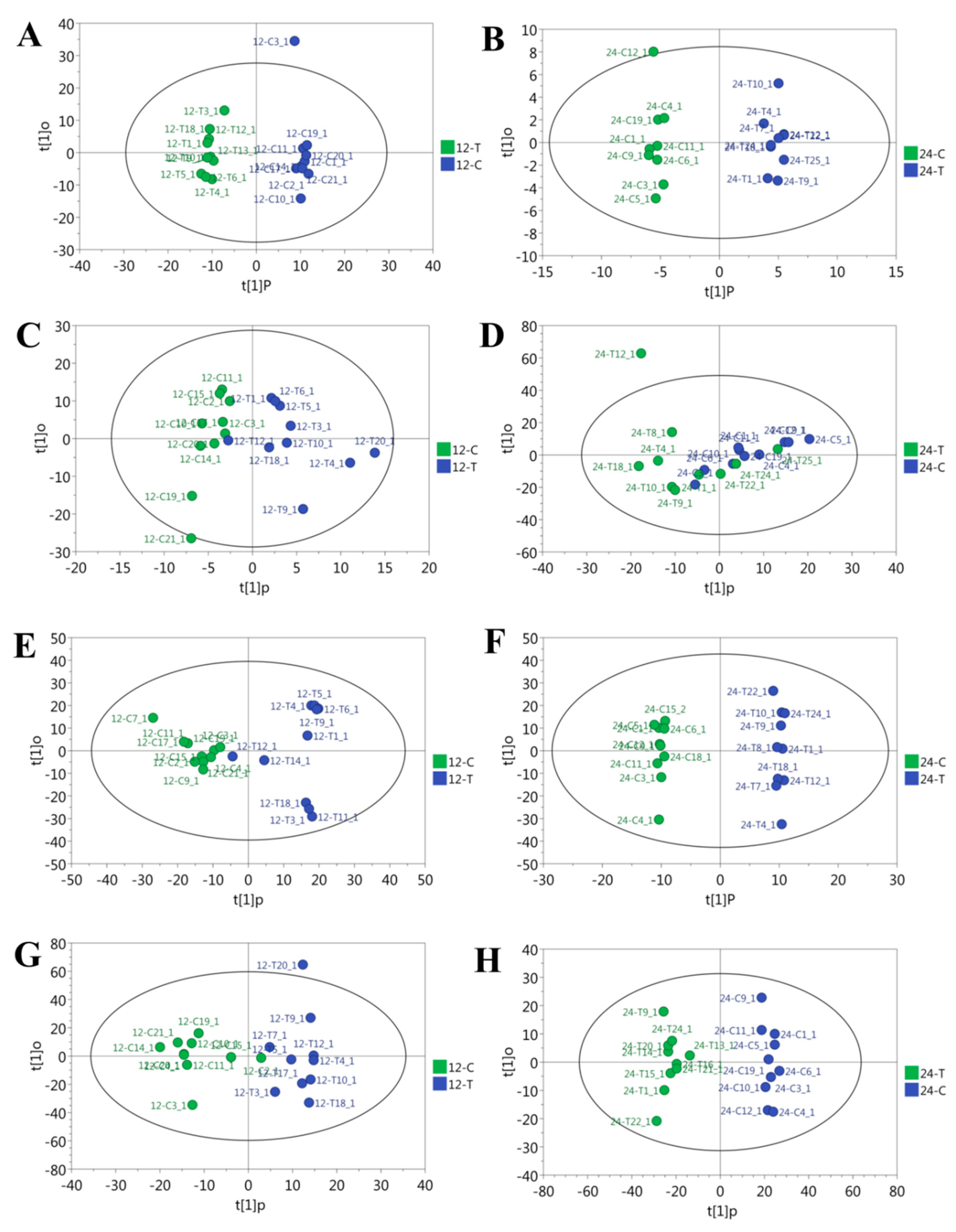
3.1. Overall Changes in Metabolite Profile Caused by Overfeeding
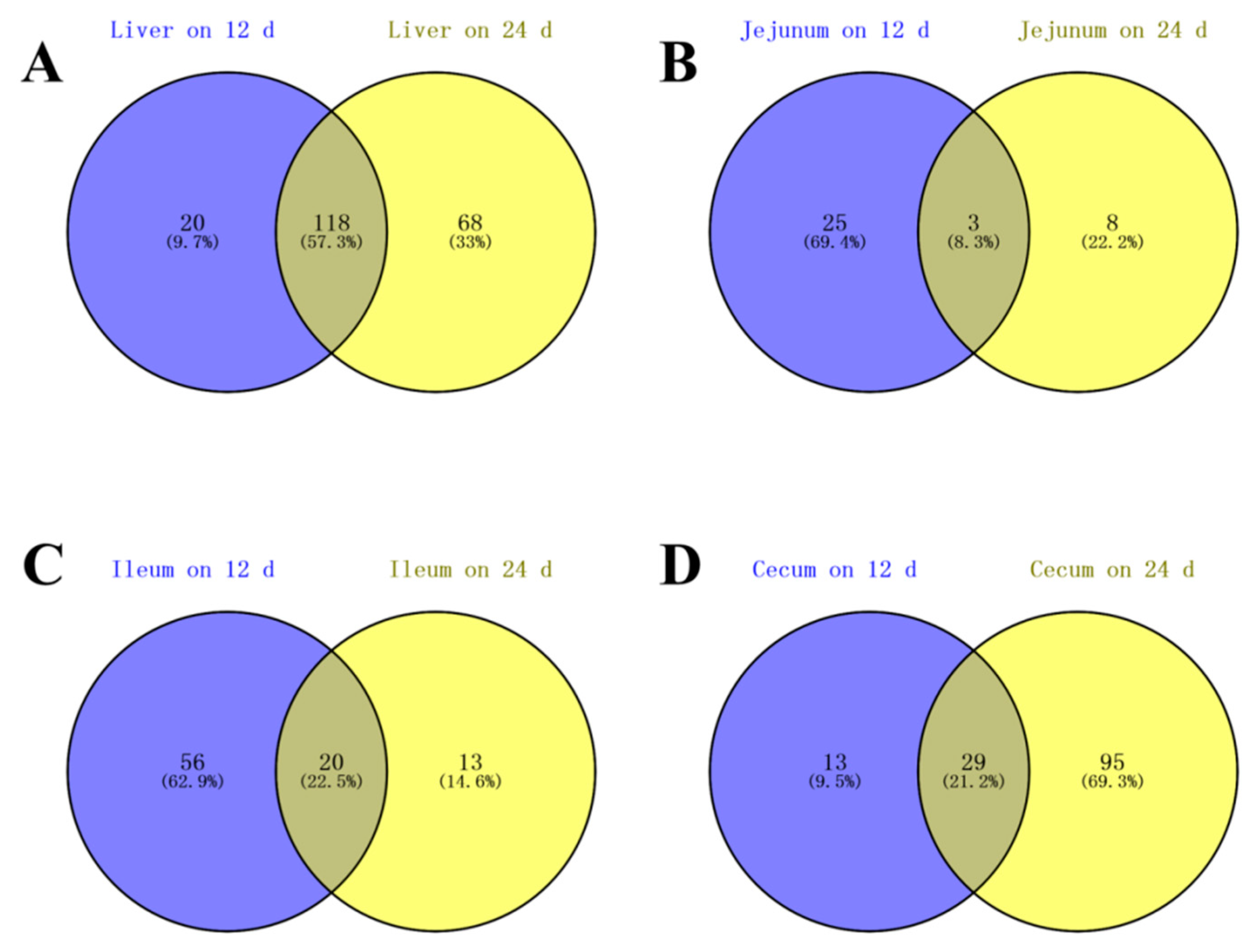
3.2. Identification of Differential Metabolites
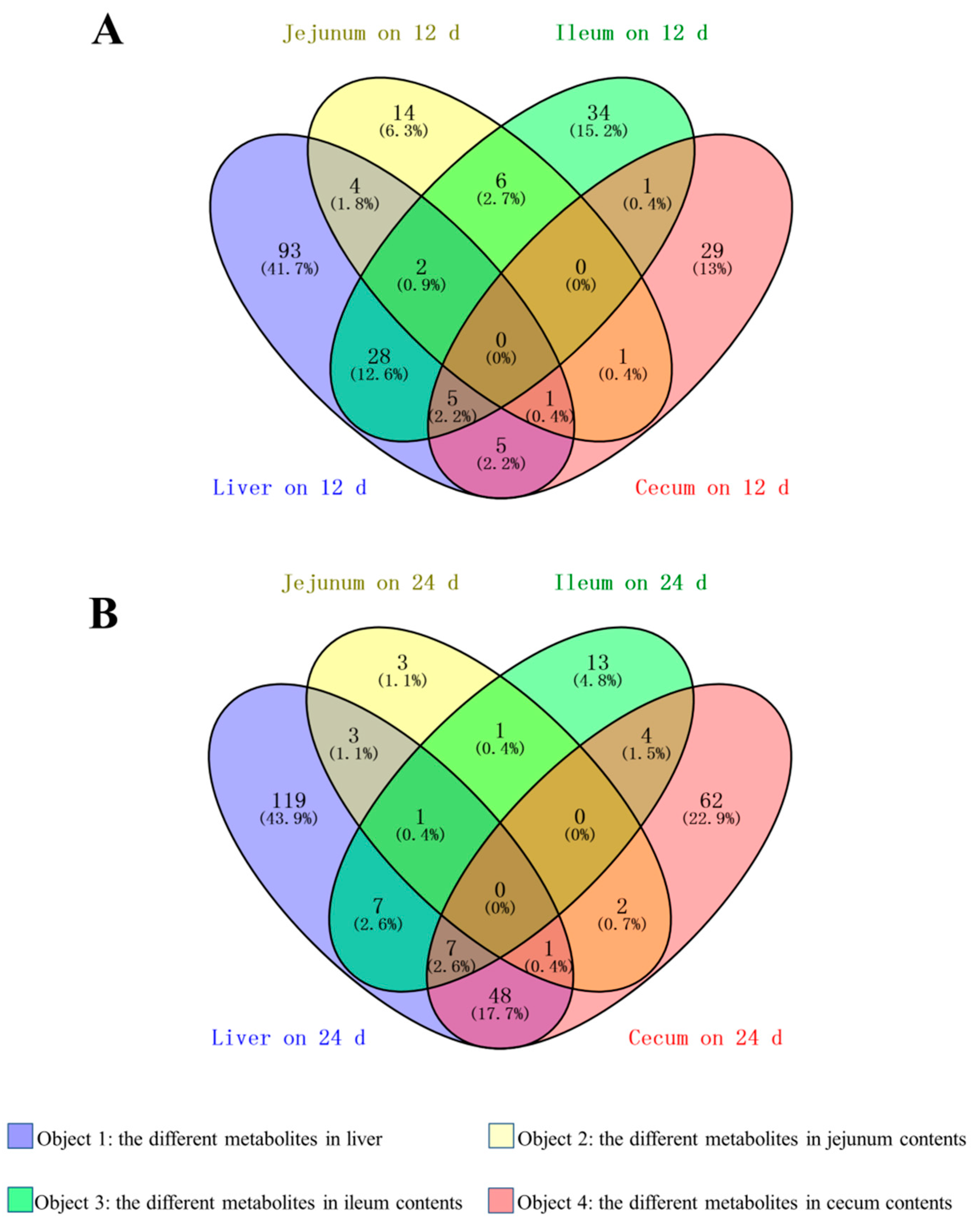
3.3. Identification of Common Differential Metabolites over Different Overfeeding Times
3.4. Identification of Common Differential Metabolites between the Liver and Intestinal Contents
3.5. The Metabolic Pathways in Which the Differential Metabolites Are Involved
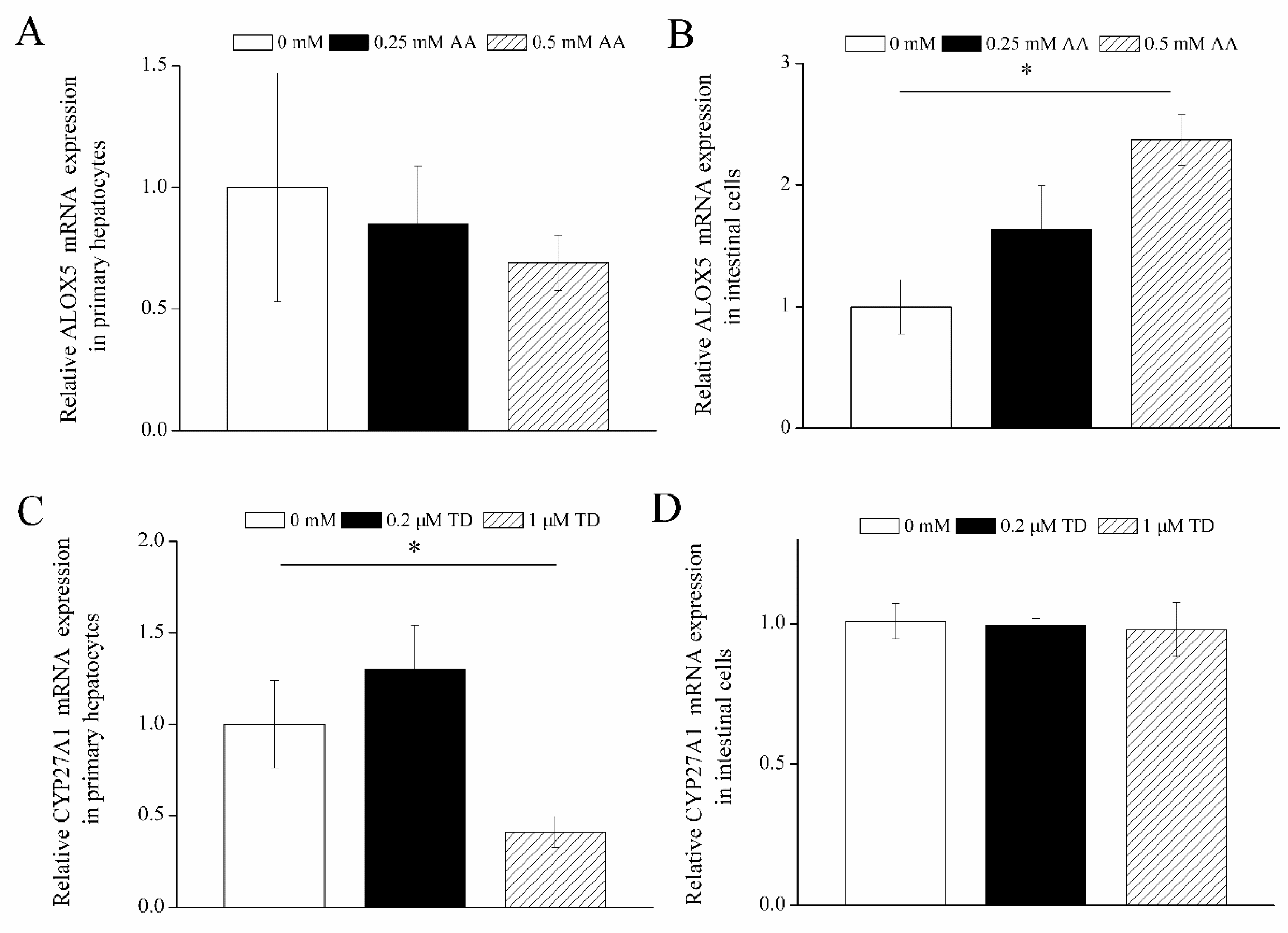
3.6. Effects of Arachidonic Acid and Tetrahydrocorticosterone on Gene Expression in Goose Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tilg, H.; Moschen, A.R. Evolution of inflammation in nonalcoholic fatty liver disease: The multiple parallel hits hypothesis. Hepatology 2010, 52, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Parola, M.; Vajro, P. Nocturnal hypoxia in obese-related obstructive sleep apnea as a putative trigger of oxidative stress in pediatric NAFLD progression. J. Hepatol. 2016, 65, 470–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wruck, W.; Graffmann, N.; Kawala, M.A.; Adjaye, J. Concise review: Current status and future directions on research related to nonalcoholic fatty liver disease. Stem Cells 2017, 35, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compare, D.; Coccoli, P.; Rocco, A.; Nardone, O.M.; Maria, S.D.; Cartenì, M.; Nardone, G. Gut–liver axis: The impact of gut microbiota on non alcoholic fatty liver disease. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 471–476. [Google Scholar] [CrossRef]
- Esteve, E.; Ricart, W.; Fernández-Real, J.M. Gut microbiota interactions with obesity, insulin resistance and type 2 diabetes: Did gut microbiote co-evolve with insulin resistance? Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 483–490. [Google Scholar] [CrossRef]
- Roy, T.L.; Llopis, M.; Lepage, P.; Bruneau, A.; Rabot, S.; Bevilacqua, C.; Martin, P.; Philippe, C.; Walker, F.; Bado, A.; et al. Intestinal microbiota determines development of non-alcoholic fatty liver disease in mice. Gut 2013, 62, 1787–1794. [Google Scholar] [CrossRef]
- Hermier, D.; Rousselot-Pailley, D.; Peresson, R.; Sellier, N. Influence of orotic acid and estrogen on hepatic lipid storage and secretion in the goose susceptible to liver steatosis. Biochim. Biophys. Acta 1994, 1211, 97–106. [Google Scholar] [CrossRef]
- Geng, T.Y.; Yang, B.; Li, F.Y.; Xia, L.L.; Wang, Q.Q.; Zhao, X.; Gong, D.Q. Identification of protective components that prevent the exacerbation of goose fatty liver: Characterization, expression and regulation of adiponectin receptors. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 194–195, 32–38. [Google Scholar] [CrossRef]
- Geng, T.Y.; Xia, L.L.; Li, F.Y.; Xia, J.; Zhang, Y.H.; Wang, Q.Q.; Yang, B.; Montgomery, S.; Cui, H.M.; Gong, D.Q. The role of endoplasmic reticulum stress and insulin resistance in the occurrence of goose fatty liver. Biochem. Biophys. Res. Commun. 2015, 465, 83–87. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, X.; Wang, Q.; Sun, X.X.; Xia, L.L.; Wang, Q.Q.; Yang, B.; Zhang, Y.H.; Montgomery, S.; Meng, H.; et al. Prosteatotic and protective components in a unique model of fatty liver: Gut microbiota and suppressed complement system. Sci. Rep. 2016, 6, 31763. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.M.; Liu, T.J.; Wang, Q.; Zhang, R.; Liu, L.; Gong, D.Q.; Geng, T.Y. Fatty acids modulate the expression of pyruvate kinase and arachidonate-lipoxygenase through PPARγ/CYP2C45 pathway: A link to goose fatty liver. Poult. Sci. 2019, 98, 4346–4358. [Google Scholar] [CrossRef]
- Zhang, J.M.; Wang, X.H.; Hao, L.H.; Wang, H.; Zhang, X.Y.; Muhammad, I.; Qi, Y.; Li, G.L.; Sun, X.Q. Nrf2 is crucial for the down-regulation of Cyp7a1 induced by arachidonic acid in Hepg2 cells. Environ. Toxicol. Pharmacol. 2017, 52, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Y.; Hao, R.J.; Du, X.D.; Wang, Q.H.; Deng, Y.W.; Sun, R.J.; Zheng, Z. GC–TOF/MS-based metabolomics studies on the effect of protein sources in formulated diet for pearl oyster Pinctada fucata martensii. Aquaculture 2017, 486, 139–147. [Google Scholar] [CrossRef]
- Wen, B.; Jin, S.R.; Chen, Z.Z.; Gao, J.Z. Physiological responses to cold stress in the gills of discus fish (Symphysodon aequifasciatus) revealed by conventional biochemical assays and GC-TOF-MS metabolomics. Sci. Total. Environ. 2018, 640–641, 1372–1381. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Pagadala, M.; Kasumov, T.; McCullough, A.J.; Zein, N.N.; Kirwan, J.P. Role of ceramides in nonalcoholic fatty liver disease. Trends Endocrinol. Metab. 2012, 23, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Nikolova-Karakashian, M. Sphingolipids at the crossroads of NAFLD and senescence. Adv. Cancer Res. 2018, 140, 155–190. [Google Scholar] [CrossRef]
- Zhao, M.M.; Liu, L.; Gong, D.Q.; Geng, T.Y. The research on the expression of hexokinase-1 in the development of goose fatty liver and the regulation of its function. in preparation.
- Carneiro, E.M.; Latorraca, M.Q.; Araujo, E.; Beltrá, M.; Oliveras, M.J.; Navarro, M.; Berná, G.; Bedoya, F.J.; Velloso, L.A.; Soria, B.; et al. Taurine supplementation modulates glucose homeostasis and islet function. J. Nutr. Biochem. 2009, 20, 503–511. [Google Scholar] [CrossRef]
- Vessey, D.A.; Whitney, J.; Gollan, J.L. The role of conjugation reactions in enhancing biliary secretion of bile acids. Biochem. J. 1983, 214, 923–927. [Google Scholar] [CrossRef] [Green Version]
- Gu, M.; Bai, N.; Kortner, T.M. Taurocholate supplementation attenuates the changes in growth performance, feed utilization, lipid digestion, liver abnormality and sterol metabolism in turbot (Scophthalmus maximus) fed high level of plant protein. Aquaculture 2017, 468, 597–604. [Google Scholar] [CrossRef] [Green Version]
- Chaurasia, B.; Summers, S.A. Ceramides-lipotoxic inducers of metabolic disorders. Trends Endocrinol. Metab. 2015, 26, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Walsh, M.T.; Hammad, S.M.; Hussain, M.M. Sphingolipids and lipoproteins in health and metabolic disorders. Trends Endocrinol. Metab. 2017, 28, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.P.; Keller, J.N. Regulation of energy metabolism by inflammation: A feedback response in obesity and calorie restriction. Aging (Albany NY) 2010, 2, 361–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divella, R.; Mazzocca, A.; Daniele, A.; Sabbà, C.; Paradiso, A. Obesity, nonalcoholic fatty liver disease and adipocytokines network in promotion of cancer. Int. J. Biol. Sci. 2019, 15, 610–616. [Google Scholar] [CrossRef] [Green Version]
- Wree, A.; Broderick, L.; Canbay, A.; Hoffman, H.M.; Feldstein, A.E. From NAFLD to NASH to cirrhosis-new insights into disease mechanisms. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 627–636. [Google Scholar] [CrossRef]
- Montefusco, D.J.; Allegood, J.C.; Spiegel, S.; Cowart, L.A. Non-alcoholic fatty liver disease: Insights from sphingolipidomics. Biochem. Biophys. Res. Commun. 2018, 504, 608–616. [Google Scholar] [CrossRef]
- Allard, J.P.; Aghdassi, E.; Mohammed, S.; Raman, M.; Avand, G.; Arendt, B.M.; Jalali, P.; Kandasamy, T.; Prayitno, N.; Sherman, M.; et al. Nutritional assessment and hepatic fatty acid composition in non-alcoholic fatty liver disease(NAFLD): A cross-sectional study. J. Hepatol. 2008, 48, 300–307. [Google Scholar] [CrossRef]
- Sánchez-Valle, V.; Chávez-Tapia, N.C.; Uribe, M.; Méndez-Sánchez, N. Role of oxidative stress and molecular changes in liver fibrosis: A review. Curr. Med. Chem. 2012, 19, 4850–4860. [Google Scholar] [CrossRef]
- Sztolsztener, K.; Chabowski, A.; Harasim-Symbor, E.; Bielawiec, P.; Konstantynowicz-Nowicka, K. Arachidonic acid as an early indicator of inflammation during non-alcoholic fatty liver disease development. Biomolecules 2020, 10, 1133. [Google Scholar] [CrossRef]
- Tarantino, G.; Finelli, C. Pathogenesis of hepatic steatosis: The link between hypercortisolism and non-alcoholic fatty liver disease. World J. Gastroenterol. 2013, 19, 6735–6743. [Google Scholar] [CrossRef]
- Starick, S.R.; Ibn-Salem, J.; Jurk, M.; Hernandez, C.; Love, M.I.; Chung, H.R.; Vingron, M.; Thomas-Chollier, M.; Meijsing, S.H. ChIP-exo signal associated with DNA-binding motifs provides insight into the genomic binding of the glucocorticoid receptor and cooperating transcription factors. Genome Res. 2015, 25, 825–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.W.; Uhlenhaut, N.H.; Rauch, A.; Weiner, J.; Hübner, S.; Hübner, N.; Won, K.J.; Lazar, M.A.; Tuckermann, J.; Steger, D.J. Genomic redistribution of GR monomers and dimers mediates transcriptional response to exogenous glucocorticoid in vivo. Genome Res. 2015, 25, 836–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenson, M.; Kilroy, G.; York, D.A.; Braymer, D. Abnormal regulation of hepatic glucocorticoid receptor mRNA and receptor protein distribution in the obese zucker rat. Obes. Res. 1996, 4, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.M.; Kornfeld, J.W.; Friedbichler, K.; Blaas, L.; Egger, G.; Esterbauer, H.; Hasselblatt, P.; Schlederer, M.; Haindl, S.; Wagner, K.U.; et al. Impairment of hepatic growth hormone and glucocorticoid receptor signaling causes steatosis and hepatocellular carcinoma in mice. Hepatology 2011, 54, 1398–1409. [Google Scholar] [CrossRef]
- Gueguen, Y.; Ferrari, L.; Souidi, M.; Batt, A.M.; Lutton, C.; Siest, G.; Visvikis, S. Compared effect of immunosuppressive drugs cyclosporine A and rapamycin on cholesterol homeostasis key enzymes CYP27A1 and HMG-CoA reductase. Basic Clin. Pharmacol. Toxicol. 2007, 100, 392–397. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name a | GenBank Number | Primer Sequence (5′ to 3′) | Product Size (bp) |
|---|---|---|---|
| ALOX5 | XM_013201170.1 | F: CAGGGAAAGCTGGAAAACAG | 240 |
| R: AGCTTTGCTCTTCCATCTCG | |||
| CYP27A1 | XM_013186353.1 | F: GACCCAGCACTTCATCGACT | 226 |
| R: CAGTGTGTTGGAGGTCGTGT | |||
| GAPDH | XM_013199522.1 | F:GCCATCAATGATCCCTTCAT | 155 |
| R:CTGGGGTCACGCTCCTG |
| Item a | Liver | Jejunum | Ileum | Cecum | ||||
|---|---|---|---|---|---|---|---|---|
| 12 d | 24 d | 12 d | 24 d | 12 d | 24 d | 12 d | 24 d | |
| Increased | 8 | 3 | 27 | 4 | 28 | 5 | 6 | 42 |
| Decreased | 130 | 183 | 1 | 7 | 48 | 28 | 36 | 82 |
| Total | 138 | 186 | 28 | 11 | 76 | 33 | 42 | 124 |
| Different Metabolites | Related Metabolic Pathway |
|---|---|
| Liver | |
| 3-Phosphoglycerate | Glycolysis/Gluconeogenesis; Glycine, serine, and threonine metabolism; Glycerolipid metabolism; Biosynthesis of terpenoids and steroids; Glucagon signaling pathway; Pentose phosphate pathway |
| Glutaric acid | Fatty acid degradation |
| 3α,7α,12α-Trihydroxycoprostane | Primary bile acid biosynthesis |
| Sphingosine | Sphingolipid metabolism; Sphingolipid signaling pathway; Apoptosis |
| Lignoceric acid | Biosynthesis of unsaturated fatty acids |
| Squalene | Biosynthesis of terpenoids and steroids |
| Glutathione | ATP-binding cassette transporters; Bile secretion |
| Jejunum | |
| Myo-inositol | ATP-binding cassette transporters; Galactose metabolism; Ascorbate and aldarate metabolism; Inositol phosphate metabolism; Phosphatidylinositol signaling system |
| Raffinose | ATP-binding cassette transporters; Galactose metabolism |
| 24, 25-Dihydrolanosterol | Steroid biosynthesis |
| O-Phosphorylethanolamine | Glycerophospholipid metabolism; Sphingolipid signaling pathway; Sphingolipid metabolism |
| Ileum | |
| 3α,7α,12α-Trihydroxycoprostane | Primary bile acid biosynthesis |
| Taurine | ATP-binding cassette transporters; Taurine and hypotaurine metabolism; Sulfur metabolism |
| Arbutin | Glycolysis/Gluconeogenesis; Phosphotransferase system |
| Salicin | Glycolysis/Gluconeogenesis; Phosphotransferase system |
| Ethanolamine | Glycerophospholipid metabolism |
| Sucrose | Phosphotransferase system; Galactose metabolism; ATP-binding cassette transporters |
| Creatine | Glycine, serine, and threonine metabolism |
| Raffinose | ATP-binding cassette transporters; Galactose metabolism |
| Maltotriose | ATP-binding cassette transporters |
| 2, 6-Diaminopimelic acid | Biosynthesis of amino acids |
| 24, 25-Dihydrolanosterol | Steroid biosynthesis |
| Arachidonic acid (ARA) | Eicosanoids; gonadotropin-releasing hormone signaling pathway; Aldosterone synthesis and secretion; Linoleic acid metabolism; Biosynthesis of unsaturated fatty acids; Fc gamma R-mediated phagocytosis; Inflammatory mediator regulation of transient receptor potential channels; ARA metabolism; Regulation of lipolysis in adipocytes |
| Cecum | |
| Adipic acid | Degradation of aromatic compounds; Caprolactam degradation |
| Pipecolinic acid | Biosynthesis of alkaloids derived from ornithine, lysine, and nicotinic acid |
| inosine | Purine metabolism |
| Phenylacetic acid | Phenylalanine metabolism |
| Different Metabolites | Related Metabolic Pathway |
|---|---|
| Liver | |
| Nicotinamide | Nicotinate and nicotinamide metabolism |
| Ileum | |
| Glycine | Primary bile acid biosynthesis |
| Taurine | Primary bile acid biosynthesis; Taurine and hypotaurine metabolism; Sulfur metabolism; ATP-binding cassette transporters |
| 24, 25-Dihydrolanosterol | Steroid biosynthesis |
| Zymosterol | Steroid biosynthesis |
| Ethanolamine | Phosphote and phosphite metabolism; Glycerophospholipid metabolism |
| Palmitic acid | Biosynthesis of unsaturated fatty acids; Fatty acid biosynthesis; Fatty acid metabolism; Fatty acid degradation; Fatty acid elongation |
| Cecum | |
| L-Malic acid | Biosynthesis of alkaloids derived from ornithine, lysine, and nicotinic acid; Glucagon signaling pathway; Biosynthesis of alkaloids derived from histidine and purine; Biosynthesis of alkaloids derived from shikimate pathway; Biosynthesis of terpenoids and steroids; Citrate cycle (TCA cycle); Pyruvate metabolism; Biosynthesis of phenylpropanoids; Biosynthesis of alkaloids derived from terpenoid and polyketide; Glyoxylate and dicarboxylate metabolism |
| Pipecolinic acid | Biosynthesis of alkaloids derived from ornithine, lysine, and nicotinic acid |
| Fumaric acid | Biosynthesis of alkaloids derived from ornithine, lysine, and nicotinic acid; Glucagon signaling pathway; Biosynthesis of alkaloids derived from histidine and purine; Biosynthesis of alkaloids derived from shikimate pathway; Biosynthesis of terpenoids and steroids; TCA cycle; Pyruvate metabolism; Biosynthesis of phenylpropanoids; Biosynthesis of alkaloids derived from terpenoid and polyketide; Phenylalanine metabolism; Nicotite and nicotimide metabolism; Arginine biosynthesis; Oxidative phosphorylation; Tyrosine metabolism; Alanine, aspartate and glutamate metabolism; Styrene degradation |
| Cholesterol | Biosynthesis of secondary metabolites; Primary bile acid biosynthesis; Steroid hormone biosynthesis; Biosynthesis of alkaloids derived from terpenoid and polyketide; Steroid degradation; Steroid biosynthesis; Vitamin digestion and absorption; Bile secretion; Fat digestion and absorption |
| Adipic acid | Caprolactam degradation; Degradation of aromatic compounds |
| Malonic acid | Pyrimidine metabolism; beta-alanine metabolism |
| Uracil | Pyrimidine metabolism; beta-alanine metabolism; Pantothete and CoA biosynthesis |
| Myo-inositol | ATP-binding cassette transporters; Galactose metabolism; Phosphatidylinositol signaling system; Inositol phosphate metabolism; Ascorbate and aldarate metabolism |
| 2-Amino-3-hydroxybutyric acid | ATP-binding cassette transporters; Glycine, serine, and threonine metabolism |
| Glycine | Primary bile acid biosynthesis |
| Creatine | Glycine, serine and threonine metabolism; Arginine and proline metabolism |
| Ethanolamine | Glycerophospholipid metabolism; Phosphote and phosphite metabolism |
| O-Phosphorylethanolamine | Glycerophospholipid metabolism; Sphingolipid metabolism; Sphingolipid signaling pathway |
| Behenic acid | Biosynthesis of unsaturated fatty acids |
| ARA | Biosynthesis of unsaturated fatty acids; ARA metabolism; Eicosanoids; Linoleic acid metabolism; Aldosterone synthesis and secretion; Regulation of lipolysis in adipocytes |
| Lignoceric acid | Biosynthesis of unsaturated fatty acids |
| Tetrahydrocorticosterone | Steroid hormone biosynthesis |
| Inosine | Purine metabolism |
| 24, 25-Dihydrolanosterol | Steroid biosynthesis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, M.; Xing, Y.; Liu, L.; Fan, X.; Liu, L.; Geng, T.; Gong, D. GC-TOF-MS-Based Metabolomics Analyses of Liver and Intestinal Contents in the Overfed vs. Normally-Fed Geese. Animals 2020, 10, 2375. https://doi.org/10.3390/ani10122375
Zhao M, Xing Y, Liu L, Fan X, Liu L, Geng T, Gong D. GC-TOF-MS-Based Metabolomics Analyses of Liver and Intestinal Contents in the Overfed vs. Normally-Fed Geese. Animals. 2020; 10(12):2375. https://doi.org/10.3390/ani10122375
Chicago/Turabian StyleZhao, Minmeng, Ya Xing, Lidong Liu, Xiang Fan, Long Liu, Tuoyu Geng, and Daoqing Gong. 2020. "GC-TOF-MS-Based Metabolomics Analyses of Liver and Intestinal Contents in the Overfed vs. Normally-Fed Geese" Animals 10, no. 12: 2375. https://doi.org/10.3390/ani10122375
APA StyleZhao, M., Xing, Y., Liu, L., Fan, X., Liu, L., Geng, T., & Gong, D. (2020). GC-TOF-MS-Based Metabolomics Analyses of Liver and Intestinal Contents in the Overfed vs. Normally-Fed Geese. Animals, 10(12), 2375. https://doi.org/10.3390/ani10122375