Effect of Melatonin on the In Vitro Maturation of Porcine Oocytes, Development of Parthenogenetically Activated Embryos, and Expression of Genes Related to the Oocyte Developmental Capability


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Oocytes Collection and IVM
2.2. Evaluation of Cumulus Expansion, Oocyte Survival Rate, and First Polar Body Extrusion Rate
2.3. Parthenogenetically Activation
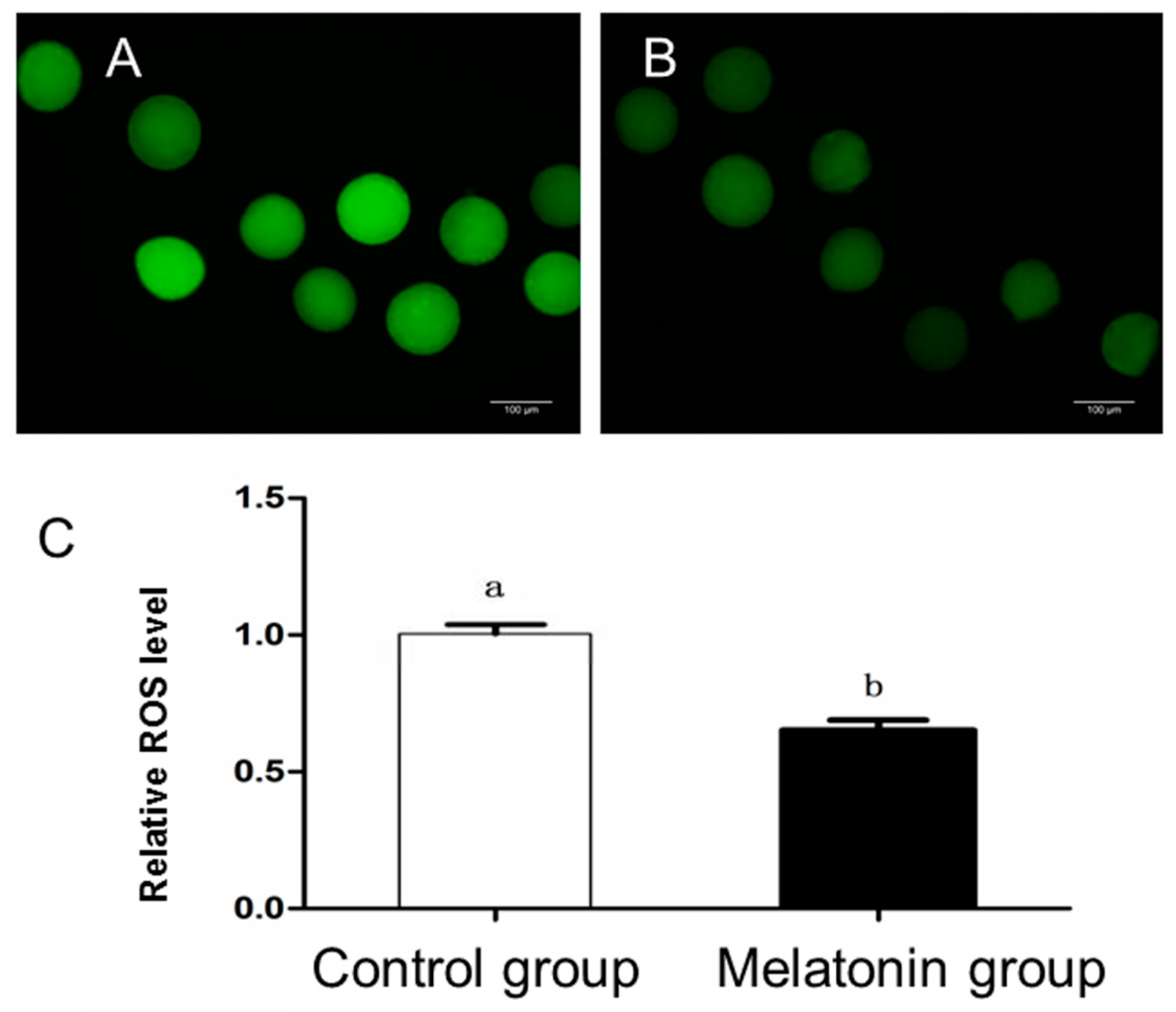
2.4. Measurement of ROS Level
2.5. Measurement of Intracellular Glutathione
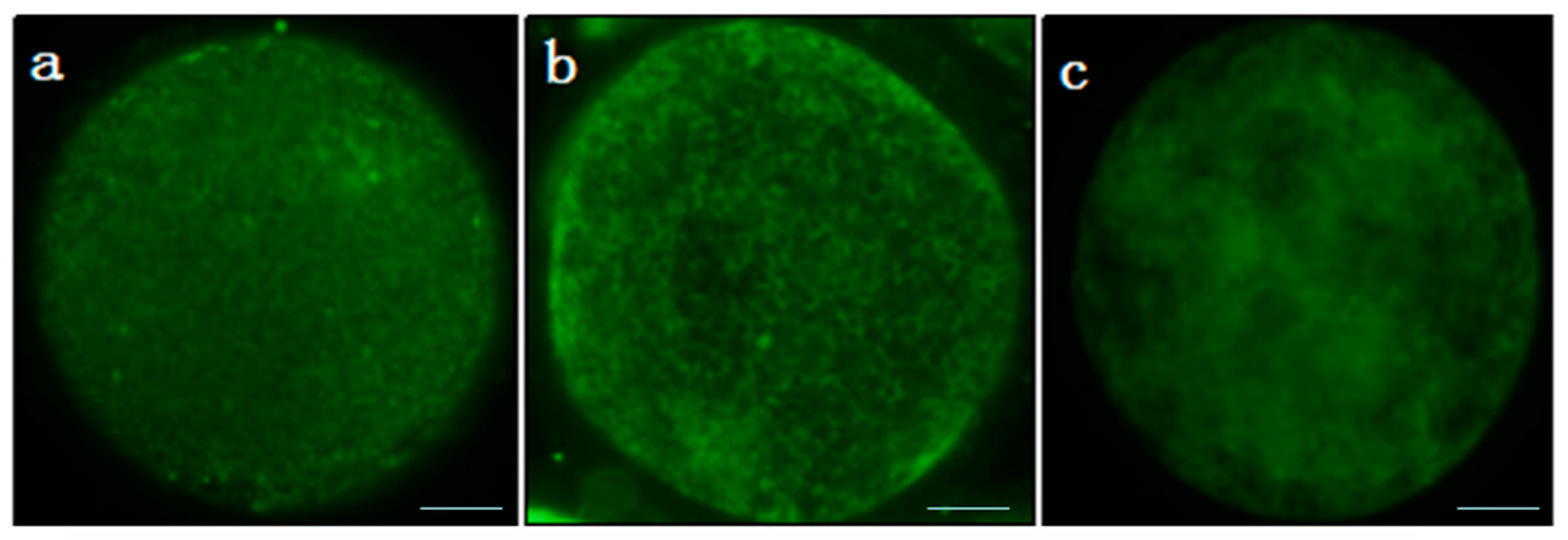
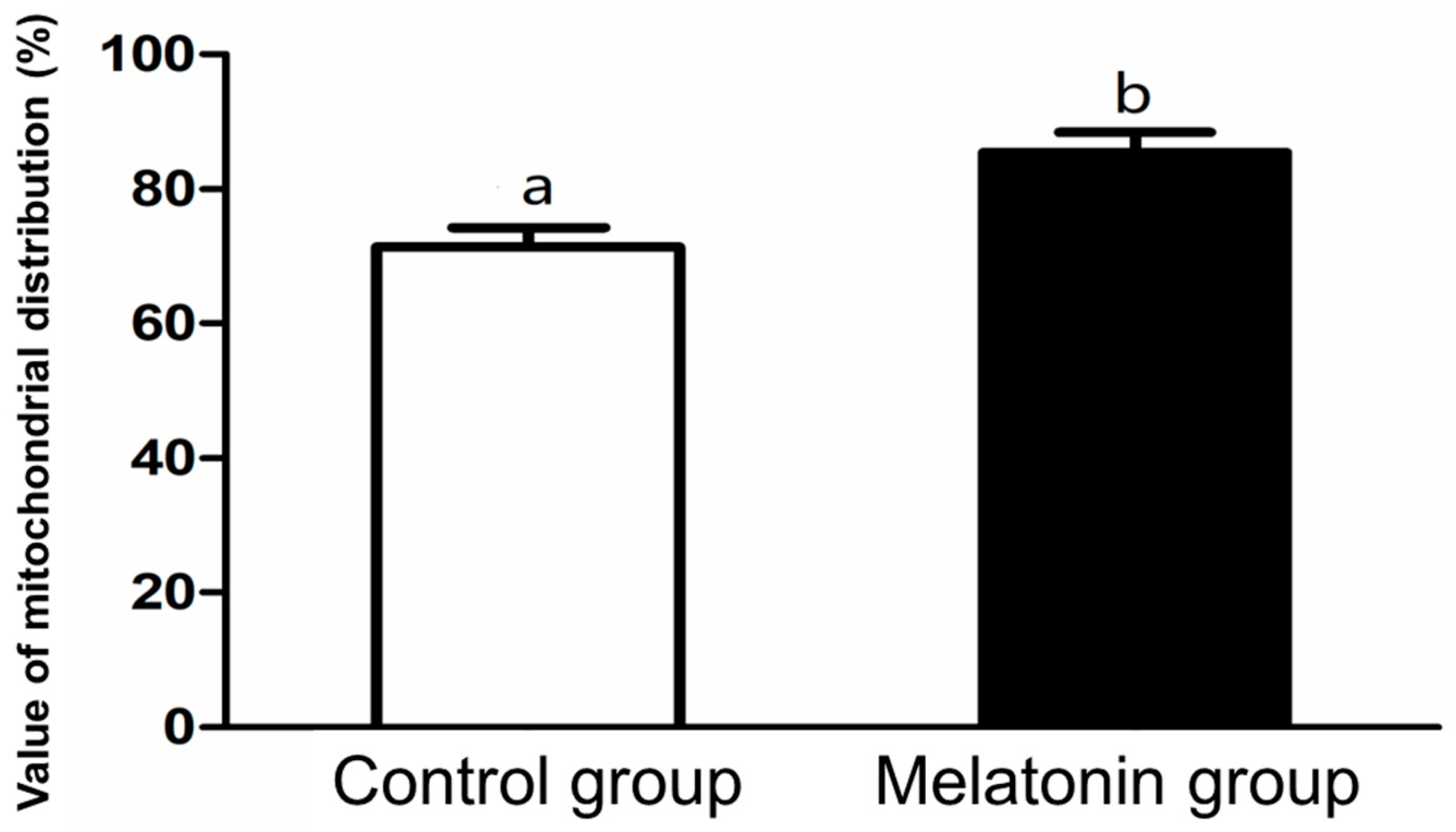
2.6. Mitochondrial Distribution Analysis
2.7. PCR Assay
2.8. Statistical Analysis
3. Results
3.1. Cumulus Expansion, Survival and First Polar Body Extrusion Rates of Oocytes, and in Vitro Development of PA Embryos in Pigs
3.2. Intracellular Levels of ROS and Glutathione; and Mitochondrial Distribution in the Oocytes
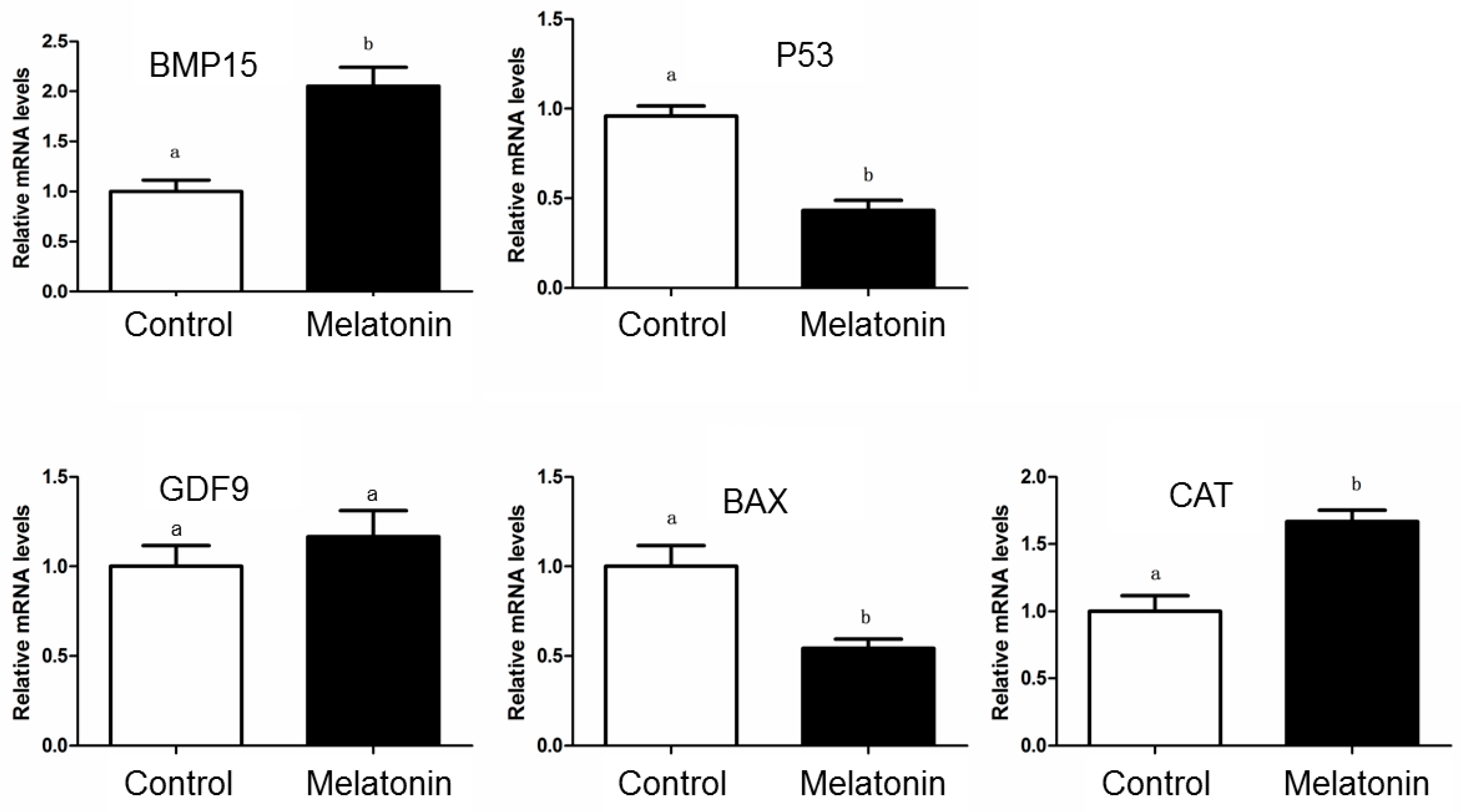
3.3. Expression of Genes Related to the Oocyte Developmental Capability
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kang, J.T.; Atikuzzaman, M.; Kwon, D.K.; Park, S.J.; Kim, S.J.; Moon, J.H.; Koo, O.J.; Jang, G.; Lee, B.C. Developmental competence of porcine oocytes after in vitro maturation and in vitro culture under different oxygen concentrations. Zygote 2012, 20, 1–8. [Google Scholar] [CrossRef]
- Guérin, P.; El Mouatassim, S.; Ménézo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Fujimoto, V.Y.; Bloom, M.S.; Huddleston, H.G.; Shelley, W.B.; Ocque, A.J.; Browne, R.W. Correlations of follicular fluid oxidative stress biomarkers and enzyme activities with embryo morphology parameters during in vitro fertilization. Fertil. Steril. 2011, 96, 1357–1361. [Google Scholar] [CrossRef]
- Hardeland, R. Antioxidative protection by melatonin: Multiplicity of mechanisms from radical detoxification to radical avoidance. Endocrine 2005, 27, 119–130. [Google Scholar] [CrossRef]
- Shi, J.M.; Tian, X.Z.; Zhou, G.B.; Wang, L.; Gao, C.; Zhu, S.E.; Zeng, S.M.; Tian, J.H.; Liu, G.S. Melatonin exists in porcine follicular fluid and improves in vitro maturation and parthenogenetic development of porcine oocytes. J. Pineal Res. 2009, 47, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.M.; Min, J.T.; Du, W.H.; Hao, H.S.; Liu, Y.; Qin, T.; Wang, D.; Zhu, H.B. Melatonin enhances the in vitro maturation and developmental potential of bovine oocytes denuded of the cumulus oophorus. Zygote 2015, 23, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; He, C.; Zhu, K.; Xu, Z.; Ma, T.; Tao, J.; Liu, G. Melatonin protects porcine oocyte in vitro maturation from heat stress. J. Pineal Res. 2015, 59, 365–375. [Google Scholar] [CrossRef]
- Cao, Z.; Gao, D.; Tong, X.; Xu, T.; Zhang, D.; Wang, Y.; Liu, Y.; Li, Y.; Zhang, Y.; Pu, Y. Melatonin improves developmental competence of oocyte-granulosa cell complexes from porcine preantral follicles. Theriogenology 2019, 133, 149–158. [Google Scholar] [CrossRef]
- Lee, S.; Jin, J.X.; Taweechaipaisankul, A.; Kim, G.A.; Lee, B.C. Synergistic effects of resveratrol and melatonin on in vitro maturation of porcine oocytes and subsequent embryo development. Theriogenology 2018, 114, 191–198. [Google Scholar] [CrossRef]
- McNatty, K.P.; Moore, L.G.; Hudson, N.L.; Quirke, L.D.; Lawrence, S.B.; Reader, K.; Hanrahan, J.P.; Smith, P.; Groome, N.P.; Laitinen, M.; et al. The oocyte and its role in regulating ovulation rate: A new paradigm in reproductive biology. Reproduction 2004, 128, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Albertini, D.F.; Nishimori, K.; Kumar, R.; Lu, N.; Matzuk, M.M. Growth differentiation factor-9 is required during early ovarian folliculogenesis. Nature 1996, 386, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Galloway, S.M.; Gregan, S.M.; Wilson, T.; McNatty, K.P.; Juengel, J.L.; Ritvos, O.; Davis, G.H. Bmp15 mutations and ovarian function. Mol. Cell Endocrinol. 2002, 191, 15–18. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Benco, A.; Tandlmajerova, A.; Vasícek, D.; Kotwica, J.; Darlak, K.; Valenzuela, F. Transcription factor p53 can regulate proliferation, apoptosis and secretory activity of luteinizing porcine ovarian granulosa cell cultured with and without ghrelin and FSH. Reproduction 2008, 136, 611–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sai, T.; Goto, Y.; Yoshioka, R.; Maeda, A.; Matsuda, F.; Sugimoto, M.; Wongpanit, K.; Jin, H.Z.; Li, J.Y.; Manabe, N. Bid and Bax are involved in granulosa cell apoptosis during follicular atresia in porcine ovaries. J. Reprod. Dev. 2011, 57, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; You, S.Y.; Cho, S.; Jeon, H.J.; Lee, S.; Cho, D.H.; Kim, J.S.; Oh, J.S. Eccentric localization of catalase to protect chromosomes from oxidative damages during meiotic maturation in mouse oocytes. Histochem. Cell Biol. 2016, 146, 281–288. [Google Scholar] [CrossRef]
- Prochazka, R.; Nemcova, L.; Nagyova, E.; Kanka, J. Expression of growth differentiation factor 9 messenger RNA in porcine growing and preovulatory ovarian follicles. Biol. Reprod. 2004, 71, 1290–1295. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, K.; Suzuki, C.; Tanaka, A.; Anas, I.M.; Iwamura, S. Birth of Piglets Derived from Porcine Zygotes Cultured in a Chemically Defined Medium. Biol. Reprod. 2002, 66, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhang, C.; Fan, X.; Li, R.; Zhang, J. Effect of C-type natriuretic peptide pretreatment on in vitro bovine oocyte maturation. In Vitro Cell. Dev. Biol. Anim. 2017, 53, 199–206. [Google Scholar] [CrossRef]
- Cui, M.S.; Fan, Y.P.; Wu, Y.; Hao, Z.D.; Liu, S.; Chen, X.J.; Zeng, S.M. Porcine cumulus cell influences ooplasmic mitochondria-lipid distributions, GSH-ATP contents and calcium release pattern after electro-activation. Theriogenology 2009, 71, 412–421. [Google Scholar] [CrossRef]
- Nagai, S.; Mabuchi, T.; Hirata, S.; Shoda, T.; Kasai, T.; Yokota, S.; Shitara, H.; Yonekawa, H.; Hoshi, K. Correlation of abnormal mitochondrial distribution in mouse oocytes with reduced developmental competence. Tohoku, J. Exp. Med. 2006, 210, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ball, G.D.; Wieben, E.D.; Byers, A.P. DNA, RNA, and protein synthesis by porcine oocyte-cumulus complexes during expansion. Biol. Reprod. 1985, 33, 739–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutin, J.A.; Audinot, V.; Ferry, G.; Delagrange, P. Molecular tools to study melatonin pathways and actions. Trends Pharmacol. Sci. 2005, 26, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zuo, X.; Li, H.; Hong, R.; Ding, B.; Liu, C.; Gao, D.; Shang, H.; Cao, Z.; Huang, W.; et al. Effects of melatonin on maturation, histone acetylation, autophagy of porcine oocytes and subsequent embryonic development. Anim. Sci. J. 2017, 88, 1298–1310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, T.; Lan, M.; Zang, X.W.; Li, Y.L.; Cui, X.S.; Kim, N.H.; Sun, S.C. Melatonin protects oocytes from MEHP exposure-induced meiosis defects in porcine. Biol. Reprod. 2018, 98, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Jin, J.X.; Taweechaipaisankul, A.; Kim, G.A.; Lee, B.C. Stimulatory Effects of Melatonin on Porcine In Vitro Maturation Are Mediated by MT2 Receptor. Int. J. Mol. Sci. 2018, 19, 1581. [Google Scholar] [CrossRef] [Green Version]
- Do, L.T.; Shibata, Y.; Taniguchi, M.; Nii, M.; Nguyen, T.V.; Tanihara, F.; Takagi, M.; Otoi, T. Melatonin Supplementation During In Vitro Maturation and Development Supports the Development of Porcine Embryos. Reprod. Domest. Anim. 2015, 50, 1054–1058. [Google Scholar] [CrossRef]
- Nakano, M.; Kato, Y.; Tsunoda, Y. Effect of melatonin treatment on the developmental potential of parthenogenetic and somatic cell nuclear-transferred porcine oocytes in vitro. Zygote 2012, 20, 199–207. [Google Scholar] [CrossRef]
- Cheng, L.; Qin, Y.; Hu, X.; Ren, L.; Zhang, C.; Wang, X.; Wang, W.; Zhang, Z.; Hao, J.; Guo, M.; et al. Melatonin protects in vitro matured porcine oocytes from toxicity of Aflatoxin B1. J. Pineal Res. 2018, 25, e12543. [Google Scholar] [CrossRef]
- Wang, F.; Tian, X.; Zhang, L.; Gao, C.; He, C.; Fu, Y.; Ji, P.; Li, Y.; Li, N.; Liu, G. Beneficial effects of melatonin on in vitro bovine embryonic development are mediated by melatonin receptor 1. J. Pineal Res. 2014, 56, 333–342. [Google Scholar] [CrossRef]
- Wang, T.; Gao, Y.Y.; Chen, L.; Nie, Z.W.; Cheng, W.; Liu, X.; Schatten, H.; Zhang, X.; Miao, Y.L. Melatonin prevents postovulatory oocyte aging and promotes subsequent embryonic development in the pig. Aging 2017, 9, 1552–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ufer, C.; Wang, C.C. The Roles of Glutathione Peroxidases during Embryo Development. Front. Mol. Neurosci. 2011, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhou, C.; Bai, Q.; Cui, Z.; ShiYang, X.; Lu, Y.; Zhang, M.; Dai, X.; Xiong, B. The protective role of melatonin in porcine oocyte meiotic failure caused by the exposure to benzo(a)pyrene. Hum. Reprod. 2018, 33, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Lee, J.E.; Kang, J.W.; Oqani, R.K.; Cho, E.S.; Kim, S.B.; Il Jin, D. Melatonin supplementation during prolonged in vitro maturation improves the quality and development of poor-quality porcine oocytes via anti-oxidative and anti-apoptotic effects. Mol. Reprod. Dev. 2018, 85, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Choe, C.; Shin, Y.W.; Kim, E.J.; Cho, S.R.; Kim, H.J.; Choi, S.H.; Han, M.H.; Han, J.; Son, D.S.; Kang, D. Synergistic effects of glutathione and β-mercaptoethanol treatment during in vitro maturation of porcine oocytes on early embryonic development in a culture system supplemented with L-cysteine. J. Reprod. Dev. 2010, 56, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, J.; Zhang, Z.; Yi, J.; He, C.; Wang, F.; Tian, X.; Yang, M.; Song, Y.; He, P.; et al. Resveratrol compares with melatonin in improving in vitro porcine oocyte maturation under heat stress. J. Anim. Sci. Biotechnol. 2016, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.M.; Wang, N.; Hao, H.S.; Li, C.Y.; Zhao, Y.H.; Yan, C.L.; Wang, H.Y.; Du, W.H.; Wang, D.; Liu, Y.; et al. Melatonin improves the fertilization capacity and developmental ability of bovine oocytes by regulating cytoplasmic maturation events. J. Pineal Res. 2018, 64, e12445. [Google Scholar] [CrossRef] [Green Version]
- Duchen, M.R. Mitochondria and calcium: From cell signalling to cell death. J. Physiol. 2000, 529, 57–68. [Google Scholar] [CrossRef]
- Schatten, H.; Sun, Q.Y.; Prather, R. The impact of mitochondrial function/dysfunction on IVF and new treatment possibilities for infertility. Reprod. Biol. Endocrinol. 2014, 12, 111. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, P.; Chabowska, A.; Malyszka, N.; Lechniak, D. Mitochondria and mitochondrial DNA in porcine oocytes and cumulus cells--A search for developmental competence marker. Mitochondrion 2016, 27, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.W.; Jiang, X.L.; Wang, Y.C.; Wang, Y.Y.; Hao, H.S.; Zhao, S.J.; Du, W.H.; Zhao, X.M.; Wang, L.; Zhu, H.B. Melatonin protects against paraquat-induced damage during in vitro maturation of bovine oocytes. J. Pineal Res. 2019, 66, e12532. [Google Scholar] [CrossRef] [PubMed]
- El-Raey, M.; Geshi, M.; Somfai, T.; Kaneda, M.; Hirako, M.; Abdel-Ghaffar, A.E.; Sosa, G.A.; El-Roos, M.E.; Nagai, T. Evidence of melatonin synthesis in the cumulus oocyte complexes and its role in enhancing oocyte maturation in vitro in cattle. Mol. Reprod. Dev. 2011, 78, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Sepasi Tehrani, H.; Moosavi-Movahedi, A.A. Catalase and its mysteries. Prog. Biophys. Mol. Biol. 2018, 140, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Li, L.; Chen, K.; Li, C.; Wang, Y.; Wang, G. Melatonin alleviates β-zearalenol and HT-2 toxin-induced apoptosis and oxidative stress in bovine ovarian granulosa cells. Environ. Toxicol. Pharmacol. 2019, 68, 52–60. [Google Scholar] [CrossRef]
- Pang, Y.W.; Sun, Y.Q.; Jiang, X.L.; Huang, Z.Q.; Zhao, S.J.; Du, W.H.; Hao, H.S.; Zhao, X.M.; Zhu, H.B. Protective effects of melatonin on bovine sperm characteristics and subsequent in vitro embryo development. Mol. Reprod. Dev. 2016, 83, 993–1002. [Google Scholar] [CrossRef]
- Tripathi, A.; PremKumar, K.V.; Pandey, A.N.; Khatun, S.; Mishra, S.K.; Shrivastav, T.G.; Chaube, S.K. Melatonin protects against clomiphene citrate-induced generation of hydrogen peroxide and morphological apoptotic changes in rat eggs. Eur. J. Pharmacol. 2011, 667, 419–424. [Google Scholar] [CrossRef]
- Paulini, F.; Melo, E.O. The role of oocyte-secreted factors GDF9 and BMP15 in follicular development and oogenesis. Reprod. Domest. Anim. 2011, 46, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Persani, L.; Rossetti, R.; Di Pasquale, E.; Cacciatore, C.; Fabre, S. The fundamental role of bone morphogenetic protein 15 in ovarian function and its involvement in female fertility disorders. Hum. Reprod. Update 2014, 20, 869–883. [Google Scholar] [CrossRef] [Green Version]
- Belli, M.; Shimasaki, S. Molecular Aspects and Clinical Relevance of GDF9 and BMP15 in Ovarian Function. Vitam. Horm. 2018, 107, 317–348. [Google Scholar]
- Sanfins, A.; Rodrigues, P.; Albertini, D.F. GDF-9 and BMP-15 direct the follicle symphony. J. Assist. Reprod. Genet. 2018, 35, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, R.Q.; Ou, S.B.; Zhang, N.F.; Ren, L.; Wei, L.N.; Zhang, Q.X.; Yang, D.Z. Increased GDF9 and BMP15 mRNA levels in cumulus granulosa cells correlate with oocyte maturation, fertilization, and embryo quality in humans. Reprod. Biol. Endocrinol. 2014, 12, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.L.; Li, Y.H.; Xu, Y.N.; Wang, Q.L.; Namgoong, S.; Cui, X.S.; Kim, N.H. Effects of growth differentiation factor 9 and bone morphogenetic protein 15 on the in vitro maturation of porcine oocytes. Reprod. Domest. Anim. 2014, 49, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Tao, J.; Chai, M.; Wu, H.; Wang, J.; Li, G.; He, C.; Xie, L.; Ji, P.; Dai, Y.; et al. Melatonin Improves the Quality of Inferior Bovine Oocytes and Promoted Their Subsequent IVF Embryo Development: Mechanisms and Results. Molecules 2017, 22, 2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Wang, F.; He, C.; Zhang, L.; Tan, D.; Reiter, R.J.; Xu, J.; Ji, P.; Liu, G. Beneficial effects of melatonin on bovine oocytes maturation: A mechanistic approach. J. Pineal Res. 2014, 57, 239–247. [Google Scholar] [CrossRef]
- Li, H.K.; Kuo, T.Y.; Yang, H.S.; Chen, L.R.; Li, S.S.; Huang, H.W. Differential gene expression of bone morphogenetic protein 15 and growth differentiation factor 9 during in vitro maturation of porcine oocytes and early embryos. Anim. Reprod. Sci. 2008, 103, 312–322. [Google Scholar] [CrossRef]
- Chipuk, J.E.; Green, D.R. Cytoplasmic p53: Bax and forward. Cell Cycle 2004, 3, 429–431. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Walls, K.C.; Ghosh, A.P.; Akhtar, R.S.; Klocke, B.J.; Roth, K.A. Cytoplasmic p53 and activated Bax regulate p53-dependent, transcription-independent neural precursor cell apoptosis. J. Histochem. Cytochem. 2010, 58, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Wang, Y.; Xing, X.; Zhang, L.; Sun, H.; Zhang, Y. Melatonin significantly improves the developmental competence of bovine somatic cell nuclear transfer embryos. J. Pineal Res. 2015, 59, 455–468. [Google Scholar] [CrossRef]
- Zhao, X.M.; Hao, H.S.; Du, W.H.; Zhao, S.J.; Wang, H.Y.; Wang, N.; Wang, D.; Liu, Y.; Qin, T.; Zhu, H.B. Melatonin inhibits apoptosis and improves the developmental potential of vitrified bovine oocytes. J. Pineal Res. 2016, 60, 132–141. [Google Scholar] [CrossRef]
- Remião, M.H.; Lucas, C.G.; Domingues, W.B.; Silveira, T.; Barther, N.N.; Komninou, E.R.; Basso, A.C.; Jornada, D.S.; Beck, R.C.; Pohlmann, A.R.; et al. Melatonin delivery by nanocapsules during in vitro bovine oocyte maturation decreased the reactive oxygen species of oocytes and embryos. Reprod. Toxicol. 2016, 63, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.W.; An, L.; Wang, P.; Yu, Y.; Yin, Q.D.; Wang, X.H.; Zhang, X.; Zhang, Q.; Yang, M.L.; Guo, M.; et al. Treatment of porcine donor cells and reconstructed embryos with the antioxidant melatonin enhances cloning efficiency. J. Pineal Res. 2013, 54, 389–397. [Google Scholar] [CrossRef] [PubMed]
- He, Y.M.; Deng, H.H.; Shi, M.H.; Bodinga, B.M.; Chen, H.L.; Han, Z.S.; Jiang, Z.L.; Li, Q.W. Melatonin modulates the functions of porcine granulosa cells via its membrane receptor MT2 in vitro. Anim. Reprod. Sci. 2016, 172, 164–172. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Product Size | GenBank Accession No. |
|---|---|---|---|
| GAPDH | F: TCAAATGGGGTGATGCTGGT R: GCAGAAGGGGCAGAGATGAT | 124 bp | XM_021091114 |
| BMP15 | F: AGCACAACCAGTCACTTTCCT R: CCCCTTGTGATTCCAGAGCT | 123 bp | NM_001005155 |
| P53 | F: AAGACCTACCCTGGCAGCTA R: ACAGCTTATTGAGGGCAGGG | 100 bp | NM_213824 |
| GDF9 | F:AGCCAGACTCCAGAGCTTTG R: TGAAGAGCCGGACAGTGTTG | 114 bp | NM_001001909.1 |
| BAX | F: GCTTCAGGGTTTCATCCAGGA R: CCAGTTCATCTCCAATGCGC | 134 bp | XM_003127290 |
| CAT | F: ACGTTGGAAAGAGGACACCC R: TCCAACGAGATCCCAATTACCA | 137 bp | NM_214301 |
| In Vitro Maturation of Oocyte and Development of Embryo | Control | Melatonin | Melatonin + Luzindole | Luzindole |
|---|---|---|---|---|
| Degree of cumulus expansion (n) | 2.74 ± 0.07 a (421) | 2.86 ± 0.08 b (385) | 2.76 ± 0.08 a (431) | 2.72 ± 0.06 a (431) |
| Survival rate % (n) | 94.23 ± 0.86 a (408/433) | 95.67 ± 0.19 a (420/439) | 93.96 ± 0.79 a (420/447) | 94.91 ± 1.62 a (410/432) |
| First polar body extrusion rate % | 79.66 ± 1.89 a (325/408) | 85.71 ± 2.26 b (360/420) | 80.48 ± 2.50 a (338/420) | 80.49 ± 1.10 a (330/410) |
| Cleavage rate % | 77.48 ± 2.05 a (117/151) | 85.63 ± 2.50 b (142/167) | 79.87 ± 1.86 a (119/149) | 76.54 ± 3.26 a (124/162) |
| Blastocyst rate % | 29.91 ± 1.75 a (35/151) | 35.92 ± 3.90 b (51/142) | 30.25 ± 1.82 a (36/119) | 29.03 ± 2.07 a (36/124) |
| Group | Number of Oocytes | Replicates | Glutathione Concentration (Pmol/Oocyte) |
|---|---|---|---|
| Control | 40 | 3 | 5.10 ± 0.13 a |
| Melatonin antagonist | 40 | 3 | 5.52 ± 0.35 a |
| Melatonin | 40 | 3 | 6.22 ± 0.21 b |
| Melatonin + receptor antagonist | 40 | 3 | 6.21 ± 0.45 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Wang, Q.; Cui, M.; Li, Q.; Mu, S.; Zhao, Z. Effect of Melatonin on the In Vitro Maturation of Porcine Oocytes, Development of Parthenogenetically Activated Embryos, and Expression of Genes Related to the Oocyte Developmental Capability. Animals 2020, 10, 209. https://doi.org/10.3390/ani10020209
Yang L, Wang Q, Cui M, Li Q, Mu S, Zhao Z. Effect of Melatonin on the In Vitro Maturation of Porcine Oocytes, Development of Parthenogenetically Activated Embryos, and Expression of Genes Related to the Oocyte Developmental Capability. Animals. 2020; 10(2):209. https://doi.org/10.3390/ani10020209
Chicago/Turabian StyleYang, Ling, Qingkai Wang, Maosheng Cui, Qianjun Li, Shuqin Mu, and Zimo Zhao. 2020. "Effect of Melatonin on the In Vitro Maturation of Porcine Oocytes, Development of Parthenogenetically Activated Embryos, and Expression of Genes Related to the Oocyte Developmental Capability" Animals 10, no. 2: 209. https://doi.org/10.3390/ani10020209
APA StyleYang, L., Wang, Q., Cui, M., Li, Q., Mu, S., & Zhao, Z. (2020). Effect of Melatonin on the In Vitro Maturation of Porcine Oocytes, Development of Parthenogenetically Activated Embryos, and Expression of Genes Related to the Oocyte Developmental Capability. Animals, 10(2), 209. https://doi.org/10.3390/ani10020209