Pathology of Coronavirus Infections: A Review of Lesions in Animals in the One-Health Perspective

 , , , , , ,
, , , , , ,  , , ,
, , , 
 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
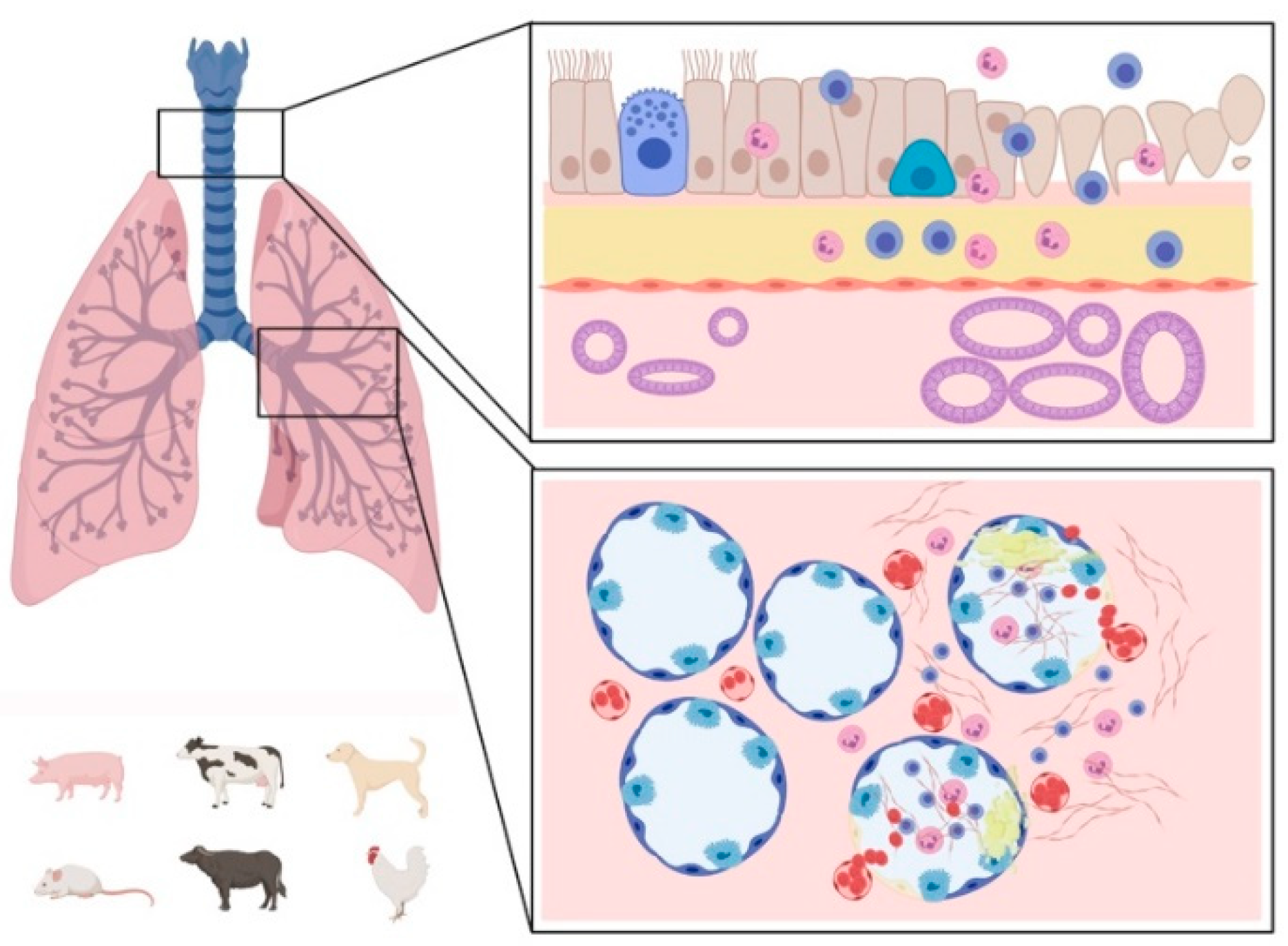
2. Pathogenesis
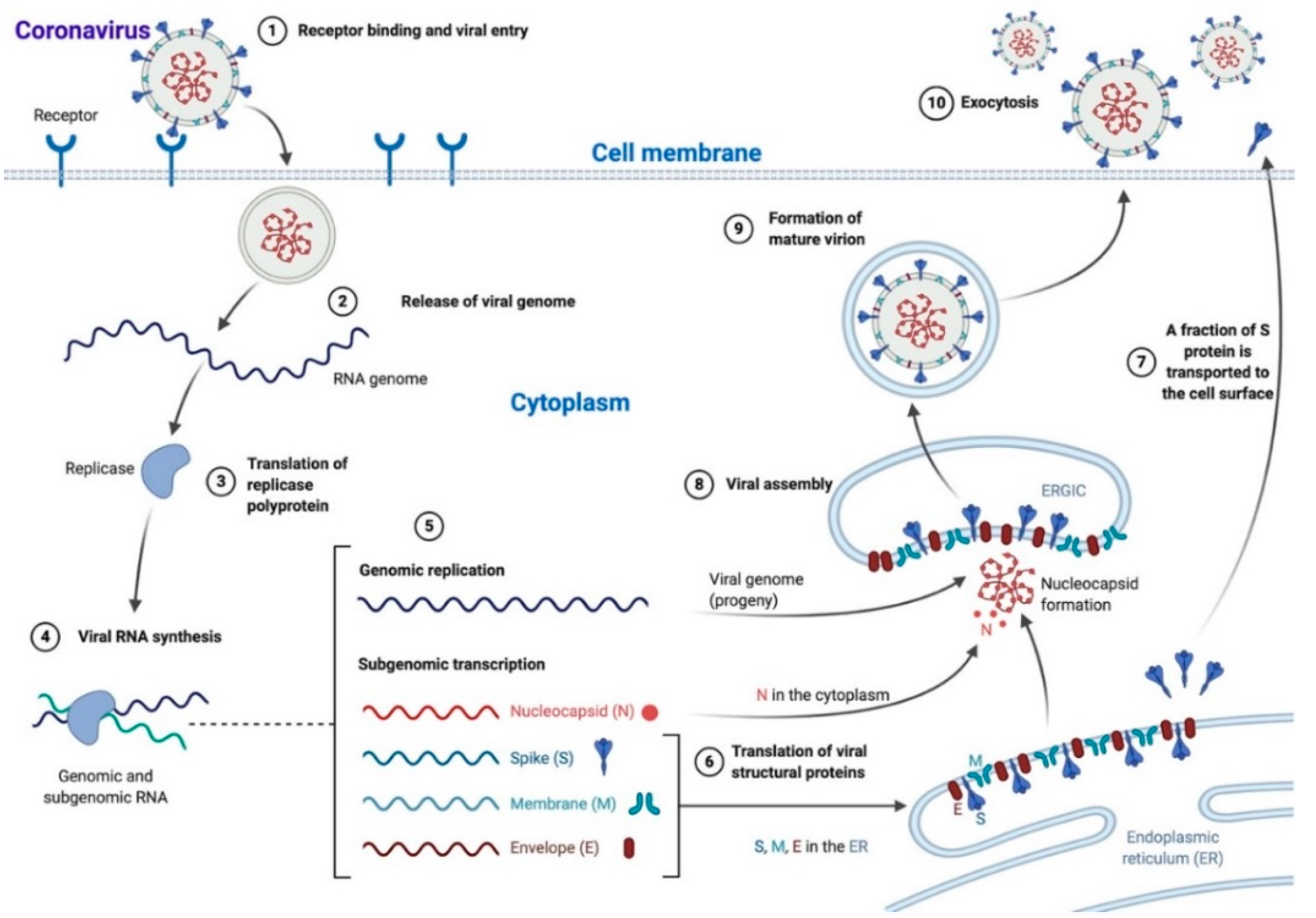
2.1. Viral Life Cycle
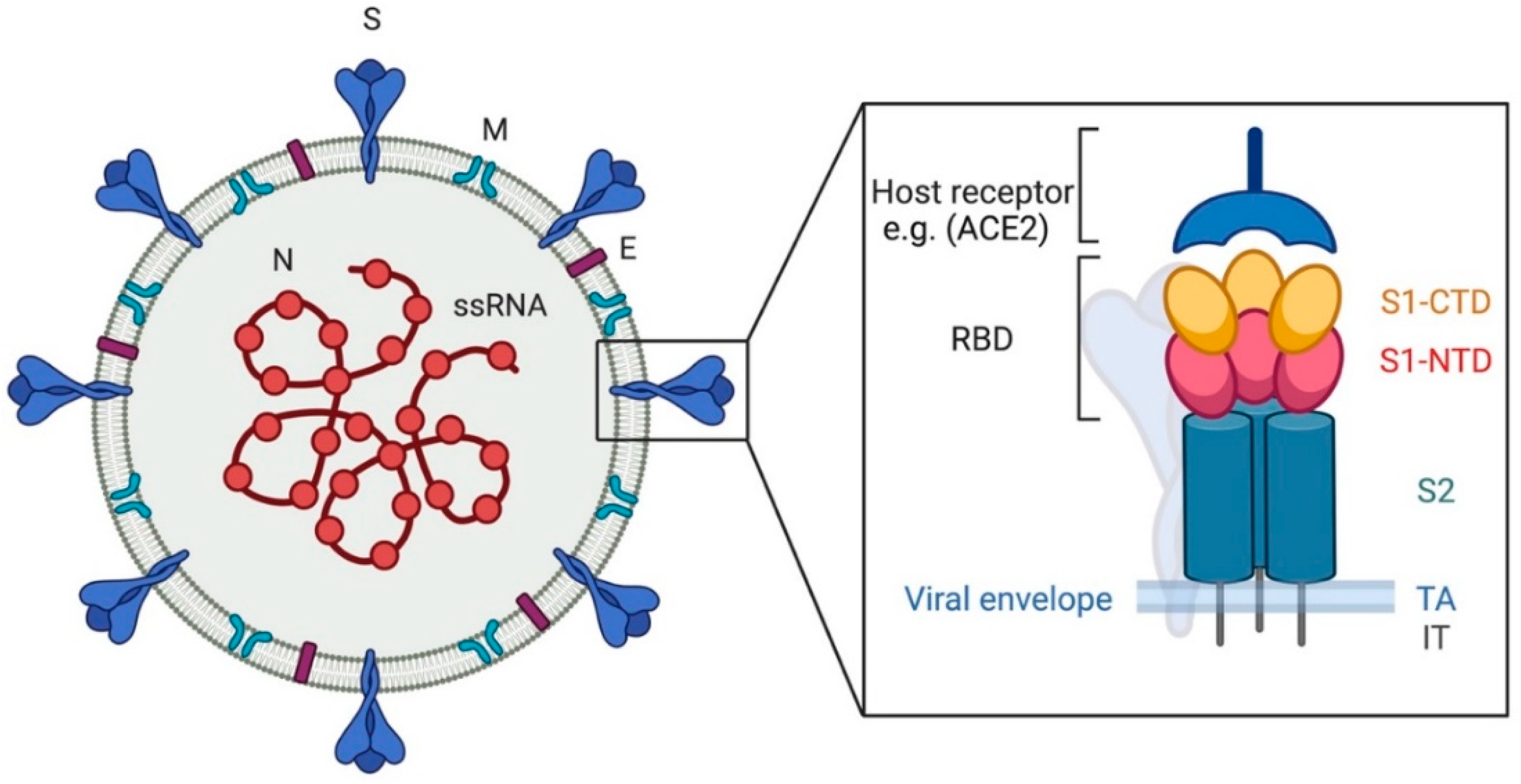
2.2. Viral Spike Protein
2.3. Viral Mutation and Recombination
2.4. Mutation
2.5. Recombination
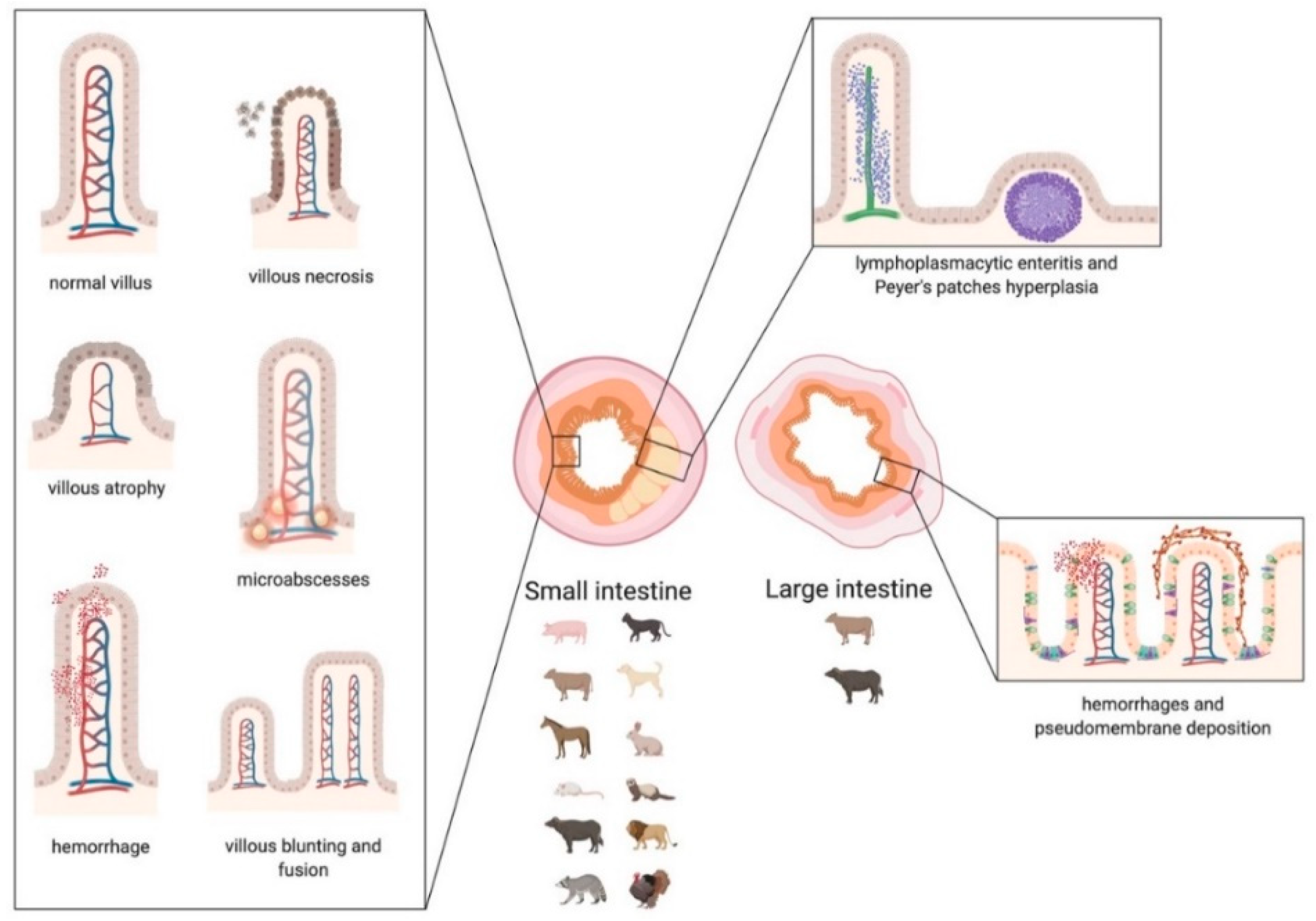
3. Coronavirus Diseases in Animals
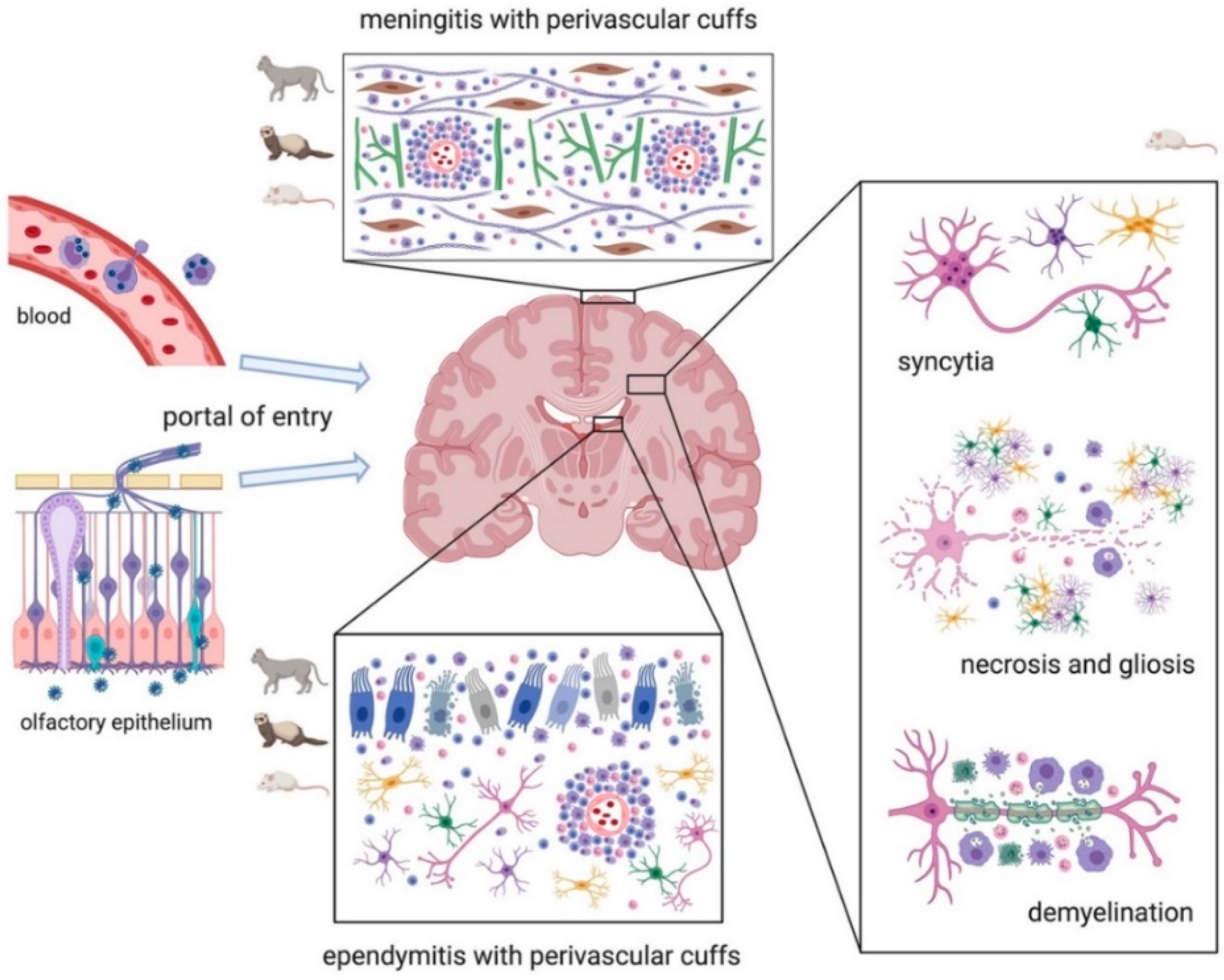
3.1. CoVs in Pets (Dogs and Cats)
3.2. CoVs in Ferrets
3.3. CoVs in Bovines
3.4. CoVs in Sheep and Goats
3.5. CoVs in Equids
3.6. CoVs in Swine
3.7. CoVs in Wild Animals
3.8. Wild Felids
3.9. Wild Ruminants
3.10. Other Wild Animals
3.11. CoVs in Non-Human Primates
3.12. CoVs in Marine Mammals
3.13. CoVs in Laboratory Animals
3.14. CoVs in Birds
3.15. Domestic Birds
3.16. Wild Birds
3.17. CoVs in Fish
3.18. CoVs in Reptiles and Amphibians
3.19. CoVs in Humans
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Wu, T.; Liu, Q.; Yang, Z. The SARS-CoV-2 outbreak: What we know. Int. J. Infect. Dis. 2020, 94, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.A.; Daszak, P.; Wood, J.L.N. One Health, emerging infectious diseases and wildlife: Two decades of progress? Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreuder Johnson, C.; Hitchens, P.L.; Smiley Evans, T.; Goldstein, T.; Thomas, K.; Clements, A.; Joly, D.O.; Wolfe, N.D.; Daszak, P.; Karesh, W.B.; et al. Spillover and pandemic properties of zoonotic viruses with high host plasticity. Sci. Rep. 2015, 5, 14830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sleeman, J.M.; DeLiberto, T.; Nguyen, N. Optimization of human, animal, and environmental health by using the One Health approach. J. Vet. Sci. 2017, 18, 263. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 2 December 2020).
- Shi, Z.L.; Guo, D.; Rottier, P.J.M. Coronavirus: Epidemiology, genome replication and the interactions with their hosts. Virol. Sin. 2016, 31, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; He, W.; Wang, L.; Lai, A.; Ji, X.; Zhai, X.; Li, G.; Suchard, M.A.; Tian, J.; Zhou, J.; et al. COVID-19: Epidemiology, Evolution, and Cross-Disciplinary Perspectives. Trends Mol. Med. 2020, 26, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef]
- Li, F. Structure, Function, and Evolution of Coronavirus Spike Proteins. Annu. Rev. Virol. 2016, 3, 237–261. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.D.; Berry, D.M.; Cunningham, C.H.; Hamre, D.; Hofstad, M.S.; Mallucci, L.; McIntosh, K.; Tyrrell, D.A.J. Virology: Coronaviruses. Nature 1968, 220, 650. [Google Scholar]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Committee on Taxonomy of Viruses ICTV. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 1 October 2020).
- Luk, H.K.H.; Li, X.; Fung, J.; Lau, S.K.P.; Woo, P.C.Y. Molecular epidemiology, evolution and phylogeny of SARS coronavirus. Infect. Genet. Evol. 2019, 71, 21–30. [Google Scholar] [CrossRef] [PubMed]
- De Wit, E.; Van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Geng, M.; Peng, Y.; Meng, L.; Lu, S. Molecular immune pathogenesis and diagnosis of COVID-19. J. Pharm. Anal. 2020, 10, 102–108. [Google Scholar] [CrossRef]
- Lam, T.T.-Y.; Jia, N.; Zhang, Y.-W.; Shum, M.H.-H.; Jiang, J.-F.; Zhu, H.-C.; Tong, Y.-G.; Shi, Y.-X.; Ni, X.-B.; Liao, Y.; et al. Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 2020, 583, 282–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Geng, X.; Tan, Y.; Li, Q.; Xu, C.; Xu, J.; Hao, L.; Zeng, Z.; Luo, X.; Liu, F.; et al. New understanding of the damage of SARS-CoV-2 infection outside the respiratory system. Biomed. Pharmacother. 2020, 127, 110195. [Google Scholar] [CrossRef]
- Weiss, S.R.; Leibowitz, J.L. Coronavirus Pathogenesis. In Advances in Virus Research; Elsevier Inc.: Amsterdam, The Netherlands, 2011; pp. 85–164. [Google Scholar]
- Masters, P.S. The Molecular Biology of Coronaviruses. Adv. Virus Res. 2006, 65, 193–292. [Google Scholar]
- Fehr, A.R.; Perlman, S. Coronaviruses: An Overview of Their Replication and Pathogenesis. In Coronaviruses; Maier, H., Bickerton, E., Britton, P., Eds.; Humana Press: New York, NY, USA, 2015; Volume 1282, pp. 1–23. [Google Scholar]
- Weiss, S.R.; Navas-Martin, S. Coronavirus Pathogenesis and the Emerging Pathogen Severe Acute Respiratory Syndrome Coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef] [Green Version]
- Hulswit, R.J.G.; de Haan, C.A.M.; Bosch, B.-J. Coronavirus spike protein and tropism changes. In Advances in Virus Research; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 29–57. [Google Scholar]
- Tortorici, M.A.; Veesler, D. Structural insights into coronavirus entry. In Advances in Virus Research; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 93–116. [Google Scholar]
- Wong, L.Y.R.; Lui, P.Y.; Jin, D.Y. A molecular arms race between host innate antiviral response and emerging human coronaviruses. Virol. Sin. 2016, 31, 12–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decaro, N.; Lorusso, A. Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses. Vet. Microbiol. 2020, 244, 108693. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, N.C. An update on feline infectious peritonitis: Virology and immunopathogenesis. Vet. J. 2014, 201, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekes, G.; Thiel, H.J. Feline Coronaviruses: Pathogenesis of Feline Infectious Peritonitis, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 96. [Google Scholar]
- Vijaykrishna, D.; Smith, G.J.D.; Zhang, J.X.; Peiris, J.S.M.; Chen, H.; Guan, Y. Evolutionary Insights into the Ecology of Coronaviruses. J. Virol. 2007, 81, 4012–4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827.e19. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. An update on canine coronaviruses: Viral evolution and pathobiology. Vet. Microbiol. 2008, 132, 221–234. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. Canine Coronavirus: Not Only an Enteric Pathogen. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1121–1132. [Google Scholar] [CrossRef]
- Erles, K.; Toomey, C.; Brooks, H.W.; Brownlie, J. Detection of a group 2 coronavirus in dogs with canine infectious respiratory disease. Virology 2003, 310, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Decaro, N.; Mari, V.; von Reitzenstein, M.; Lucente, M.S.; Cirone, F.; Elia, G.; Martella, V.; King, V.L.; Di Bello, A.; Varello, K.; et al. A pantropic canine coronavirus genetically related to the prototype isolate CB/05. Vet. Microbiol. 2012, 159, 239–244. [Google Scholar] [CrossRef]
- Buonavoglia, C.; Decaro, N.; Martella, V.; Elia, G.; Campolo, M.; Desario, C.; Castagnaro, M.; Tempesta, M. Canine Coronavirus Highly Pathogenic for Dogs. Emerg. Infect. Dis. 2006, 12, 492–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipar, A.; Meli, M.L. Feline Infectious Peritonitis: Still an Enigma? Vet. Pathol. 2014, 51, 505–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drechsler, Y.; Alcaraz, A.; Bossong, F.J.; Collisson, E.W.; Diniz, P.P.V.P. Feline Coronavirus in Multicat Environments. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1133–1169. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; May, H.; Menger, S.; Weber, M.; Leukert, W.; Reinacher, M. Morphologic features and development of granulomatous vasculitis in feline infectious peritonitis. Vet. Pathol. 2005, 42, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Sigurardóttir, Ó.G.; Kolbjornsen, O.; Lutz, H. Orchitis in a cat associated with coronavirus infection. J. Comp. Pathol. 2001, 124, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Rota, A.; Paltrinieri, S.; Jussich, S.; Ubertalli, G.; Appino, S. Priapism in a castrated cat associated with feline infectious peritonitis. J. Feline Med. Surg. 2008, 10, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Osumi, T.; Mitsui, I.; Leutenegger, C.M.; Okabe, R.; Ide, K.; Nishifuji, K. First identification of a single amino acid change in the spike protein region of feline coronavirus detected from a coronavirus-associated cutaneous nodule in a cat. J. Feline Med. Surg. Open Rep. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Ernandes, M.A.; Cantoni, A.M.; Armando, F.; Corradi, A.; Ressel, L.; Tamborini, A. Feline coronavirus-associated myocarditis in a domestic longhair cat. J. Feline Med. Surg. Open Rep. 2019, 5. [Google Scholar] [CrossRef]
- Foley, J.E.; Leutenegger, C. A Review of Coronavirus Infection in the Central Nervous System of Cats and Mice. J. Vet. Intern. Med. 2001, 15, 438–444. [Google Scholar] [CrossRef]
- Zappulli, V.; Caliari, D.; Cavicchioli, L.; Tinelli, A.; Castagnaro, M. Systemic fatal type II coronavirus infection in a dog: Pathological findings and immunohistochemistry. Res. Vet. Sci. 2008, 84, 278–282. [Google Scholar] [CrossRef]
- Evermann, J.F.; Abbott, J.R.; Han, S. Canine coronavirus-associated puppy mortality without evidence of concurrent canine parvovirus infection. J. Vet. Diagn. Investig. 2005, 17, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Erles, K.; Brownlie, J. Investigation into the causes of canine infectious respiratory disease: Antibody responses to canine respiratory coronavirus and canine herpesvirus in two kennelled dog populations. Arch. Virol. 2005, 150, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Erles, K.; Brownlie, J. Canine Respiratory Coronavirus: An Emerging Pathogen in the Canine Infectious Respiratory Disease Complex. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Brooks, H.W.; Szladovits, B.; Erles, K.; Gibbons, R.; Shields, S.; Brownlie, J. Tropism and pathological findings associated with canine respiratory coronavirus (CRCoV). Vet. Microbiol. 2013, 162, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Kipar, A.; Köhler, K.; Leukert, W.; Reinacher, M. A comparison of lymphatic tissues from cats with spontaneous feline infectious peritonitis (FIP), cats with FIP virus infection but no FIP, and cats with no infection. J. Comp. Pathol. 2001, 125, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, J.D. Coronavirus infection in cats. Vet. Clin. N. Am. Small Anim. Pract. 1993, 23, 1–16. [Google Scholar] [CrossRef]
- Kipar, A.; Kremendahl, J.; Addie, D.D.; Leukert, W.; Grant, C.K.; Reinacher, M. Fatal enteritis associated with coronavirus infection in cats. J. Comp. Pathol. 1998, 119, 1–14. [Google Scholar] [CrossRef]
- Licitra, B.N.; Whittaker, G.R.; Dubovi, E.J.; Duhamel, G.E. Genotypic characterization of canine coronaviruses associated with fatal canine neonatal enteritis in the United States. J. Clin. Microbiol. 2014, 52, 4230–4238. [Google Scholar] [CrossRef] [Green Version]
- Priestnall, S.L.; Mitchell, J.A.; Walker, C.A.; Erles, K.; Brownlie, J. New and Emerging Pathogens in Canine Infectious Respiratory Disease. Vet. Pathol. 2014, 51, 492–504. [Google Scholar] [CrossRef] [Green Version]
- Tilocca, B.; Soggiu, A.; Musella, V.; Britti, D.; Sanguinetti, M.; Urbani, A.; Roncada, P. Molecular basis of COVID-19 relationships in different species: A One Health perspective. Microbes Infect. 2020, 22, 218–220. [Google Scholar] [CrossRef]
- Priestnall, S.L. Canine Respiratory Coronavirus: A Naturally Occurring Model of COVID-19? Vet. Pathol. 2020, 57, 467–471. [Google Scholar] [CrossRef]
- Xia, X. Extreme Genomic CpG Deficiency in SARS-CoV-2 and Evasion of Host Antiviral Defense. Mol. Biol. Evol. 2020, 37, 2699–2705. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, P.J.; Hatta, M.; Chiba, S.; Maemura, T.; Fan, S.; Takeda, M.; Kinoshita, N.; Hattori, S.I.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; et al. Transmission of SARS-CoV-2 in Domestic Cats. N. Engl. J. Med. 2020, 383, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS–coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.H.; Kiupel, M.; West, K.H.; Raymond, J.T.; Grant, C.K.; Glickman, L.T. Coronavirus-associated epizootic catarrhal enteritis in ferrets. J. Am. Vet. Med. Assoc. 2000, 217, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.; Kiupel, M.; Maes, R.K. Ferret coronavirus-associated diseases. Vet. Clin. N. Am. Exot. Anim. Pract. 2010, 13, 543–560. [Google Scholar] [CrossRef] [PubMed]
- Wise, A.G.; Kiupel, M.; Maes, R.K. Molecular characterization of a novel coronavirus associated with epizootic catarrhal enteritis (ECE) in ferrets. Virology 2006, 349, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Autieri, C.R.; Miller, C.L.; Scott, K.E.; Kilgore, A.; Papscoe, V.A.; Garner, M.M.; Haupt, J.L.; Bakthavatchalu, V.; Muthupalani, S.; Fox, J.G. Systemic Coronaviral Disease in 5 Ferrets. Comp. Med. 2015, 65, 508–516. [Google Scholar]
- Lescano, J.; Quevedo, M.; Gonzales-Viera, O.; Luna, L.; Keel, M.K.; Gregori, F. First Case of Systemic Coronavirus Infection in a Domestic Ferret (Mustela putorius furo) in Peru. Transbound. Emerg. Dis. 2015, 62, 581–585. [Google Scholar] [CrossRef]
- Doria-Torra, G.; Vidaña, B.; Ramis, A.; Amarilla, S.P.; Martínez, J. Coronavirus Infection in Ferrets: Antigen Distribution and Inflammatory Response. Vet. Pathol. 2016, 53, 1180–1186. [Google Scholar] [CrossRef] [Green Version]
- Fujii, Y.; Tochitani, T.; Kouchi, M.; Matsumoto, I.; Yamada, T.; Funabashi, H. Glomerulonephritis in a ferret with feline coronavirus infection. J. Vet. Diagn. Investig. 2015, 27, 637–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wills, S.E.; Beaufrère, H.H.; Brisson, B.A.; Fraser, R.S.; Smith, D.A. Pancreatitis and systemic coronavirus infection in a ferret (Mustela putorius furo). Comp. Med. 2018, 68, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T.; et al. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: An experimental transmission study. Lancet Microbe 2020, 1, e218–e225. [Google Scholar] [CrossRef]
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Chang, J.H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 2020, 27, 704–709.e2. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.; Kok, A.; de Meulder, D.; Bestebroer, T.M.; Lamers, M.M.; Okba, N.M.A.; Fentener van Vlissingen, M.; Rockx, B.; Haagmans, B.L.; Koopmans, M.P.G.; et al. SARS-CoV-2 is transmitted via contact and via the air between ferrets. Nat. Commun. 2020, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Boileau, M.J.; Kapil, S. Bovine Coronavirus Associated Syndromes. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 123–146. [Google Scholar] [CrossRef]
- Decaro, N.; Campolo, M.; Desario, C.; Cirone, F.; D’Abramo, M.; Lorusso, E.; Greco, G.; Mari, V.; Colaianni, M.L.; Elia, G.; et al. Respiratory disease associated with bovine coronavirus infection in cattle herds in Southern Italy. J. Vet. Diagn. Investig. 2008, 20, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Decaro, N.; Mari, V.; Desario, C.; Campolo, M.; Elia, G.; Martella, V.; Greco, G.; Cirone, F.; Colaianni, M.L.; Cordioli, P.; et al. Severe outbreak of bovine coronavirus infection in dairy cattle during the warmer season. Vet. Microbiol. 2008, 126, 30–39. [Google Scholar] [CrossRef]
- Lin, X.Q.; O’Reilly, K.L.; Storz, J. Antibody responses of cattle with respiratory coronavirus infections during pathogenesis of shipping fever pneumonia are lower with antigens of enteric strains than with those of a respiratory strain. Clin. Diagn. Lab. Immunol. 2002, 9, 1010–1013. [Google Scholar] [CrossRef] [Green Version]
- Saif, L.J. Bovine respiratory coronavirus. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 349–364. [Google Scholar] [CrossRef]
- McNulty, M.S.; Bryson, D.G.; Allan, G.M.; Logan, E.F. Coronavirus infection of the bovine respiratory tract. Vet. Microbiol. 1984, 9, 425–434. [Google Scholar] [CrossRef]
- Hick, P.; Read, A.; Lugton, I.; Busfield, F.; Dawood, K.; Gabor, L.; Hornitzky, M.; Kirkland, P. Coronavirus infection in intensively managed cattle with respiratory disease. Aust. Vet. J. 2012, 90, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Storz, J.; Lin, X.; Purdy, C.W.; Chouljenko, V.N.; Kousoulas, K.G.; Enright, F.M.; Gilmore, W.C.; Briggs, R.E.; Loan, R.W. Coronavirus and Pasteurella infections in bovine shipping fever pneumonia and Evans’ criteria for causation. J. Clin. Microbiol. 2000, 38, 3291–3298. [Google Scholar] [CrossRef] [PubMed]
- Kapil, S.; Trent, A.M.; Goyal, S.M. Excretion and persistence of bovine coronavirus in neonatal calves. Arch. Virol. 1990, 115, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulton, R.W.; Herd, H.R.; Sorensen, N.J.; Confer, A.W.; Ritchey, J.W.; Ridpath, J.F.; Burge, L.J. Enteric disease in postweaned beef calves associated with Bovine coronavirus clade 2. J. Vet. Diagn. Investig. 2015, 27, 97–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.-O.; Halbur, P.G.; Bruna, J.D.; Sorden, S.D.; Yoon, K.-J.; Janke, B.H.; Chang, K.-O.; Saif, L.J. Detection and isolation of coronavirus from feces of three herds of feedlot cattle during outbreaks of winter dysentery-like disease. J. Am. Vet. Med. Assoc. 2000, 217, 1191–1194. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, R.; Singh, K.P.; Singh, V.; Malik, Y.P.S.; Kamdi, B.; Singh, R.; Kashyap, G. Immunohistochemical and molecular detection of natural cases of bovine rotavirus and coronavirus infection causing enteritis in dairy calves. Microb. Pathog. 2020, 138, 103814. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Redman, D.R.; Moorhead, P.D.; Theil, K.W. Experimentally induced coronavirus infections in calves: Viral replication in the respiratory and intestinal tracts. Am. J. Vet. Res. 1986, 47, 1426–1432. [Google Scholar] [PubMed]
- Ellis, J. What is the evidence that bovine coronavirus is a biologically significant respiratory pathogen in cattle? Can. Vet. J. 2019, 60, 147–152. [Google Scholar] [PubMed]
- Park, S.J.; Kim, G.Y.; Choy, H.E.; Hong, Y.J.; Saif, L.J.; Jeong, J.H.; Park, S.I.; Kim, H.H.; Kim, S.K.; Shin, S.S.; et al. Dual enteric and respiratory tropisms of winter dysentery bovine coronavirus in calves. Arch. Virol. 2007, 152, 1885–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valarcher, J.-F.; Taylor, G. Bovine respiratory syncytial virus infection. Vet. Res. 2007, 38, 153–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amer, H.M. Bovine-like coronaviruses in domestic and wild ruminants. Anim. Heal. Res. Rev. 2018, 19, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandeil, A.; Gomaa, M.; Shehata, M.; El-Taweel, A.; Kayed, A.E.; Abiadh, A.; Jrijer, J.; Moatasim, Y.; Kutkat, O.; Bagato, O.; et al. Middle East respiratory syndrome coronavirus infection in non-camelid domestic mammals. Emerg. Microbes Infect. 2019, 8, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemoto, M.; Kanno, T.; Bannai, H.; Tsujimura, K.; Yamanaka, T.; Kokado, H. Antibody response to equine coronavirus in horses inoculated with a bovine coronavirus vaccine. J. Vet. Med. Sci. 2017, 79, 1889–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, J.S.; Breslin, J.J.; Breuhaus, B.; Vivrette, S.; Smith, L.G. Characterization of a coronavirus isolated from a diarrheic foal. J. Clin. Microbiol. 2000, 38, 4523–4526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusterla, N.; Mapes, S.; Wademan, C.; White, A.; Ball, R.; Sapp, K.; Burns, P.; Ormond, C.; Butterworth, K.; Bartol, J.; et al. Emerging outbreaks associated with equine coronavirus in adult horses. Vet. Microbiol. 2013, 162, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Vin, R.; Leutenegger, C.; Mittel, L.D.; Divers, T.J. Equine coronavirus: An emerging enteric virus of adult horses. Equine Vet. Educ. 2016, 28, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Fielding, C.L.; Higgins, J.K.; Higgins, J.C.; Mcintosh, S.; Scott, E.; Giannitti, F.; Mete, A.; Pusterla, N. Disease Associated with Equine Coronavirus Infection and High Case Fatality Rate. J. Vet. Intern. Med. 2015, 29, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Giannitti, F.; Diab, S.; Mete, A.; Stanton, J.B.; Fielding, L.; Crossley, B.; Sverlow, K.; Fish, S.; Mapes, S.; Scott, L.; et al. Necrotizing Enteritis and Hyperammonemic Encephalopathy Associated With Equine Coronavirus Infection in Equids. Vet. Pathol. 2015, 52, 1148–1156. [Google Scholar] [CrossRef] [Green Version]
- Narita, M.; Nobumoto, K.; Takeda, H.; Moriyama, T.; Morita, Y.; Nakaoka, Y. Prevalence of Disease with Inference of Equine Coronavirus Infection Among Horses Stabled in a Draft-Horse Racecourse. J. Jpn. Vet. Med. Assoc. 2011, 64, 535–539. [Google Scholar] [CrossRef] [Green Version]
- Oue, Y.; Ishihara, R.; Edamatsu, H.; Morita, Y.; Yoshida, M.; Yoshima, M.; Hatama, S.; Murakami, K.; Kanno, T. Isolation of an equine coronavirus from adult horses with pyrogenic and enteric disease and its antigenic and genomic characterization in comparison with the NC99 strain. Vet. Microbiol. 2011, 150, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Oue, Y.; Morita, Y.; Kondo, T.; Nemoto, M. Epidemic of equine coronavirus at obihiro racecourse, Hokkaido, Japan in 2012. J. Vet. Med. Sci. 2013, 75, 1261–1265. [Google Scholar] [CrossRef] [Green Version]
- Miszczak, F.; Tesson, V.; Kin, N.; Dina, J.; Balasuriya, U.B.R.; Pronost, S.; Vabret, A. First detection of equine coronavirus (ECoV) in Europe. Vet. Microbiol. 2014, 171, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.; Rush, B.R.; Cox, J.; DeBey, B.; Kapil, S. Neonatal enterocolitis associated with coronavirus infection in a foal: A case report. J. Vet. Diagn. Investig. 2000, 12, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Pusterla, N.; Vin, R.; Leutenegger, C.M.; Mittel, L.D.; Divers, T.J. Enteric coronavirus infection in adult horses. Vet. J. 2018, 231, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Gonzales Arguedas, M. Coronavirus infections. In Equine Infectious Diseases; Sellon, D., Long, M., Eds.; Saunders Elsevier: Amsterdam, The Netherlands, 2007; pp. 184–185. [Google Scholar]
- Moon, H.W.; Norman, J.O.; Lambert, G. Age dependent resistance to transmissible gastroenteritis of swine (TGE). I. Clinical signs and some mucosal dimensions in small intestine. Can. J. Comp. Med. Rev. Can. Med. Comp. 1973, 37, 157–166. [Google Scholar]
- Kemeny, L.; Wiltsey, V.; Riley, J. Upper respiratory infection of lactating sows with transmissible gastroenteritis virus following contact exposure to infected piglets. Cornell Vet. 1975, 65, 352–362. [Google Scholar] [PubMed]
- Saif, L.J. Animal coronavirus vaccines: Lessons for SARS. Dev. Biol. (Basel) 2004, 119, 129–140. [Google Scholar]
- Saif, L.J. Animal coronaviruses: What can they teach us about the severe acute respiratory syndrome? Rev. Sci. Tech. Off. Int. Epiz. 2004, 23, 643–660. [Google Scholar] [CrossRef]
- Shoup, D.I.; Swayne, D.E.; Jackwood, D.J.; Saif, L.J. Immunohistochemistry of transmissible gastroenteritis virus antigens in fixed paraffin-embedded tissues. J. Vet. Diagn. Investig. 1996, 8, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Thake, D.C. Jejunal epithelium in transmissible gastroenteritis of swine. An electron microscopic and histochemical study. Am. J. Pathol. 1968, 53, 149–168. [Google Scholar] [PubMed]
- Pensaert, M.B.; Debouck, P.; Reynolds, D.J. An immunoelectron microscopic and immunofluorescent study on the antigenic relationship between the coronavirus-like agent, CV 777, and several coronaviruses. Arch. Virol. 1981, 68, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, G.W.; Hoang, H.; Schwartz, K.J.; Burrough, E.R.; Sun, D.; Madson, D.; Cooper, V.L.; Pillatzki, A.; Gauger, P.; Schmitt, B.J.; et al. Emergence of Porcine epidemic diarrhea virus in the United States: Clinical signs, lesions, and viral genomic sequences. J. Vet. Diagn. Investig. 2013, 25, 649–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, K.; Wang, Q.; Scheuer, K.A.; Lu, Z.; Zhang, Y.; Saif, L.J. Pathology of US porcine epidemic diarrhea virus strain PC21A in gnotobiotic pigs. Emerg. Infect. Dis. 2014, 20, 662–665. [Google Scholar] [CrossRef] [PubMed]
- Madson, D.M.; Magstadt, D.R.; Arruda, P.H.E.; Hoang, H.; Sun, D.; Bower, L.P.; Bhandari, M.; Burrough, E.R.; Gauger, P.C.; Pillatzki, A.E.; et al. Pathogenesis of porcine epidemic diarrhea virus isolate (US/Iowa/18984/2013) in 3-week-old weaned pigs. Vet. Microbiol. 2014, 174, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Niederwerder, M.C.; Hesse, R.A. Swine enteric coronavirus disease: A review of 4 years with porcine epidemic diarrhoea virus and porcine deltacoronavirus in the United States and Canada. Transbound. Emerg. Dis. 2018, 65, 660–675. [Google Scholar] [CrossRef] [Green Version]
- Saif, L.; Pensaert, M.; Sestak, K.; Yeo, S.; Jung, K. Coronaviruses. In Diseases of Swine; Zimmerman, J., Ed.; Elsevier Science BV: Amsterdam, The Netherlands, 2012; pp. 501–524. [Google Scholar]
- Wang, L.; Hayes, J.; Sarver, C.; Byrum, B.; Zhang, Y. Porcine deltacoronavirus: Histological lesions and genetic characterization. Arch. Virol. 2016, 161, 171–175. [Google Scholar] [CrossRef]
- Park, J.; Shin, H. Porcine epidemic diarrhea virus infects and replicates in porcine alveolar macrophages. Virus Res. 2014, 191, 143–152. [Google Scholar] [CrossRef]
- Kawaguchi, H.; Horie, M.; Onoue, K.; Noguchi, M.; Akioka, K.; Masatani, T.; Miura, N.; Ozawa, M.; Tanimoto, A. Development of a Model of Porcine Epidemic Diarrhea in Microminipigs. Vet. Pathol. 2019, 56, 711–714. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.; Liang, X.; Lou, F.; Oglesbee, M.; Krakowka, S.; Li, J. Origin, Evolution, and Virulence of Porcine Deltacoronaviruses in the United States. MBio 2015, 6, 1472–1474. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Hu, H.; Eyerly, B.; Lu, Z.; Chepngeno, J.; Saif, L.J. Pathogenicity of 2 porcine deltacoronavirus strains in gnotobiotic pigs. Emerg. Infect. Dis. 2015, 21, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Vitosh-Sillman, S.; Loy, J.D.; Brodersen, B.; Kelling, C.; Doster, A.; Topliff, C.; Nelson, E.; Bai, J.; Schirtzinger, E.; Poulsen, E.; et al. Experimental infection of conventional nursing pigs and their dams with Porcine deltacoronavirus. J. Vet. Diagn. Investig. 2016, 28, 486–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Gauger, P.; Stafne, M.; Thomas, J.; Arruda, P.; Burrough, E.; Madson, D.; Brodie, J.; Magstadt, D.; Derscheid, R.; et al. Pathogenicity and pathogenesis of a United States porcine deltacoronavirus cell culture isolate in 5-day-old neonatal piglets. Virology 2015, 482, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 2018, 556, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Vlasova, A.N.; Kenney, S.P.; Saif, L.J. Emerging and re-emerging coronaviruses in pigs. Curr. Opin. Virol. 2019, 34, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Han, Y.; Shi, H.; Chen, J.; Zhang, X.; Wang, X.; Zhou, L.; Liu, J.; Zhang, J.; Ji, Z.; et al. Swine acute diarrhea syndrome coronavirus-induced apoptosis is caspase- and cyclophilin D- dependent. Emerg. Microbes Infect. 2020, 9, 439–456. [Google Scholar] [CrossRef]
- Mora-Díaz, J.C.; Piñeyro, P.E.; Houston, E.; Zimmerman, J.; Giménez-Lirola, L.G. Porcine hemagglutinating encephalomyelitis virus: A review. Front. Vet. Sci. 2019, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, B.; Lu, H.; Zhao, K.; Liu, W.; Gao, W.; Lan, Y.; Zhao, J.; Tang, B.; Song, D.; He, W.; et al. Identification and genetic characterization of porcine hemagglutinating encephalomyelitis virus from domestic piglets in China. Arch. Virol. 2014, 159, 2329–2337. [Google Scholar] [CrossRef]
- Quiroga, M.A.; Cappuccio, J.; Piñeyro, P.; Basso, W.; Moré, G.; Kienast, M.; Schonfeld, S.; Cáncer, J.L.; Arauz, S.; Pintos, M.E.; et al. Hemagglutinating encephalomyelitis coronavirus infection in pigs, Argentina. Emerg. Infect. Dis. 2008, 14, 484–486. [Google Scholar] [CrossRef]
- Cox, E.; Hooyberghs, J.; Pensaert, M.B. Sites of replication of a porcine respiratory coronavirus related to transmissible gastroenteritis virus. Res. Vet. Sci. 1990, 48, 165–169. [Google Scholar] [CrossRef]
- Opriessnig, T.; Giménez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Evermann, J.F.; Foreyt, W.; Maag-Miller, L.; Leathers, C.W.; McKeirnan, A.J.; LeaMaster, B. Acute hemorrhagic enteritis associated with canine coronavirus and parvovirus infections in a captive coyote population. J. Am. Vet. Med. Assoc. 1980, 177, 784–786. [Google Scholar] [PubMed]
- Foreyt, W.J.; Evermann, J.F. Serologic survey of canine coronavirus in wild coyotes in the western United States, 1972–1982. J. Wildl. Dis. 1985, 21, 428–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsunemitsu, H.; El-Kanawati, Z.R.; Smith, D.R.; Reed, H.H.; Saif, L.J. Isolation of coronaviruses antigenically indistinguishable from bovine coronavirus from wild ruminants with diarrhea. J. Clin. Microbiol. 1995, 33, 3264–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evermann, J.F.; Heeney, J.L.; Roelke, M.E.; McKeirnan, A.J.; O’Brien, S.J. Biological and pathological consequences of feline infectious peritonitis virus infection in the cheetah. Arch. Virol. 1988, 102, 155–171. [Google Scholar] [CrossRef]
- Heeney, J.L.; Evermann, J.F.; McKeirnan, A.J.; Marker-Kraus, L.; Roelke, M.E.; Bush, M.; Wildt, D.E.; Meltzer, D.G.; Colly, L.; Lukas, J. Prevalence and implications of feline coronavirus infections of captive and free-ranging cheetahs (Acinonyx jubatus). J. Virol. 1990, 64, 1964–1972. [Google Scholar] [CrossRef] [Green Version]
- Van Rensburg, I.B.; Silkstone, M.A. Concomitant feline infectious peritonitis and toxoplasmosis in a cheetah (Acinonyx jubatus). J. S. Afr. Vet. Assoc. 1984, 55, 205–207. [Google Scholar]
- Evermann, J.; Roelke, M.; Briggs, M. Feline coronavirus infections of cheetahs. Feline Pract. 1986, 16, 21–28. [Google Scholar]
- Stephenson, N.; Swift, P.; Moeller, R.B.; Worth, S.J.; Foley, J. Feline infectious peritonitis in a mountain lion (Puma concolor), California, USA. J. Wildl. Dis. 2013, 49, 408–412. [Google Scholar] [CrossRef]
- Paul-Murphy, J.; Work, T.; Hunter, D.; McFie, E.; Fjelline, D. Serologic survey and serum biochemical reference ranges of the free-ranging mountail lion (Felis Concolor) in California. J. Wildl. Dis. 1994, 30, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Mwase, M.; Shimada, K.; Mumba, C.; Yabe, J.; Squarre, D.; Madarame, H. Positive Immunolabelling for Feline Infectious Peritonitis in an African Lion (Panthera leo) with Bilateral Panuveitis. J. Comp. Pathol. 2015, 152, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Watt, N.J.; MacIntyre, N.J.; McOrist, S. An extended outbreak of infectious peritonitis in a closed colony of european wildcats (Felis silvestris). J. Comp. Pathol. 1993, 108, 73–79. [Google Scholar] [CrossRef]
- Kim, J.H.; Jang, J.H.; Yoon, S.W.; Noh, J.Y.; Ahn, M.J.; Kim, Y.; Jeong, D.G.; Kim, H.K. Detection of bovine coronavirus in nasal swab of non-captive wild water deer, Korea. Transbound. Emerg. Dis. 2018, 65, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, J.Y.; Kim, H.R.; Bae, Y.C.; Lee, O.S.; Oem, J.K. Detection and characterization of bovine-like coronaviruses from four species of zoo ruminants. Vet. Microbiol. 2011, 148, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Hasoksuz, M.; Alekseev, K.; Vlasova, A.; Zhang, X.; Spiro, D.; Halpin, R.; Wang, S.; Ghedin, E.; Saif, L.J. Biologic, Antigenic, and Full-Length Genomic Characterization of a Bovine-Like Coronavirus Isolated from a Giraffe. J. Virol. 2007, 81, 4981–4990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cebra, C.K.; Mattson, D.E.; Baker, R.J.; Sonn, R.J.; Dearing, P.L. Potential pathogens in feces from unweaned llamas and alpacas with diarrhea. J. Am. Vet. Med. Assoc. 2003, 223, 1806–1808. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Cebra, C.K.; Baker, R.J.; Mattson, D.E.; Cohen, S.A.; Alvarado, D.E.; Rohrmann, G.F. Analysis of the genome sequence of an alpaca coronavirus. Virology 2007, 365, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Vergara-Alert, J.; van den Brand, J.M.A.; Widagdo, W.; Muñoz, M.; Raj, V.S.; Schipper, D.; Solanes, D.; Cordón, I.; Bensaid, A.; Haagmans, B.L.; et al. Livestock susceptibility to infection with middle east respiratory syndrome coronavirus. Emerg. Infect. Dis. 2017, 23, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Crossley, B.M.; Mock, R.E.; Callison, S.A.; Hietala, S.K. Identification and characterization of a novel alpaca respiratory coronavirus most closely related to the human coronavirus 229E. Viruses 2012, 4, 3689–3700. [Google Scholar] [CrossRef] [Green Version]
- Killerby, M.E.; Biggs, H.M.; Midgley, C.M.; Gerber, S.I.; Watson, J.T. Middle east respiratory syndrome coronavirus transmission. Emerg. Infect. Dis. 2020, 26, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Haagmans, B.L.; Al Dhahiry, S.H.S.; Reusken, C.B.E.M.; Raj, V.S.; Galiano, M.; Myers, R.; Godeke, G.J.; Jonges, M.; Farag, E.; Diab, A.; et al. Middle East respiratory syndrome coronavirus in dromedary camels: An outbreak investigation. Lancet Infect. Dis. 2014, 14, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Hemida, M.G.; Elmoslemany, A.; Al-Hizab, F.; Alnaeem, A.; Almathen, F.; Faye, B.; Chu, D.K.W.; Perera, R.A.P.M.; Peiris, M. Dromedary Camels and the Transmission of Middle East Respiratory Syndrome Coronavirus (MERS-CoV). Transbound. Emerg. Dis. 2017, 64, 344–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farag, E.; Sikkema, R.S.; Mohamedani, A.A.; De Bruin, E.; Oude Munnink, B.B.; Chandler, F.; Kohl, R.; Van Der Linden, A.; Okba, N.M.A.; Haagmans, B.L.; et al. MERS-CoV in camels but not camel handlers, Sudan, 2015 and 2017. Emerg. Infect. Dis. 2019, 25, 2333–2335. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.Y.; Lau, S.K.P.; Wernery, U.; Wong, E.Y.M.; Tsang, A.K.L.; Johnson, B.; Yip, C.C.Y.; Lau, C.C.Y.; Sivakumar, S.; Cai, J.P.; et al. Novel betacoronavirus in dromedaries of the Middle East, 2013. Emerg. Infect. Dis. 2014, 20, 560–572. [Google Scholar] [CrossRef] [Green Version]
- Decaro, N.; Martella, V.; Elia, G.; Campolo, M.; Mari, V.; Desario, C.; Lucente, M.S.; Lorusso, A.; Greco, G.; Corrente, M.; et al. Biological and genetic analysis of a bovine-like coronavirus isolated from water buffalo (Bubalus bubalis) calves. Virology 2008, 370, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Decaro, N.; Cirone, F.; Mari, V.; Nava, D.; Tinelli, A.; Elia, G.; Di Sarno, A.; Martella, V.; Colaianni, M.L.; Aprea, G.; et al. Characterisation of bubaline coronavirus strains associated with gastroenteritis in water buffalo (Bubalus bubalis) calves. Vet. Microbiol. 2010, 145, 245–251. [Google Scholar] [CrossRef]
- Majhdi, F.; Minocha, H.C.; Kapil, S. Isolation and characterization of a coronavirus from elk calves with diarrhea. J. Clin. Microbiol. 1997, 35, 2937–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chasey, D.; Reynolds, D.J.; Bridger, J.C.; Debney, T.G.; Scott, A.C. Identification of coronaviruses in exotic species of Bovidae. Vet. Rec. 1984, 115, 602–603. [Google Scholar] [CrossRef]
- Larsen, A.E.; Gorham, J.R. A new mink enteritis: An initial report. Vet. Med. Small Anim. Clin. 1975, 70, 291–292. [Google Scholar]
- Vlasova, A.N.; Halpin, R.; Wang, S.; Ghedin, E.; Spiro, D.J.; Saif, L.J. Molecular characterization of a new species in the genus Alphacoronavirus associated with mink epizootic catarrhal gastroenteritis. J. Gen. Virol. 2011, 92, 1369–1379. [Google Scholar] [CrossRef]
- Oreshkova, N.; Molenaar, R.J.; Vreman, S.; Harders, F.; Oude Munnink, B.B.; Hakze-van der Honing, R.W.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R.S.; et al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Eurosurveillance 2020, 25, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Molnar, B.; Duchamp, C.; Möstl, K.; Diehl, P.A.; Betschart, B. Comparative survey of canine parvovirus, canine distemper virus and canine enteric coronavirus infection in free-ranging wolves of central Italy and south-eastern France. Eur. J. Wildl. Res. 2014, 60, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Alfano, F.; Dowgier, G.; Valentino, M.P.; Galiero, G.; Tinelli, A.; Decaro, N.; Fusco, G. Identification of pantropic canine coronavirus in a wolf (Canis lupus italicus) in Italy. J. Wildl. Dis. 2019, 55, 504–508. [Google Scholar] [PubMed] [Green Version]
- Rosa, G.M.; Santos, N.; Grøndahl-Rosado, R.; Fonseca, F.P.; Tavares, L.; Neto, I.; Cartaxeiro, C.; Duarte, A. Unveiling patterns of viral pathogen infection in free-ranging carnivores of northern Portugal using a complementary methodological approach. Comp. Immunol. Microbiol. Infect. Dis. 2020, 69, 101432. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jing, H.; Xu, H.; Jiang, X.; Kan, B.; Liu, Q.; Wan, K.; Cui, B.; Zheng, H.; Cui, Z.; et al. Surveillance on severe acute respiratory syndrome associated coronavirus in animals at a live animal market of Guangzhou in 2004. Zhonghua Liu Xing Bing Xue Za Zhi 2005, 26, 84–87. [Google Scholar] [PubMed]
- Guan, Y.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Guan, Y.J.; et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in Southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Martin, H.D.; Zeidner, N.S. Concomitant Cryptosporidia, Coronavirus and Parvovirus Infection in a Raccoon (Procyon lotor). J. Wildl. Dis. 1992, 28, 113–115. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.Q.; Liu, W.; Fan, X.H.; Vijaykrishna, D.; Tang, X.C.; Gao, F.; Li, L.F.; Li, G.J.; Zhang, J.X.; Yang, L.Q.; et al. Detection of a Novel and Highly Divergent Coronavirus from Asian Leopard Cats and Chinese Ferret Badgers in Southern China. J. Virol. 2007, 81, 6920–6926. [Google Scholar] [CrossRef] [Green Version]
- Song, H.D.; Tu, C.C.; Zhang, G.W.; Wang, S.Y.; Zheng, K.; Lei, L.C.; Chen, Q.X.; Gao, Y.W.; Zhou, H.Q.; Xiang, H.; et al. Cross-host evolution of severe acute respiratory syndrome coronavirus in palm civet and human. Proc. Natl. Acad. Sci. USA 2005, 102, 2430–2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- East, M.L.; Moestl, K.; Benetka, V.; Pitra, C.; Höner, O.P.; Wachter, B.; Hofer, H. Coronavirus infection of spotted hyenas in the Serengeti ecosystem. Vet. Microbiol. 2004, 102, 1–9. [Google Scholar] [CrossRef]
- Goller, K.V.; Fickel, J.; Hofer, H.; Beier, S.; East, M.L. Coronavirus genotype diversity and prevalence of infection in wild carnivores in the Serengeti National Park, Tanzania. Arch. Virol. 2013, 158, 729–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corman, V.M.; Kallies, R.; Philipps, H.; Gopner, G.; Muller, M.A.; Eckerle, I.; Brunink, S.; Drosten, C.; Drexler, J.F. Characterization of a Novel Betacoronavirus Related to Middle East Respiratory Syndrome Coronavirus in European Hedgehogs. J. Virol. 2014, 88, 717–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monchatre-Leroy, E.; Boué, F.; Boucher, J.-M.; Renault, C.; Moutou, F.; Ar Gouilh, M.; Umhang, G. Identification of Alpha and Beta Coronavirus in Wildlife Species in France: Bats, Rodents, Rabbits, and Hedgehogs. Viruses 2017, 9, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delogu, M.; Cotti, C.; Lelli, D.; Sozzi, E.; Trogu, T.; Lavazza, A.; Garuti, G.; Castrucci, M.R.; Vaccari, G.; De Marco, M.A.; et al. Eco-Virological Preliminary Study of Potentially Emerging Pathogens in Hedgehogs (Erinaceus europaeus) Recovered at a Wildlife Treatment and Rehabilitation Center in Northern Italy. Animals 2020, 10, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Lin, X.-D.; Liao, Y.; Guan, X.-Q.; Guo, W.-P.; Xing, J.-G.; Holmes, E.C.; Zhang, Y.-Z. Discovery of a Highly Divergent Coronavirus in the Asian House Shrew from China Illuminates the Origin of the Alphacoronaviruses. J. Virol. 2017, 91, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrono, L.V.; Samuni, L.; Corman, V.M.; Nourifar, L.; Röthemeier, C.; Wittig, R.M.; Drosten, C.; Calvignac-Spencer, S.; Leendertz, F.H. Human coronavirus OC43 outbreak in wild chimpanzees, Côte d’Ivoire, 2016 correspondence. Emerg. Microbes Infect. 2018, 7, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Mihindukulasuriya, K.A.; Wu, G.; St. Leger, J.; Nordhausen, R.W.; Wang, D. Identification of a Novel Coronavirus from a Beluga Whale by Using a Panviral Microarray. J. Virol. 2008, 82, 5084–5088. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Tsang, A.K.L.; Hui, S.-W.; Fan, R.Y.Y.; Martelli, P.; Yuen, K.-Y. Discovery of a Novel Bottlenose Dolphin Coronavirus Reveals a Distinct Species of Marine Mammal Coronavirus in Gammacoronavirus. J. Virol. 2014, 88, 1318–1331. [Google Scholar] [CrossRef] [Green Version]
- Bryda, E.C. The Mighty Mouse: The impact of rodents on advances in biomedical research. Mo. Med. 2013, 110, 207–211. [Google Scholar]
- Wang, W.; Lin, X.; Guo, W.; Zhou, R.; Wang, M.; Wang, C.-Q.; Ge, S.; Mei, S.-H.; Li, M.-H.; Shi, M.; et al. Discovery, diversity and evolution of novel coronaviruses sampled from rodents in China. Virology 2015, 474, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.Y.; Yang, W.H.; Zhou, J.H.; Li, B.; Zhang, W.; Shi, Z.L.; Zhang, Y.Z. Detection of alpha- and betacoronaviruses in rodents from Yunnan, China. Virol. J. 2017, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, S.K.P.; Woo, P.C.Y.; Li, K.S.M.; Tsang, A.K.L.; Fan, R.Y.Y.; Luk, H.K.H.; Cai, J.-P.; Chan, K.-H.; Zheng, B.-J.; Wang, M.; et al. Discovery of a Novel Coronavirus, China Rattus Coronavirus HKU24, from Norway Rats Supports the Murine Origin of Betacoronavirus 1 and Has Implications for the Ancestor of Betacoronavirus Lineage A. J. Virol. 2015, 89, 3076–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoleridis, T.; Onianwa, O.; Horncastle, E.; Dayman, E.; Zhu, M.; Danjittrong, T.; Wachtl, M.; Behnke, J.M.; Chapman, S.; Strong, V.; et al. Discovery of novel alphacoronaviruses in European rodents and shrews. Viruses 2016, 8, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, R.; Li, F.; Dragomir, I.; Chua, M.M.W.; Gregory, B.D.; Weiss, S.R. Analysis of the Host Transcriptome from Demyelinating Spinal Cord of Murine Coronavirus-Infected Mice. PLoS ONE 2013, 8, e75346c. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparicio, J.L.; Peña, C.; Retegui, L.A. Autoimmune hepatitis-like disease in C57BL/6 mice infected with mouse hepatitis virus A59. Int. Immunopharmacol. 2011, 11, 1591–1598. [Google Scholar] [CrossRef]
- Homberger, F.R. Enterotropic mouse hepatitis virus. Lab. Anim. 1997, 31, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Compton, S.R.; Ball-Goodrich, L.J.; Johnson, L.K.; Johnson, E.A.; Paturzo, F.X.; Macy, J.D. Pathogenesis of enterotropic mouse hepatitis virus in immunocompetent and immunodeficient mice. Comp. Med. 2004, 54, 681–689. [Google Scholar]
- Percy, D.H.; Barthold, S.W. Pathology of Laboratory Rodents and Rabbits, 4th ed.; Barthold, S.W., Griffey, S.M., Percy, D.H., Eds.; John Wiley & Sons, Inc.: Chichester, UK, 2016; ISBN 9781118924051. [Google Scholar]
- Blau, D.M.; Turbide, C.; Tremblay, M.; Olson, M.; Létourneau, S.; Michaliszyn, E.; Jothy, S.; Holmes, K.V.; Beauchemin, N. Targeted Disruption of the Ceacam1(MHVR) Gene Leads to Reduced Susceptibility of Mice to Mouse Hepatitis Virus Infection. J. Virol. 2001, 75, 8173–8186. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, F.; Hirai-Yuki, A. Mouse hepatitis virus receptor as a determinant of the mouse susceptibility to MHV infection. Front. Microbiol. 2012, 3, 1999–2002. [Google Scholar] [CrossRef] [Green Version]
- Leibowitz, J.L.; Srinivasa, R.; Williamson, S.T.; Chua, M.M.; Liu, M.; Wu, S.; Kang, H.; Ma, X.-Z.; Zhang, J.; Shalev, I.; et al. Genetic Determinants of Mouse Hepatitis Virus Strain 1 Pneumovirulence. J. Virol. 2010, 84, 9278–9291. [Google Scholar] [CrossRef] [Green Version]
- Barthold, S.W.; Smith, A.L. Mouse hepatitis virus S in weanling Swiss mice following intranasal inoculation. Lab. Anim. Sci. 1983, 33, 355–360. [Google Scholar] [PubMed]
- Skinner, D.; Marro, B.S.; Lane, T.E. Chemokine CXCL10 and Coronavirus-Induced Neurologic Disease. Viral Immunol. 2019, 32, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Biswas, K.; Chatterjee, D.; Addya, S.; Khan, R.S.; Kenyon, L.C.; Choe, A.; Cohrs, R.J.; Shindler, K.S.; Das Sarma, J. Demyelinating strain of mouse hepatitis virus infection bridging innate and adaptive immune response in the induction of demyelination. Clin. Immunol. 2016, 170, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Shindler, K.S.; Chatterjee, D.; Biswas, K.; Goyal, A.; Dutt, M.; Nassrallah, M.; Khan, R.S.; Das Sarma, J. Macrophage-Mediated Optic Neuritis Induced by Retrograde Axonal Transport of Spike Gene Recombinant Mouse Hepatitis Virus. J. Neuropathol. Exp. Neurol. 2011, 70, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Khan, R.S.; Dine, K.; Das Sarma, J.; Shindler, K.S. Intracranial inoculation is more potent than intranasal inoculation for inducing optic neuritis in the mouse hepatitis virus-induced model of multiple sclerosis. Front. Cell. Infect. Microbiol. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Kishore, A.; Maity, D.; Sunanda, P.; Krishnarjuna, B.; Vappala, S.; Raghothama, S.; Kenyon, L.C.; Pal, D.; Sarma, J. Das A proline insertion-deletion in the spike glycoprotein fusion peptide of mouse hepatitis virus strongly alters neuropathology. J. Biol. Chem. 2019, 294, 8064–8087. [Google Scholar] [CrossRef] [Green Version]
- Kakizaki, M.; Kashiwazaki, H.; Watanabe, R. Mutant murine hepatitis virus-induced apoptosis in the hippocampus. Jpn. J. Infect. Dis. 2014, 67, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Kashiwazaki, H.; Nomura, R.; Matsuyama, S.; Taguchi, F.; Watanabe, R. Spongiform degeneration induced by neuropathogenic murine coronavirus infection. Pathol. Int. 2011, 61, 184–191. [Google Scholar] [CrossRef]
- Hooks, J.J.; Percopo, C.; Wang, Y.; Detrick, B. Retina and retinal pigment epithelial cell autoantibodies are produced during murine coronavirus retinopathy. J. Immunol. 1993, 151, 3381–3389. [Google Scholar]
- Bender, S.J.; Weiss, S.R. Pathogenesis of murine coronavirus in the central nervous system. J. Neuroimmune Pharmacol. 2010, 5, 336–354. [Google Scholar] [CrossRef] [Green Version]
- Tardieu, M.; Goffinet, A.; Harmant-van Rijckevorsel, G.; Lyon, G. Ependymitis, leukoencephalitis, hydrocephalus, and thrombotic vasculitis following chronic infection by mouse hepatitis virus 3 (MHV 3). Acta Neuropathol. 1982, 58, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Virelizier, J.L.; Dayan, A.D.; Allison, A.C. Neuropathological effect of persistent infection of mice by mouse hepatitis virus. Infect. Immun. 1975, 12, 1127–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compton, S.R.; Ball-Goodrich, L.J.; Zeiss, C.J.; Johnson, L.K.; Johnson, E.A.; Macy, J.D. Pathogenesis of mouse hepatitis virus infection in gamma interferon-deficient mice is modulated by co-infection with Helicobacter hepaticus. Comp. Med. 2003, 53, 197–206. [Google Scholar] [PubMed]
- Compton, S.R.; Ball-Goodrich, L.J.; Paturzo, F.X.; Macy, J.D. Transmission of enterotropic mouse hepatitis virus from immunocompetent and immunodeficient mice. Comp. Med. 2004, 54, 29–35. [Google Scholar] [PubMed]
- Manjunath, S.; Kulkarni, P.G.; Nagavelu, K.; Samuel, R.J.; Srinivasan, S.; Ramasamy, N.; Hegde, N.R.; Gudde, R.S. Sero-prevalence of rodent pathogens in India. PLoS ONE 2015, 10, e0131706. [Google Scholar] [CrossRef]
- Percy, D.H.; Lynch, J.A.; Descôteaux, J.P. Central Nervous System Lesions in Suckling Mice and Rats Inoculated Intranasally with Sialodacryoadenitis Virus. Vet. Pathol. 1986, 23, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Compton, S.R.; Vivas-Gonzalez, B.E.; Macy, J.D. Reverse transcriptase polymerase chain reaction-based diagnosis and molecular characterization of a new rat coronavirus strain. Lab. Anim. Sci. 1999, 49, 506–513. [Google Scholar] [PubMed]
- Liang, C.T.; Shih, A.; Chang, Y.H.; Liu, C.W.; Lee, Y.T.; Hsieh, W.C.; Huang, Y.L.; Huang, W.T.; Kuang, C.H.; Lee, K.H.; et al. Microbial contaminations of laboratory mice and rats in Taiwan from 2004 to 2007. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 381–386. [Google Scholar] [PubMed]
- Yoo, D.; Pei, Y.; Christie, N.; Cooper, M. Primary structure of the sialodacryoadenitis virus genome: Sequence of the structural-protein region and its application for differential diagnosis. Clin. Diagn. Lab. Immunol. 2000, 7, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunita, S.; Mori, M.; Terada, E. Sequence Analysis of the Nucleocapsid Protein Gene of Rat Coronavirus SDAV-681. Virology 1993, 193, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.N.; Percy, D.H.; Jonas, A.M. Characterization of the virus of sialodacryoadenitis of rats: A member of the coronavirus group. J. Infect. Dis. 1972, 126, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.J.; Manzer, R.; Miura, T.A.; Groshong, S.D.; Ito, Y.; Travanty, E.A.; Leete, J.; Holmes, K.V.; Mason, R.J. Rat respiratory coronavirus infection: Replication in airway and alveolar epithelial cells and the innate immune response. J. Gen. Virol. 2009, 90, 2956–2964. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.A.; Wang, J.; Holmes, K.V.; Mason, R.J. Rat coronaviruses infect rat alveolar type I epithelial cells and induce expression of CXC chemokines. Virology 2007, 369, 288–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.C.; Schoeb, T.R.; Davis, J.K.; Simecka, J.W.; Cassell, G.H.; Lindsey, J.R. Comparative severity of respiratory lesions of sialodacryoadenitis virus and Sendai virus infections in LEW and F344 rats. Vet. Pathol. 1995, 32, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Bihun, C.G.; Percy, D.H. Morphologic changes in the nasal cavity associated with sialodacryoadenitis virus infection in the Wistar rat. Vet. Pathol. 1995, 32, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wojcinski, Z.W.; Percy, D.H. Sialodacryoadenitis Virus-associated Lesions in the Lower Respiratory Tract of Rats. Vet. Pathol. 1986, 23, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Schoeb, T.R. Respiratory Diseases of Rodents. Vet. Clin. N. Am. Exot. Anim. Pract. 2000, 3, 481–496. [Google Scholar] [CrossRef]
- Wickham, L.A.; Huang, Z.; Lambert, R.W.; Sullivan, D.A. Effect of sialodacryoadenitis virus exposure on acinar epithelial cells from the rat lacrimal gland. Ocul. Immunol. Inflamm. 1997, 5, 181–195. [Google Scholar] [CrossRef]
- Jaax, G.P.; Jaax, N.K.; Petrali, J.P.; Corcoran, K.D.; Vogel, A.P. Coronavirus-like virions associated with a wasting syndrome in guinea pigs. Lab. Anim. Sci. 1990, 40, 375–378. [Google Scholar]
- Liang, L.; He, C.; Lei, M.; Li, S.; Hao, Y.; Zhu, H.; Duan, Q. Pathology of guinea pigs experimentally infected with a novel reovirus and coronavirus isolated from SARS patients. DNA Cell Biol. 2005, 24, 485–490. [Google Scholar] [CrossRef]
- Kerr, P.J.; Donnelly, T.M. Viral Infections of Rabbits. Vet. Clin. N. Am. Exot. Anim. Pract. 2013, 16, 437–468. [Google Scholar] [CrossRef] [PubMed]
- Descôteaux, J.P.; Lussier, G. Experimental infection of young rabbits with a rabbit enteric coronavirus. Can. J. Vet. Res. 1990, 54, 473–476. [Google Scholar]
- Cavanagh, D. Coronaviruses in poultry and other birds. Avian Pathol. 2005, 34, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Miłek, J.; Blicharz-Domańska, K. Coronaviruses in avian species-review with focus on epidemiology and diagnosis in wild birds. J. Vet. Res. 2018, 62, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavi. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, M.M.; Cho, K.O.; Ward, L.A.; Saif, L.J.; Saif, Y.M. Experimental bovine coronavirus in turkey poults and young chickens. Avian Dis. 2001, 45, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.M.; Tang, Y.; Saif, Y.M. Pathogenicity of Turkey coronavirus in Turkeys and chickens. Avian Dis. 2003, 47, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.K.W.; Leung, C.Y.H.; Gilbert, M.; Joyner, P.H.; Ng, E.M.; Tse, T.M.; Guan, Y.; Peiris, J.S.M.; Poon, L.L.M. Avian Coronavirus in Wild Aquatic Birds. J. Virol. 2011, 85, 12815–12820. [Google Scholar] [CrossRef] [Green Version]
- Liais, E.; Croville, G.; Mariette, J.; Delverdier, M.; Lucas, M.N.; Klopp, C.; Lluch, J.; Donnadieu, C.; Guy, J.S.; Corrand, L.; et al. Novel avian coronavirus and fulminating disease in Guinea Fowl, France. Emerg. Infect. Dis. 2014, 20, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Jonassen, C.M.; Kofstad, T.; Larsen, I.L.; Løvland, A.; Handeland, K.; Follestad, A.; Lillehaug, A. Molecular identification and characterization of novel coronaviruses infecting graylag geese (Anser anser), feral pigeons (Columbia livia) and mallards (Anas platyrhynchos). J. Gen. Virol. 2005, 86, 1597–1607. [Google Scholar] [CrossRef]
- Jordan, B.J.; Hilt, D.A.; Poulson, R.; Stallknecht, D.E.; Jackwood, M.W. Identification of avian coronavirus in wild aquatic birds of the central and eastern USA. J. Wildl. Dis. 2015, 51, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, I.N.A.; van Beurden, S.J.; Weerts, E.A.W.S.; Verheije, M.H. The avian coronavirus spike protein. Virus Res. 2014, 194, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Ducatez, M.F. Recommendations for a standardized avian coronavirus (AvCoV) nomenclature: Outcome from discussions within the framework of the European Union COST Action FA1207: “towards control of avian coronaviruses: Strategies for vaccination, diagnosis and surveilla. Avian Pathol. 2016, 45, 602–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catroxo, M.H.B.; Martins, A.M.C.R.P.F.; Petrella, S.; Curi, N.A.; Melo, N. Research of Viral Agent in Free-living Pigeon Feces (Columba livia) in the City of São Paulo, SP, Brazil, for Transmission Electron Microscopy. Int. J. Morphol. 2011, 29, 628–635. [Google Scholar] [CrossRef]
- Zhuang, Q.; Liu, S.; Zhang, X.; Jiang, W.; Wang, K.; Wang, S.; Peng, C.; Hou, G.; Li, J.; Yu, X.; et al. Surveillance and taxonomic analysis of the coronavirus dominant in pigeons in China. Transbound. Emerg. Dis. 2020, 67, 1981–1990. [Google Scholar] [CrossRef] [PubMed]
- Qian, D.H.; Zhu, G.J.; Wu, L.Z.; Hua, G.X. Isolation and characterization of a coronavirus from pigeons with pancreatitis. Am. J. Vet. Res. 2006, 67, 1575–1579. [Google Scholar] [CrossRef]
- Fu, J.; Zhang, G.; Liao, M.; Ren, T.; Xin, C. Study on the S1 gene of the two isolated stain of coronavirus from the wild birds. Chin. J. Prev. Vet. Med. 2005, 27, 380–384. [Google Scholar]
- Barr, D.; Reece, R.; O’Rourke, D.; Button, C.; Faragher, J. Isolation of infectious bronchitis virus from a flock of racing pigeons. Aust. Vet. J. 1988, 65, 228. [Google Scholar] [CrossRef]
- Cook, J.K.A.; Jackwood, M.; Jones, R.C. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012, 41, 239–250. [Google Scholar] [CrossRef]
- Day, J.M.; Gonder, E.; Jennings, S.; Rives, D.; Robbins, K.; Tilley, B.; Wooming, B. Investigating turkey enteric coronavirus circulating in the Southeastern United States and Arkansas during 2012 and 2013. Avian Dis. 2014, 58, 313–317. [Google Scholar] [CrossRef]
- Liu, S.; Chen, J.; Chen, J.; Kong, X.; Shao, Y.; Han, Z.; Feng, L.; Cai, X.; Gu, S.; Liu, M. Isolation of avian infectious bronchitis coronavirus from domestic peafowl (Pavo cristatus) and teal (Anas). J. Gen. Virol. 2005, 86, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Ghany, H.M.A.; Elseddawy, N.M. Diagnostic Studies of Infectious Bronchitis Disease in Broilers using Pathological and Molecular Investigations in Kaliobeya Governorate, Egypt. Adv. Environ. Biol. 2019, 13, 1–6. [Google Scholar]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackwood, M. Review of infectious bronchitis virus around the world. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, J.J.S.; Cook, J.K.A.; van der Heijden, H.M.J.F. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Tian, Y.; Guan, R.; Gao, W.; Yang, X.; Zhou, L.; Wang, H. Infectious Bronchitis Virus Infection Induces Apoptosis during Replication in Chicken Macrophage HD11 Cells. Viruses 2017, 9, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amarasinghe, A.; Abdul-Cader, M.S.; Almatrouk, Z.; van der Meer, F.; Cork, S.C.; Gomis, S.; Abdul-Careem, M.F. Induction of innate host responses characterized by production of interleukin (IL)-1β and recruitment of macrophages to the respiratory tract of chickens following infection with infectious bronchitis virus (IBV). Vet. Microbiol. 2018, 215, 1–10. [Google Scholar] [CrossRef]
- Reddy, V.R.A.P.; Trus, I.; Desmarets, L.M.B.; Li, Y.; Theuns, S.; Nauwynck, H.J. Productive replication of nephropathogenic infectious bronchitis virus in peripheral blood monocytic cells, a strategy for viral dissemination and kidney infection in chickens. Vet. Res. 2016, 47, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.S.; Liu, D.X. Human Coronavirus: Host-Pathogen Interaction. Annu. Rev. Microbiol. 2019, 73, 529–557. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; He, X.; Du, L.J.; Liu, Y.Y.; You, G.J.; Li, S.Y.; Liu, P.; Cao, S.J.; Han, X.F.; Huang, Y. Preparation and protective efficacy of a chicken embryo kidney cell-attenuation GI-19/QX-like avian infectious bronchitis virus vaccine. Vaccine 2018, 36, 4087–4094. [Google Scholar] [CrossRef]
- Abou El-Fetouh, M.; Mohamed, M.; Refat, N.; Ahmed, M.; El-Zanaty, A.E. Pathological Studies on Infectious Bronchitis Disease in Chickens. Zagazig Vet. J. 2016, 44, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Shivaprasad, H.L. Pathology of birds—An overview. In Proceedings of the C.L. Davis Foundation Conference on Gross Morbid Anatomy of Animals, Washington, DC, USA, 8–12 April 2002; pp. 1–50. [Google Scholar]
- Jackwood, M.; De Wit, S. Infectious bronchitis. In Diseases of Poultry; Swayne, D., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 167–188. [Google Scholar]
- Yu, L.; Wang, Z.; Jiang, Y.; Low, S.; Kwang, J. Molecular Epidemiology of Infectious Bronchitis Virus Isolates from China and Southeast Asia. Avian Dis. 2001, 45, 201. [Google Scholar] [CrossRef] [PubMed]
- Owen, R.L.; Cowen, B.S.; Hattel, A.L.; Naqi, S.A.; Wilson, R.A. Detection of viral antigen following exposure of one-day-old chickens to the Holland 52 strain of infectious bronchitis virus. Avian Pathol. 1991, 20, 663–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toro, H.; Godoy, V.; Larenas, J.; Reyes, E.; Kaleta, E.F. Avian infectious bronchitis: Viral persistence in the harderian gland and histological changes after eyedrop vaccination. Avian Dis. 1996, 40, 114–120. [Google Scholar] [CrossRef]
- Van Ginkel, F.; Van Santen, V.; Gulley, S.; Toro, H. Infectious bronchitis virus in the chicken Harderian gland and lachrymal fluid: Viral load, infectivity, immune cell responses, and effects of viral immunodeficiency. Avian Dis. 2008, 52, 608–617. [Google Scholar] [CrossRef]
- Gough, R.E.; Cox, W.J.; Winkler, C.E.; Sharp, M.W.; Spackman, D. Isolation and identification of infectious bronchitis virus from pheasants. Vet. Rec. 1996, 138, 208–209. [Google Scholar] [CrossRef]
- Lister, S.; Beer, J.; Gough, R.; Holmes, R.; JMW, J.; Orton, R. Outbreaks of nephritis in pheasants (phasianus colchicus) with a possible coronavirus aetiology. Vet. Rec. 1985, 117, 612–613. [Google Scholar] [CrossRef]
- Cavanagh, D.; Mawditt, K.; Welchman, D.D.B.; Britton, P.; Gough, R.E. Coronaviruses from pheasants (Phasianus colchicus) are genetically closely related to coronaviruses of domestic fowl (infectious bronchitis virus) and turkeys. Avian Pathol. 2002, 31, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Spackman, D.; Cameron, I. Isolation of infectious bronchitis virus from pheasants. Vet. Rec. 1983, 113, 354–355. [Google Scholar] [CrossRef]
- Han, Z.; Liwen, X.; Ren, M.; Sheng, J.; Ma, T.; Sun, J.; Zhao, Y.; Liu, S. Genetic, antigenic and pathogenic characterization of avian coronaviruses isolated from pheasants (Phasianus colchicus) in China. Vet. Microbiol. 2020, 240, 108513. [Google Scholar] [CrossRef]
- Pennycott, T.W. Causes of mortality and culling in adult pheasants. Vet. Rec. 2000, 146, 273–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welchman, D.B.; Bradbury, J.M.; Cavanagh, D.; Aebischer, N.J. Infectious agents associated with respiratory disease in pheasants. Vet. Rec. 2002, 150, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Qian, H.; Yao, C. Isolation and identification of pigeon coronavirus. J. Shanghai Jiaotong Univ. Agric. Sci. 2005, 23, 275–279. [Google Scholar]
- Brown, P.A.; Courtillon, C.; Weerts, E.A.W.S.; Andraud, M.; Allée, C.; Vendembeuche, A.; Amelot, M.; Rose, N.; Verheije, M.H.; Eterradossi, N. Transmission Kinetics and histopathology induced by European Turkey Coronavirus during experimental infection of specific pathogen free turkeys. Transbound. Emerg. Dis. 2019, 66, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, M.H.; Yoo, D.; Ojkic, D.; Barta, J.R. Infection with a pathogenic turkey coronavirus isolate negatively affects growth performance and intestinal morphology of young turkey poults in Canada. Avian Pathol. 2009, 38, 279–286. [Google Scholar] [CrossRef]
- Gomaa, M.H.; Barta, J.R.; Ojkic, D.; Yoo, D. Complete genomic sequence of turkey coronavirus. Virus Res. 2008, 135, 237–246. [Google Scholar] [CrossRef]
- Chien, C.L.; Ching, C.W.; Tsang, L.L. Comparison of 3′-end encoding regions of Turkey coronavirus isolates from Indiana, North Carolina, and Minnesota with chicken infectious bronchitis coronavirus strains. Intervirology 2006, 49, 230–238. [Google Scholar]
- Wang, Y.; Cui, X.; Chen, X.; Yang, S.; Ling, Y.; Song, Q.; Zhu, S.; Sun, L.; Li, C.; Li, Y.; et al. A recombinant infectious bronchitis virus from a chicken with a spike gene closely related to that of a turkey coronavirus. Arch. Virol. 2020, 165, 703–707. [Google Scholar] [CrossRef] [Green Version]
- Bouwman, K.M.; Delpont, M.; Broszeit, F.; Berger, R.; Weerts, E.A.W.S.; Lucas, M.-N.; Delverdier, M.; Belkasmi, S.; Papanikolaou, A.; Boons, G.-J.; et al. Guinea Fowl Coronavirus Diversity Has Phenotypic Consequences for Glycan and Tissue Binding. J. Virol. 2019, 93, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Circella, E.; Camarda, A.; Martella, V.; Bruni, G.; Lavazza, A.; Buonavoglia, C. Coronavirus associated with an enteric syndrome on a quail farm. Avian Pathol. 2007, 36, 251–258. [Google Scholar] [CrossRef]
- Torres, C.A.; Villarreal, L.Y.B.; Ayres, G.R.R.; Richtzenhain, L.J.; Brandão, P.E. An avian coronavirus in quail with respiratory and reproductive signs. Avian Dis. 2013, 57, 295–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gough, R.E.; Drury, S.E.; Culver, F.; Britton, P.; Cavanagh, D. Isolation of a coronavirus from a green-cheeked Amazon parrot (Amazon viridigenalis Cassin). Avian Pathol. 2006, 35, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Muradrasoli, S.; Bálint, Á.; Wahlgren, J.; Waldenström, J.; Belák, S.; Blomberg, J.; Olsen, B. Prevalence and Phylogeny of Coronaviruses in Wild Birds from the Bering Strait Area (Beringia). PLoS ONE 2010, 5, e13640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granzow, H.; Weiland, F.; Fichtner, D.; Schütze, H.; Karger, A.; Mundt, E.; Dresenkamp, B.; Martin, P.; Mettenleiter, T.C. Identification and ultrastructural characterization of a novel virus from fish. J. Gen. Virol. 2001, 82, 2849–2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schütze, H. Coronaviruses in Aquatic Organisms. In Aquaculture Virology; Elsevier: Amsterdam, The Netherlands, 2016; pp. 327–335. [Google Scholar]
- Huchzermeyer, F.W.; Gerdes, G.H.; Foggin, C.M.; Huchzermeyer, K.D.; Limper, L.C. Hepatitis in farmed hatchling Nile crocodiles (Crocodylus niloticus) due to chlamydial infection. J. S. Afr. Vet. Assoc. 1994, 65, 20–22. [Google Scholar] [PubMed]
- Chams, N.; Chams, S.; Badran, R.; Shams, A.; Araji, A.; Raad, M.; Mukhopadhyay, S.; Stroberg, E.; Duval, E.J.; Barton, L.M.; et al. COVID-19: A Multidisciplinary Review. Front. Public Heal. 2020, 8, 1–20. [Google Scholar] [CrossRef]
- Marraha, F.; Al Faker, I.; Gallouj, S. A Review of the Dermatological Manifestations of Coronavirus Disease 2019 (COVID-19). Dermatol. Res. Pract. 2020, 2020, 9360476. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, X.; Chen, Z.; Duan, J.; Hashimoto, K.; Yang, L.; Liu, C.; Yang, C. Nervous system involvement after infection with COVID-19 and other coronaviruses. Brain. Behav. Immun. 2020, 87, 18–22. [Google Scholar] [CrossRef]
- Gu, J.; Korteweg, C. Pathology and pathogenesis of severe acute respiratory syndrome. Am. J. Pathol. 2007, 170, 1136–1147. [Google Scholar] [CrossRef] [Green Version]
- Hwang, D.M.; Chamberlain, D.W.; Poutanen, S.M.; Low, D.E.; Asa, S.L.; Butany, J. Pulmonary pathology of severe acute respiratory syndrome in Toronto. Mod. Pathol. 2005, 18, 1–10. [Google Scholar] [CrossRef]
- Van Den Brand, J.M.A.; Smits, S.L.; Haagmans, B.L. Pathogenesis of Middle East respiratory syndrome coronavirus. J. Pathol. 2015, 235, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsaad, K.O.; Hajeer, A.H.; Al Balwi, M.; Al Moaiqel, M.; Al Oudah, N.; Al Ajlan, A.; AlJohani, S.; Alsolamy, S.; Gmati, G.E.; Balkhy, H.; et al. Histopathology of Middle East respiratory syndrome coronovirus (MERS-CoV) infection—Clinicopathological and ultrastructural study. Histopathology 2018, 72, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zheng, X.; Tong, Q.; Li, W.; Wang, B.; Sutter, K.; Trilling, M.; Lu, M.; Dittmer, U.; Yang, D. Overlapping and discrete aspects of the pathology and pathogenesis of the emerging human pathogenic coronaviruses SARS-CoV, MERS-CoV, and 2019-nCoV. J. Med. Virol. 2020, 92, 491–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellul, M.A.; Benjamin, L.; Singh, B.; Lant, S.; Michael, B.D.; Easton, A.; Kneen, R.; Defres, S.; Sejvar, J.; Solomon, T. Neurological associations of COVID-19. Lancet Neurol. 2020, 19, 767–783. [Google Scholar] [CrossRef]
- Bradley, B.T.; Maioli, H.; Johnston, R.; Chaudhry, I.; Fink, S.L.; Xu, H.; Najafian, B.; Deutsch, G.; Lacy, J.M.; Williams, T.; et al. Histopathology and ultrastructural findings of fatal COVID-19 infections in Washington State: A case series. Lancet 2020, 396, 320–332. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.M.D.; Stark, H.; Tzankov, A.; et al. Pulmonary vascular endothelialitis, thrombosis, and angiogenesis in Covid-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Hanley, B.; Naresh, K.N.; Roufosse, C.; Nicholson, A.G.; Weir, J.; Cooke, G.S.; Thursz, M.; Manousou, P.; Corbett, R.; Goldin, R.; et al. Histopathological findings and viral tropism in UK patients with severe fatal COVID-19: A post-mortem study. Lancet Microbe 2020, 1, e245–e253. [Google Scholar] [CrossRef]
- Ghosh, A.; Colling, R. An overview of COVID-19 for diagnostic pathologists: Clinicopathological correlation and diagnostic techniques. Diagnostic Histopathol. 2020, 1–8. [Google Scholar] [CrossRef]
- Martines, R.B.; Ritter, J.M.; Matkovic, E.; Gary, J.; Bollweg, B.C.; Bullock, H.; Goldsmith, C.S.; Silva-Flannery, L.; Seixas, J.N.; Reagan-Steiner, S.; et al. Pathology and pathogenesis of SARS-CoV-2 associated with fatal coronavirus disease, united states. Emerg. Infect. Dis. 2020, 26, 2005–2015. [Google Scholar] [CrossRef]
- Infection with SARS-CoV-2 in Animals. Available online: https://www.oie.int/fileadmin/Home/MM/A_Factsheet_SARS-CoV-2__1_.pdf (accessed on 1 October 2020).
- McMahon, B.J.; Morand, S.; Gray, J.S. Ecosystem change and zoonoses in the Anthropocene. Zoonoses Public Health 2018, 65, 755–765. [Google Scholar] [CrossRef]
- Gouilh, M.A.; Puechmaille, S.J.; Gonzalez, J.P.; Teeling, E.; Kittayapong, P.; Manuguerra, J.C. SARS-Coronavirus ancestor’s foot-prints in South-East Asian bat colonies and the refuge theory. Infect. Genet. Evol. 2011, 11, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Baric, R.S.; Fu, K.; Chen, W.; Yount, B. High Recombination and Mutation Rates in Mouse Hepatitis Virus Suggest that Coronaviruses may be Potentially Important Emerging Viruses. In Corona- and Related Viruses. Advances in Experimental Medicine and Biology; Talbot, P., Levy, G., Eds.; Springer: Boston, MA, USA, 1995; pp. 571–576. [Google Scholar]
- Baric, R.S.; Yount, B.; Hensley, L.; Peel, S.A.; Chen, W. Episodic evolution mediates interspecies transfer of a murine coronavirus. J. Virol. 1997, 71, 1946–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thackray, L.B.; Holmes, K.V. Amino acid substitutions and an insertion in the spike glycoprotein extend the host range of the murine coronavirus MHV-A59. Virology 2004, 324, 510–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.Y.; Lau, S.K.P.; Huang, Y.; Yuen, K.Y. Coronavirus diversity, phylogeny and interspecies jumping. Exp. Biol. Med. 2009, 234, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Morse, S.S.; Mazet, J.A.; Woolhouse, M.; Parrish, C.R.; Carroll, D.; Karesh, W.B.; Zambrana-Torrelio, C.; Lipkin, W.I.; Daszak, P. Prediction and prevention of the next pandemic zoonosis. Lancet 2012, 380, 1956–1965. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zappulli, V.; Ferro, S.; Bonsembiante, F.; Brocca, G.; Calore, A.; Cavicchioli, L.; Centelleghe, C.; Corazzola, G.; De Vreese, S.; Gelain, M.E.; et al. Pathology of Coronavirus Infections: A Review of Lesions in Animals in the One-Health Perspective. Animals 2020, 10, 2377. https://doi.org/10.3390/ani10122377
Zappulli V, Ferro S, Bonsembiante F, Brocca G, Calore A, Cavicchioli L, Centelleghe C, Corazzola G, De Vreese S, Gelain ME, et al. Pathology of Coronavirus Infections: A Review of Lesions in Animals in the One-Health Perspective. Animals. 2020; 10(12):2377. https://doi.org/10.3390/ani10122377
Chicago/Turabian StyleZappulli, Valentina, Silvia Ferro, Federico Bonsembiante, Ginevra Brocca, Alessandro Calore, Laura Cavicchioli, Cinzia Centelleghe, Giorgia Corazzola, Steffen De Vreese, Maria Elena Gelain, and et al. 2020. "Pathology of Coronavirus Infections: A Review of Lesions in Animals in the One-Health Perspective" Animals 10, no. 12: 2377. https://doi.org/10.3390/ani10122377
APA StyleZappulli, V., Ferro, S., Bonsembiante, F., Brocca, G., Calore, A., Cavicchioli, L., Centelleghe, C., Corazzola, G., De Vreese, S., Gelain, M. E., Mazzariol, S., Moccia, V., Rensi, N., Sammarco, A., Torrigiani, F., Verin, R., & Castagnaro, M. (2020). Pathology of Coronavirus Infections: A Review of Lesions in Animals in the One-Health Perspective. Animals, 10(12), 2377. https://doi.org/10.3390/ani10122377