Targeted-Release Organic Acids and Essential Oils Improve Performance and Digestive Function in Broilers under a Necrotic Enteritis Challenge

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Birds and Experimental Design
2.2.1. Trial 1
2.2.2. Trial 2
2.3. Animal Husbandry
2.4. Experimental Diets
2.5. Necrotic Enteritis Challenge Procedure
2.6. Sampling Procedure and Analyses
2.6.1. Feed
2.6.2. Bacteria Counts
2.6.3. Acute Phase Proteins
2.6.4. Histomorphological Analysis
2.6.5. Preparation of the 16S rRNA Gene Amplicon Library for MiSeq Sequencing
Library Preparation and Sequencing
Amplicon Sequences Processing and Analysis
2.7. Statistical Analysis
3. Results
3.1. First Trial
3.1.1. Growth Performance
3.1.2. Histomorphological Analysis
3.1.3. Acute Phase Proteins
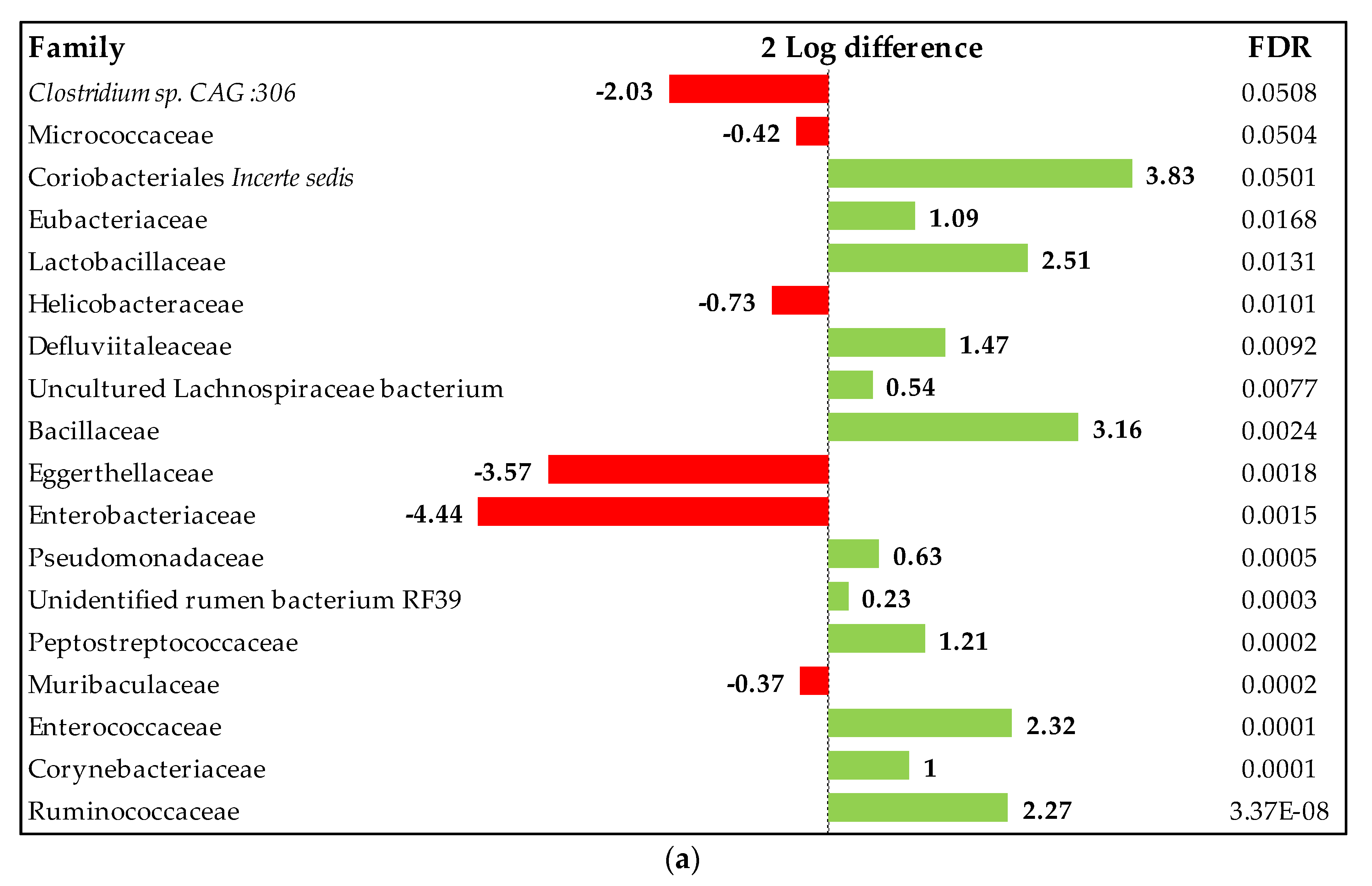
3.1.4. Ileal Microbiota Analysis
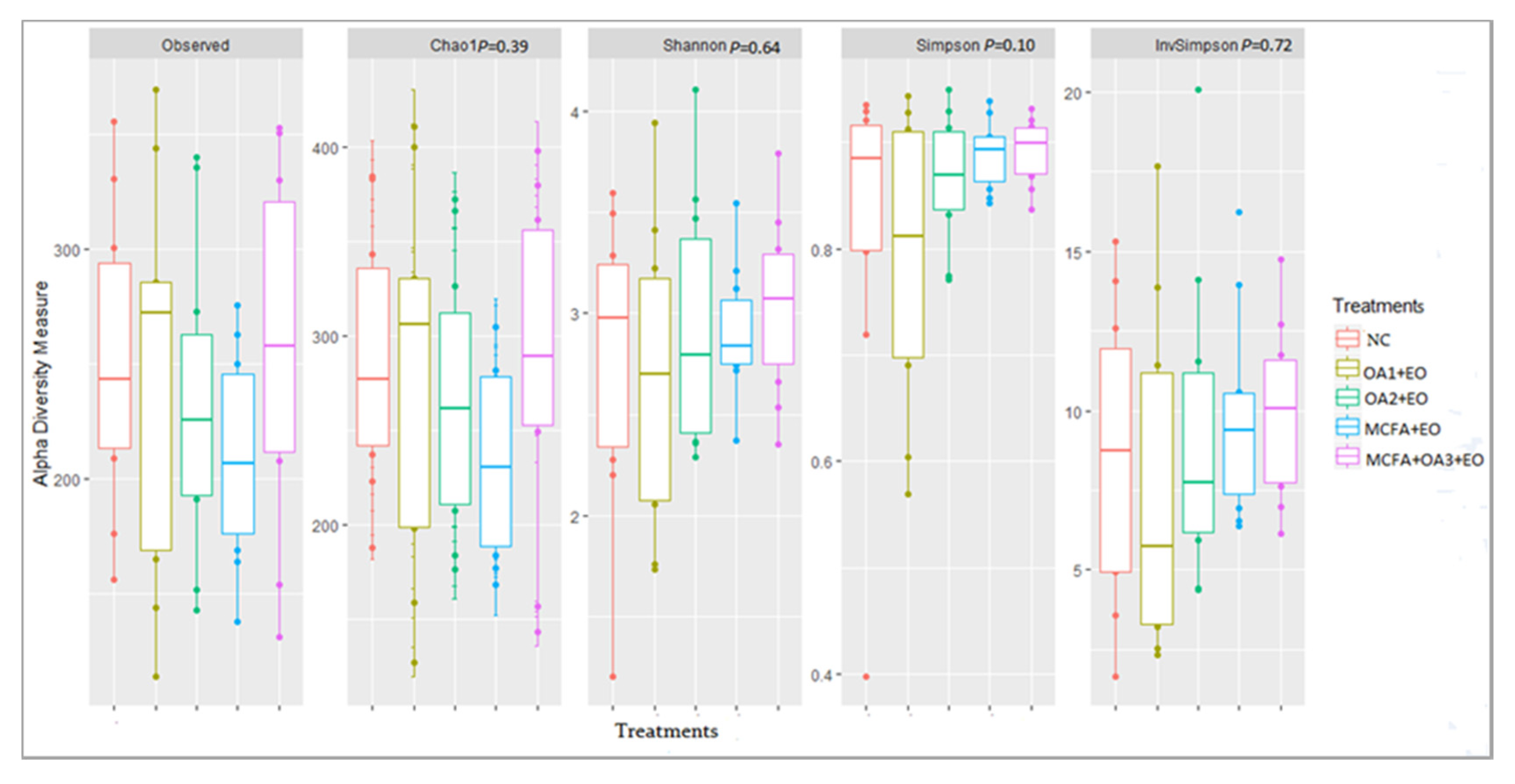
Alpha and Beta Diversity
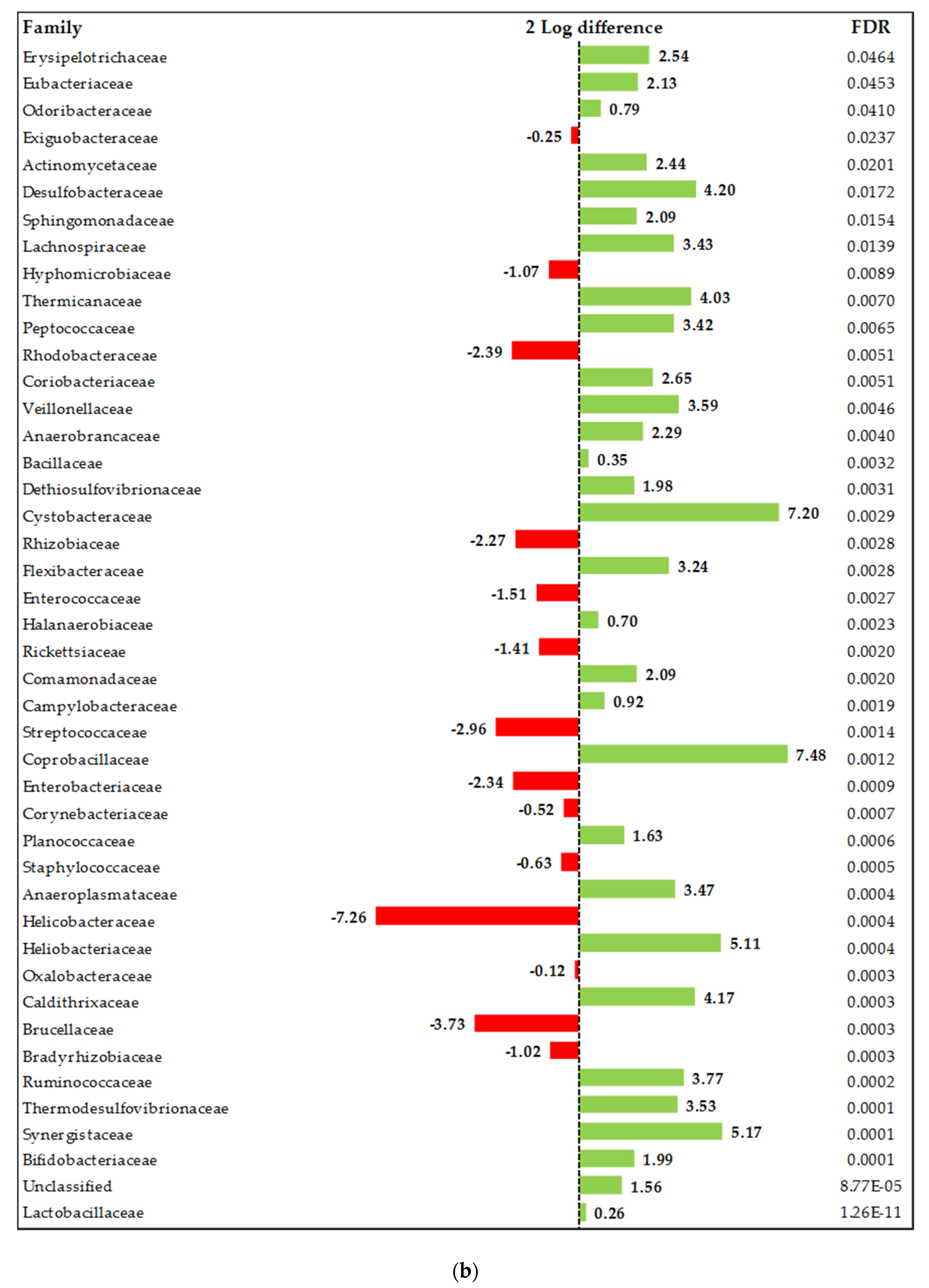
Composition of the Ileal Microbiota
3.2. Second Trial
3.2.1. Growth Performance
3.2.2. Histomorphological Analysis
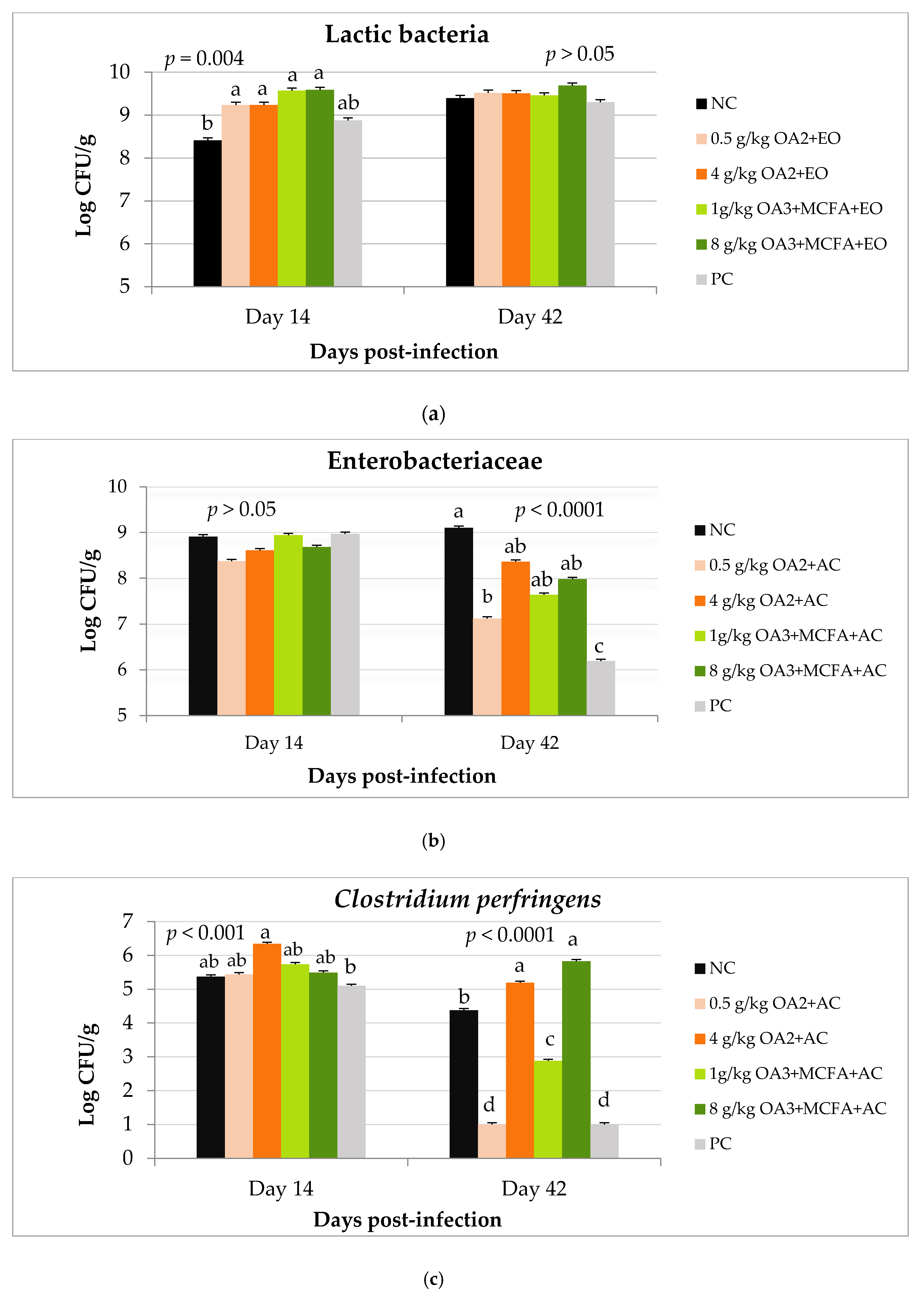
3.2.3. Bacteria Counts
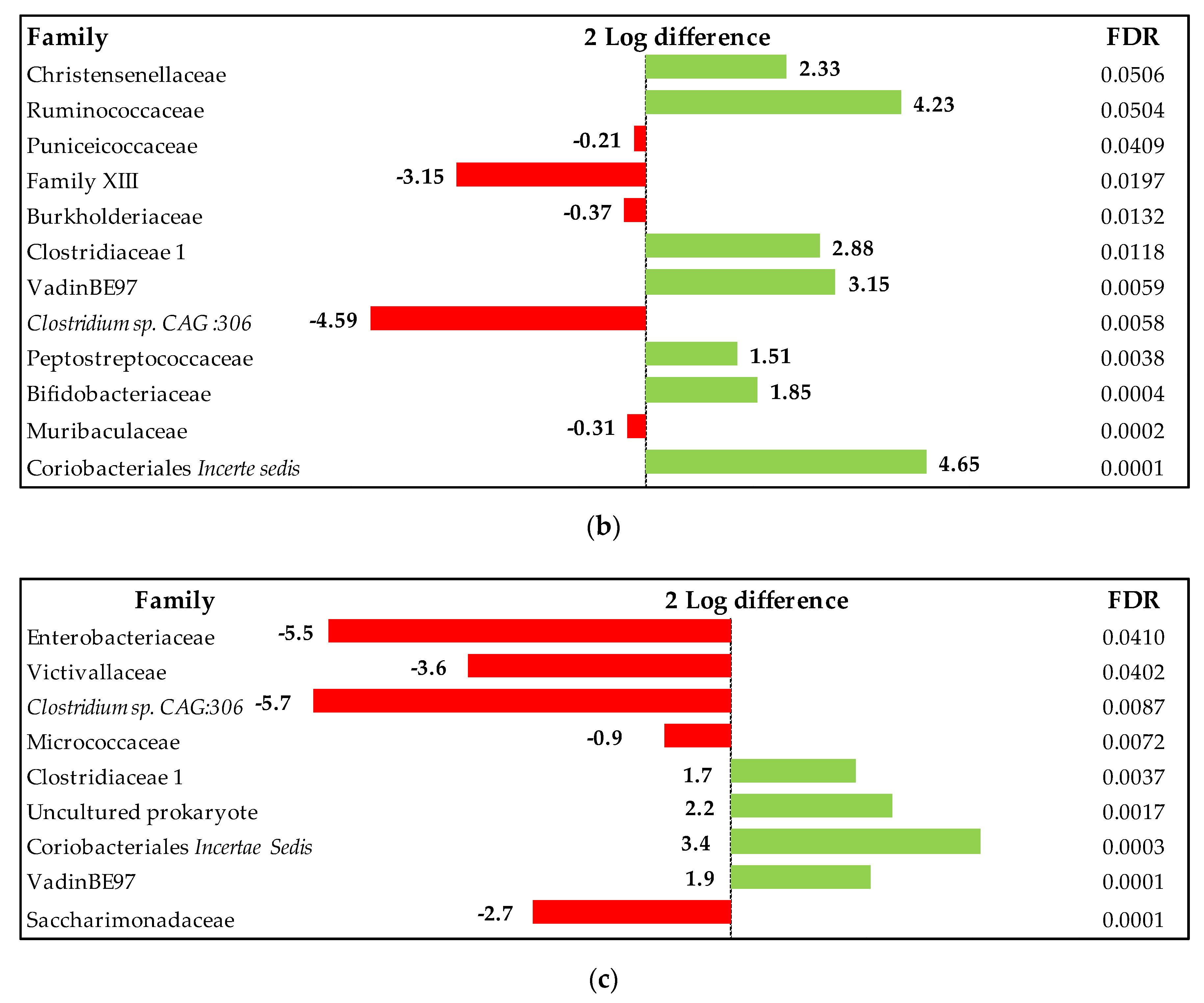
3.2.4. Cecal Microbiota Analysis
Alpha and Beta Diversity
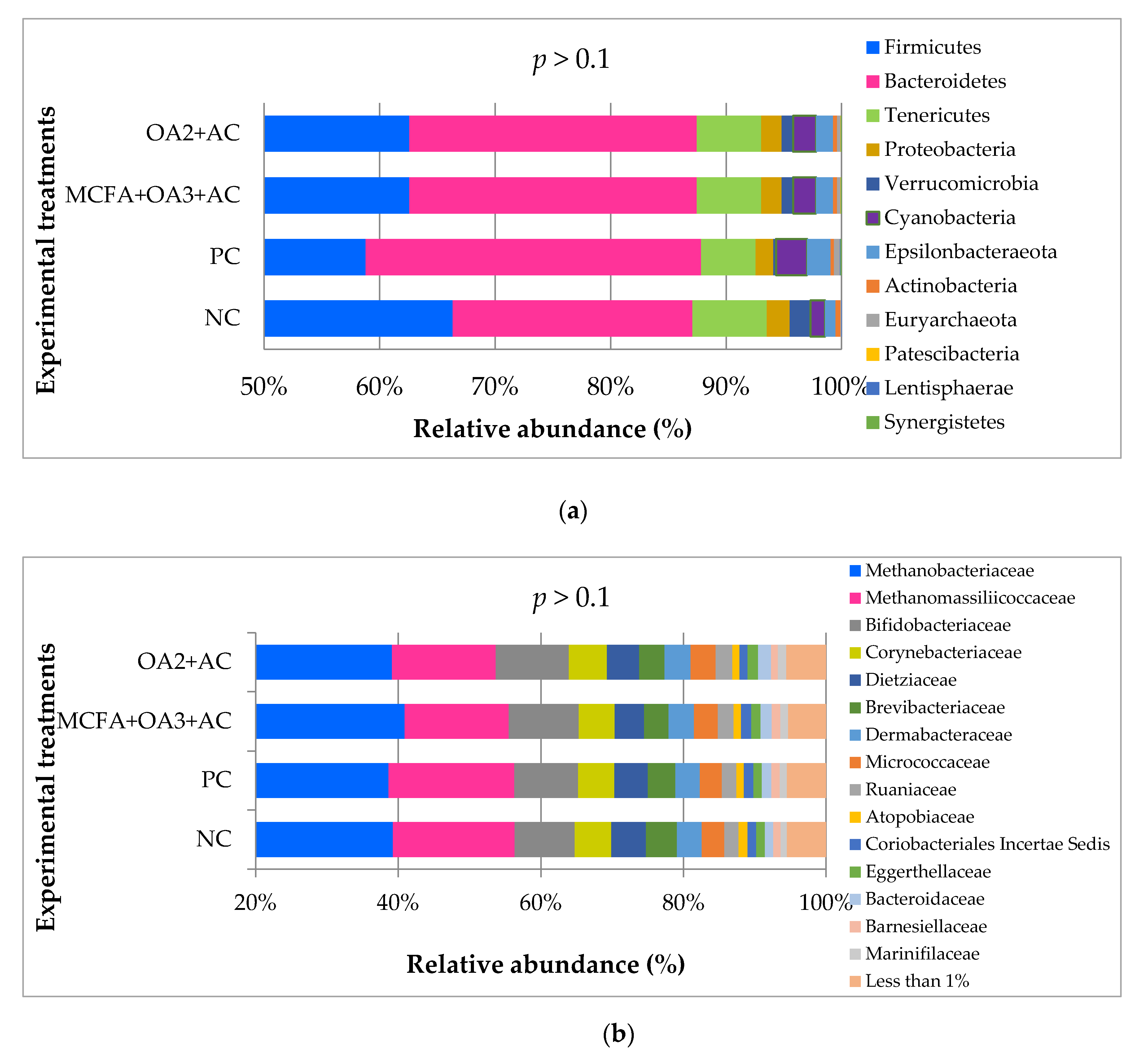
Composition of the Cecal Microbiota
4. Discussion
4.1. The Relevance of Organic Acids and Essential Oils Combination on Growth Performance
4.2. High Doses of the Additives May Become Deleterious
4.3. The Relevance of Organic Acids and Essential Oils Combination on Ileum and Caeca Microbiota
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liu, J.D.; Lumpkins, B.; Mathis, G.; Williams, S.M.; Fowler, J. Evaluation of encapsulated sodium butyrate with varying releasing times on growth performance and necrotic enteritis mitigation in broilers. Poult. Sci. 2019, 98, 1–6. [Google Scholar] [CrossRef]
- Sokale, A.O.; Menconi, A.; Mathis, G.F.; Lumpkins, B.; Sims, M.D.; Whelan, R.A.; Doranalli, K. Effect of Bacillus subtilis DSM 32315 on the intestinal structural integrity and growth performance of broiler chickens under necrotic enteritis challenge. Poult. Sci. 2019, 98, 5392–5400. [Google Scholar] [CrossRef]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The successful experimental induction of necrotic enteritis in chickens by Clostridium perfringens: A critical review. Vet. Res. 2012, 43, 1–12. [Google Scholar] [CrossRef] [Green Version]
- The True Cost of Necrotic Enteritis. Available online: https://www.poultryworld.net/Meat/Articles/2015/10/The-true-cost-of-necrotic-enteritis-2699819W/ (accessed on 18 December 2019).
- Latorre, J.D.; Adhikari, B.; Park, S.H.; Teague, K.D.; Graham, L.E.; Mahaffey, B.D.; Baxter, M.F.A.; Hernandez-Velasco, X.; Kwon, Y.M.; Ricke, S.C.; et al. Evaluation of the Epithelial Barrier Function and Ileal Microbiome in an Established Necrotic Enteritis Challenge Model in Broiler Chickens. Front. Vet. Sci. 2018, 5, 1–11. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric Pathogens and Their Toxin-Induced Disruption of the Intestinal Barrier through Alteration of Tight Junctions in Chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Kadykalo, S.; Roberts, T.; Thompson, M.; Wilson, J.; Lang, M.; Espeisse, O. The value of anticoccidials for sustainable global poultry production. Int. J. Antimicrob. Agents 2018, 51, 304–310. [Google Scholar] [CrossRef]
- Seal, B.S.; Lillehoj, H.S.; Donovan, D.M.; Gay, C.G. Alternatives to antibiotics: A symposium on the challenges and solutions for animal production. Anim. Health Res. Rev. 2013, 14, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Brenes, A.; Roura, E. Essential oils in poultry nutrition: Main effects and modes of action. Anim. Feed Sci. Technol. 2010, 158, 1–14. [Google Scholar] [CrossRef]
- Wlodarska, M.; Willing, B.P.; Bravo, D.M.; Finlay, B.B. Phytonutrient diet supplementation promotes beneficial Clostridia species and intestinal mucus secretion resulting in protection against enteric infection. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Awaad, M.H.H.; Elmenawey, M.; Ahmed, K.A. Effect of a specific combination of carvacrol, cinnamaldehyde, and Capsicum oleoresin on the growth performance, carcass quality and gut integrity of broiler chickens. Vet. World 2014, 7, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, S.; Yahyavi, M.; Zare, D.N. Influence of Dietary Organic Acids Supplementation on Reproductive Performance of Freshwater Angelfish (Pterophyllum scalare). Glob. Vet. 2014, 13, 373–377. [Google Scholar] [CrossRef]
- Dittoe, D.K.; Ricke, S.C.; Kiess, A.S. Organic acids and Potential for Modifying the Avian Gastrointestinal Tract and Reducing Pathogens and Disease. Front. Vet. Sci. 2018, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, 1–8. [Google Scholar] [CrossRef]
- Khan, S.H.; Iqbal, J. Recent advances in the role of organic acids in poultry nutrition. J. Appl. Anim. Res. 2016, 44, 359–369. [Google Scholar] [CrossRef]
- Zentek, J.; Buchheit-Renko, S.; Ferrara, F.; Vahjen, W.; Van Kessel, A.G.; Pieper, R. Nutritional and physiological role of medium-chain triglycerides and medium-chain fatty acids in piglets. Anim. Health Res. Rev. 2011, 12, 83–93. [Google Scholar] [CrossRef]
- Antibacterial Effect of MCFAs in Broilers. Available online: https://www.poultryworld.net/Health/Articles/2017/10/Antibacterial-effect-of-MCFAs-in-broilers-194522E/ (accessed on 23 January 2020).
- Giannenas, I.; Papaneophytou, C.P.; Tsalie, E.; Pappas, I.; Triantafillou, E.; Tontis, D.; Kontopidis, G.A. Dietary Supplementation of Benzoic Acid and Essential Oil Compounds Affects Buffering Capacity of the Feeds, Performance of Turkey Poults and Their Antioxidant Status, pH in the Digestive Tract, Intestinal Microbiota and Morphology. Asian Australas. J. Anim. Sci. 2014, 27, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Wang, L.; Ammeter, E.; Lahaye, L.; Liu, S.; Nyachoti, M.; Yang, C. Evaluation of lipid matrix microencapsulation for intestinal delivery of thymol in weaned pigs. Transl. Anim. Sci. 2019, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- EU. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes (Text with EEA relevance). Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar]
- Blok, M.C. CVB Table Booklet Feeding of Poultry, 2nd ed.; Product Board Animal Feed: Zoetermeer, The Netherlands, 2009; pp. 1–34. [Google Scholar]
- Cravens, R.L.; Goss, G.R.; Chi, F.; De Boer, E.D.; Davis, S.W.; Hendrix, S.M.; Richardson, J.A.; Johnston, S.L. The effects of necrotic enteritis, aflatoxin B1, and virginiamycin on growth performance, necrotic enteritis lesion scores, and mortality in young broilers. Poult. Sci. 2013, 92, 1997–2004. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemist (AOAC). Official Methods of Analysis, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Adams, M.R.; Moss, M.O. Food Microbiology, 3rd ed.; Royal Society of Chemistry: London, UK, 2007; pp. 1–459. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucl. Acids Res. 2013, 41, 1–11. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. QIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. PeerJ Prepr. 2018, 1–21. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2-Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “Vegan” Version 2.5-6. 2019. Available online: https://github.com/vegandevs/vegan (accessed on 5 February 2020).
- Paulson, J.N.; Talukder, H.; Pop, M.; Bravo, H.C. MetagenomeSeq: Statistical Analysis for Sparse High-Throughput Sequencing. Bioconductor Package: 1.12.0. 2016. Available online: http://cbcb.umd.edu/software/metagenomeSeq (accessed on 5 February 2020).
- Mehdi, Y.; Létourneau-Montminy, M.P.; Gaucher, M.L.; Chorfi, Y.; Suresh, G.; Rouissi, T.; Brar, S.K.; Côté, C.; Ramirez, A.A.; Godbout, S. Use of antibiotics in broiler production: Global impacts and alternatives. Anim. Nutr. 2018, 4, 170–178. [Google Scholar] [CrossRef]
- M’Sadeq, S.A.; Wu, S.; Swick, R.A.; Choct, M. Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide. Anim. Nutr. 2015, 1, 1–11. [Google Scholar] [CrossRef]
- Dibner, J.J.; Buttin, P. Use of Organic Acids as a Model to Study the Impact of Gut Microflora on Nutrition and Metabolism. J. Appl. Poult. Res. 2002, 11, 453–463. [Google Scholar] [CrossRef]
- Manafi, M.; Hedayati, M.; Khalaji, S.; Kamely, M. Assessment of a natural, non-antibiotic blend on performance, blood biochemistry, intestinal microflora, and morphology of broilers challenged with Escherichia coli. Rev. Bras. Zootec 2016, 45, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Suiryanrayna, M.V.A.N.; Ramana, J.V. A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, R. Intestinal health, the key to productivity: The case of organic acids. In Proceedings of the XXVII Convention ANECA-WPDC, Puerto Vallarta, Mexico, 3–6 April 2002. [Google Scholar]
- Eshak, M.G.; Elmenawey, M.A.; Atta, A.; Gharib, H.B.; Shalaby, B.; Awaad, M.H.H. The efficacy of Na-butyrate encapsulated in palm fat on performance of broilers infected with necrotic enteritis with gene expression analysis. Vet. World 2016, 9, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Basmacioǧlu Malayoǧlu, H.; Baysal, Ş.; Misirliǒlu, Z.; Polat, M.; Yilmaz, H.; Turan, N. Effects of oregano essential oil with or without feed enzymes on growth performance, digestive enzyme, nutrient digestibility, lipid metabolism and immune response of broilers fed on wheat-soybean meal diets. Br. Poult. Sci. 2010, 51, 67–80. [Google Scholar] [CrossRef]
- Zhou, F.; Ji, B.; Zhang, H.; Jiang, H.; Yang, Z.; Li, J.; Li, J.; Ren, Y.; Yan, W. Synergistic effect of thymol and carvacrol combined with chelators and organic acids against Salmonella Typhimurium. J. Food Prot. 2007, 70, 1704–1709. [Google Scholar] [CrossRef]
- Bozkurt, M.; Küçükyilmaz, K.; Çatli, A.U.; Çinar, M.; Çabuk, M.; Alçiçek, A. Effects of administering an essential oil mixture and an organic acid blend separately and combined to diets on broiler performance. Arch. Geflugelkd. 2012, 76, 81–87. [Google Scholar]
- Gao, Y.Y.; Zhang, X.L.; Xu, L.-H.; Peng, H.; Wang, C.-K.; Bi, Y.-Z. Encapsulated blends of essential oils and organic acids improved performance, intestinal morphology, cecal microflora, and jejunal enzyme activity of broilers. Czech J. Anim. Sci. 2019, 64, 189–198. [Google Scholar] [CrossRef]
- Sabour, S.; Tabeidian, S.A.; Sadeghi, G. Dietary organic acid and fiber sources affect performance, intestinal morphology, immune responses and gut microflora in broilers. Anim. Nutr. 2019, 5, 156–162. [Google Scholar] [CrossRef]
- Yang, X.; Xin, H.; Yang, C.; Yang, X. Impact of essential oils and organic acids on the growth performance, digestive functions and immunity of broiler chickens. Anim. Nutr. 2018, 4, 388–393. [Google Scholar] [CrossRef]
- Wang, H.; Liang, S.; Li, X.; Yang, X.; Long, F.; Yang, X. Effects of encapsulated essential oils and organic acids on laying performance, egg quality, intestinal morphology, barrier function, and microflora count of hens during the early laying period. Poult. Sci. 2019, 98, 2751–2760. [Google Scholar] [CrossRef]
- Sun, L.; Dong, H.; Zhang, Z.; Liu, J.; Hu, Y.; Ni, Y.; Grossmann, R.; Zhao, R. Activation of epithelial proliferation induced by Eimeria acervulina infection in the duodenum may be associated with cholesterol metabolism. Oncotarget 2016, 7, 27627–27640. [Google Scholar] [CrossRef] [Green Version]
- Jerzsele, A.; Szeker, K.; Csizinszky, R.; Gere, E.; Jakab, C.; Mallo, J.J.; Galfi, P. Efficacy of protected sodium butyrate, a protected blend of essential oils, their combination, and Bacillus amyloliquefaciens spore suspension against artificially induced necrotic enteritis in broilers. Poult. Sci. 2012, 91, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Stefanello, C.; Rosa, D.P.; Dalmoro, Y.K.; Segatto, A.L.; Vieira, M.S.; Moraes, M.L.; Santin, E. Protected Blend of Organic Acids and Essential Oils Improves Growth Performance, Nutrient Digestibility, and Intestinal Health of Broiler Chickens Undergoing an Intestinal Challenge. Front. Vet. Sci. 2020, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Patlan, D.; Sol, B.; Pontin, K.P.; Latorre, J.D.; Baxter, M.F.A.; Hernandez-Velasco, X.; Merino-Guzman, R.; Abraham, M.; Hargis, B.M.; Lopez-Arellano, R.; et al. Evaluation of the Dietary Supplementation of a Formulation Containing Ascorbic Acid and a Solid Dispersion of Curcumin with Boric Acid against Broiler Chickens. Animals 2019, 9, 184. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.Y.; Yan, F.; Keen, C.A.; Waldroup, P. Evaluation of Microencapsulated Essential Oils and Organic Acids in Diets for Broiler Chickens. Int. J. Poult. Sci. 2005, 4, 612–619. [Google Scholar]
- Lin, J.; Nafday, S.M.; Chauvin, S.N.; Magid, M.S.; Pabbatireddy, S.; Holzman, I.R.; Babyatsky, M.W. Variable Effects of Short Chain Fatty Acids and Lactic Acid in Inducing Intestinal Mucosal Injury in Newborn Rats. J. Pediatr. Gastroenterol. Nutr. 2002, 35, 545–550. [Google Scholar] [CrossRef]
- Timbermont, L.; Lanckriet, A.; Dewulf, J.; Nollet, N.; Schwarzer, K.; Haesebrouck, F.; Ducatelle, R.; van Immerseel, F. Control of clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils. Avian Pathol. 2010, 39, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Barcelo, A.; Claustre, J.; Moro, F.; Chayvialle, J.A.; Cuber, J.C.; Plaisancié, P. Mucin secretion is modulated by luminal factors in the isolated vascularly perfused rat colon. Gut 2000, 46, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of butyrate on intestinal barrier function in a caco-2 cell monolayer model of intestinal barrier. Pediatr. Res. 2007, 61, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. J. Anim. Sci. Biotechnol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Iacob, S.; Iacob, D.G.; Luminos, L.M. Intestinal Microbiota as a Host Defense Mechanism to Infectious Threats. Front. Microbiol. 2019, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bedford, A.; Gong, J. Implications of butyrate and its derivatives for gut health and animal production. Anim. Nutr. 2018, 4, 151–159. [Google Scholar] [CrossRef]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I.; Anaerostipes, L.; Faecalibacterium, F. Chicken Cecal Microbiota. Appl. Environ. Microbiol. 2016, 82, 1569–1576. [Google Scholar] [CrossRef] [Green Version]
- Eeckhaut, V.; van Immerseel, F.; Croubels, S.; de Baere, S.; Haesebrouck, F.; Ducatelle, R.; Louis, P.; Vandamme, P. Butyrate production in phylogenetically diverse Firmicutes isolated from the chicken caecum. Microb. Biotechnol. 2011, 4, 503–512. [Google Scholar] [CrossRef] [Green Version]
- De Maesschalck, C.; Van Immerseel, F.; Eeckhaut, V.; De Baere, S.D.; Cnockaert, M.; Croubels, S.; Haesebrouck, F.; Ducatelle, R.; Vandamme, P. Faecalicoccus acidiformans gen. nov., Sp. nov., Isolated from the chicken caecum, And reclassification of streptococcus pleomorphus (barnes et al. 1977), Eubacterium biforme (eggerth 1935) and eubacterium cylindroides (cato et al. 1974) as faecalicoccus p. Int. J. Syst. Evol. Microbiol. 2014, 64, 3877–3884. [Google Scholar] [CrossRef] [Green Version]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Lyons, A.; O’Mahony, D.; O’Brien, F.; MacSharry, J.; Sheil, B.; Ceddia, M.; Russell, W.M.; Forsythe, P.; Bienenstock, J.; Kiely, B.; et al. Bacterial strain-specific induction of Foxp3+ T regulatory cells is protective in murine allergy models. Clin. Exp. Allergy 2010, 40, 811–819. [Google Scholar] [CrossRef]
- O’Mahony, C.; Scully, P.; O’Mahony, D.; Murphy, S.; O’Brien, F.; Lyons, A.; Sherlock, G.; MacSharry, J.; Kiely, B.; Shanahan, F.; et al. Commensal-induced regulatory T cells mediate protection against pathogen-stimulated NF-κB activation. PLoS Pathog. 2008, 4, e1000112. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Lin, Y.; Zeng, D.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K.; Jing, B.; et al. Bacillus licheniformis normalize the ileum microbiota of chickens infected with necrotic enteritis. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Shang, Y.; Kumar, S.; Thippareddi, H.; Kim, W.K. Effect of dietary fructooligosaccharide (FOS) supplementation on ileal microbiota in broiler chickens. Poult. Sci. 2018, 97, 1–13. [Google Scholar] [CrossRef]
- Ceelen, L.M.; Decostere, A.; Chiers, K.; Ducatelle, R.; Maes, D.; Haesebrouck, F. Pathogenesis of Helicobacter pullorum infections in broilers. Int. J. Food Microbiol. 2007, 116, 207–213. [Google Scholar] [CrossRef]
- Javed, S.; Gul, F.; Javed, K.; Bokhari, H. Helicobacter pullorum: An emerging zoonotic pathogen. Front. Microbiol. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.M.; Shah, T.; Deshpande, S.; Jakhesara, S.J.; Koringa, P.G.; Rank, D.N.; Joshi, C.G. High through put 16S rRNA gene-based pyrosequencing analysis of the fecal microbiota of high FCR and low FCR broiler growers. Mol. Biol. Rep. 2012, 39, 10595–10602. [Google Scholar] [CrossRef] [PubMed]
- Crisol-Martínez, E.; Stanley, D.; Geier, M.S.; Hughes, R.J.; Moore, R.J. Sorghum and wheat differentially affect caecal microbiota and associated performance characteristics of meat chickens. PeerJ 2017, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, P.; Liu, P.; Song, P.; Chen, X.; Ma, X. Moderate dietary protein restriction alters the composition of gut microbiota and improves ileal barrier function in adult pig model OPEN. Sci. Rep. 2017, 7, 43412. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Starter | Growing | Finishing |
|---|---|---|---|
| Ingredient composition (%) | |||
| Maize | 41.20 | 40.30 | 40.50 |
| Wheat | 15.00 | 20.00 | 25.00 |
| Soybean meal 48 | 28.80 | 32.60 | 25.50 |
| L-lysine HCL | 0.15 | 0.07 | 0.05 |
| DL-methionine | 0.23 | 0.13 | 0.14 |
| L-threonine | - | - | 0.05 |
| Soy oil | 1.10 | 1.40 | - |
| Palm oil | - | 2.50 | 6.00 |
| Extruded soybean meal | 10.00 | - | - |
| Limestone | 1.09 | 0.70 | 0.70 |
| Dicalcium phosphate | 1.49 | 1.44 | 1.08 |
| Salt | 0.20 | 0.20 | 0.20 |
| Vitamin-mineral premix * | 0.40 | 0.40 | 0.40 |
| Sodium bicarbonate | 0.34 | 0.26 | 0.27 |
| Calculated composition (%) | |||
| Dry matter | 88.1 | 88.1 | 88.4 |
| G.E (kcal/kg) | 3009 | 3101 | 3249 |
| Crude protein | 22.0 | 21.0 | 18.0 |
| Lysine | 1.35 | 1.18 | 1.06 |
| Methionine | 0.58 | 0.46 | 0.43 |
| Ca | 0.95 | 0.78 | 0.67 |
| Total P | 0.64 | 0.62 | 0.53 |
| Available P | 0.45 | 0.44 | 0.37 |
| Analyzed composition (%) | |||
| Dry matter | 90.2 | 90.4 | 90.6 |
| GE, kcal/kg | 4081 | 4332 | 4395 |
| Crude protein | 22.5 | 21.3 | 18.5 |
| Ether extract | 4.8 | 6.0 | 7.9 |
| Crude fiber | 3.7 | 4.7 | 3.8 |
| Items | Experimental Treatments | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| NC | OA1 + AC | OA2 + AC | MCFA + AC | MCFA + OA3 + AC | |||
| BW (g) | |||||||
| d0 | 42.8 | 42.7 | 42.8 | 42.7 | 42.9 | 0.05 | 0.4008 |
| d10 | 228.4 c | 240.5 a,b | 229.3 b,c | 242.6 a | 237.9 a,b,c | 2.84 | 0.0026 |
| d28 | 1020 y | 1093 x | 1101 x | 1100 x | 1141 x | 28.8 | 0.0803 |
| d41 | 1942 b | 2188 a | 2285 a | 2085 a,b | 2203 a | 51.3 | 0.0005 |
| ADG (g/d) | |||||||
| d0–10 | 18.6 c | 19.8 a,b | 18.6 b,c | 20.0 a | 19.5 a,b,c | 0.28 | 0.0022 |
| d11–28 | 44.0 | 47.4 | 48.4 | 47.6 | 50.1 | 1.70 | 0.1116 |
| d29–41 | 70.9 | 84.3 a,b | 91.0 a | 75.8 b,c | 81.7 a,b | 2.74 | <0.0001 |
| d0–41 | 46.3 b | 52.3 a | 54.6 a | 49.8 a,b | 52.7 a | 1.268 | <0.0001 |
| ADFI (g/d) | |||||||
| d0–10 | 32.0 x | 31.1 x,y | 31.1 x,y | 30.5 y | 32.0 x,y | 0.41 | 0.0721 |
| d11–28 | 77.5 | 82.8 | 84.0 | 80.5 | 82.9 | 1.78 | 0.1248 |
| d29–41 | 162.3 a,b | 167.2 a,b | 173.9 a | 158.4 b,c | 170.1 a,b | 3.57 | <0.0001 |
| d0–41 | 88.5 b | 96.9 a,b | 99.6 a | 93.0 a,b | 98.1a,b | 1.93 | 0.0248 |
| FCR | |||||||
| d0–10 | 1.72 a | 1.57 a,b | 1.67 a,b | 1.53 b | 1.64 a,b | 0.032 | 0.0051 |
| d11–28 | 1.82 | 1.77 | 1.76 | 1.69 | 1.70 | 0.035 | 0.1203 |
| d29–41 | 2.29 a | 1.98 b | 1.91 b | 2.01 a,b | 2.08 a,b | 0.042 | 0.0007 |
| d0–41 | 1.87 a | 1.78 b | 1.78 b | 1.75 b | 1.79 b | 0.034 | <0.0001 |
| Items | Experimental Treatments | Statistics | |||||
|---|---|---|---|---|---|---|---|
| NC | OA1 + AC | OA2 + AC | MCFA + AC | MCFA + OA3 + AC | SEM | p-Value | |
| Villus height (μm) | 828.5 c | 1044.0 a,b | 1088.4 a | 925.1 b,c | 1054.6 a,b | 35.90 | <0.0001 |
| Crypt depth (μm) | 219.6 a | 174.5 b | 179.2 b | 191.5 b | 182.5 b | 4.90 | <0.0001 |
| VH:CD ratio | 3.79 c | 6.04 a | 6.10a | 4.89 b | 5.82 a | 0.22 | <0.0001 |
| Goblet cells density/100 μm of villus height | 15.8 | 13.1 | 11.4 | 13.5 | 13.9 | 1.07 | 0.6134 |
| Intraepithelial lymphocyte density/100 μm of villus height | 7.9 | 6.6 | 6.0 | 7.1 | 6.8 | 0.37 | 0.6424 |
| Items | Experimental Treatments | p-Value | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NC | OA2 + AC (A) | OA3 + MCFA + AC (B) | SEM | |||||||||||
| 0.5 g/kg | 1 g/kg | 2 g/kg | 4 g/kg | 1 g/kg | 2 g/kg | 4 g/kg | 8 g/kg | Linear | Quadratic | |||||
| A | B | A | B | |||||||||||
| BW (g) | ||||||||||||||
| d0 | 39.3 | 39.4 | 39.4 | 39.4 | 39.4 | 39.4 | 39.4 | 39.4 | 39.4 | 0.04 | 0.85 | 0.85 | 0.22 | 0.4 |
| d10 | 228.5 | 246.3 | 234.6 | 234.3 | 227.2 | 240.5 | 249 | 243.2 | 239.8 | 5.01 | 0.15 | 0.42 | 0.23 | 0.03 |
| d28 | 1132 | 1283 | 1255 | 1241 | 1203 | 1194 | 1296 | 1263 | 1247 | 23.1 | 0.78 | 0.01 | 0.001 | 0.001 |
| d42 | 2369 | 2598 | 2561 | 2573 | 2327 | 2469 | 2549 | 2549 | 2527 | 42.8 | 0.007 | 0.06 | <0.0001 | 0.02 |
| ADG (g/d) | ||||||||||||||
| d0–10 | 18.9 | 20.7 | 19.5 | 19.5 | 18.8 | 20.1 | 21.0 | 20.4 | 20.0 | 0.46 | 0.13 | 0.34 | 0.41 | 0.03 |
| d0–42 | 55.5 | 60.9 | 60.0 | 60.3 | 54.5 | 57.8 | 59.8 | 59.8 | 59.2 | 1.07 | 0.006 | 0.004 | <0.0001 | 0.11 |
| ADFI (g/d) | ||||||||||||||
| d0–10 | 26.8 | 27.5 | 26.2 | 27.8 | 26.9 | 26.7 | 28.9 | 27.9 | 27.6 | 0.78 | 0.88 | 0.56 | 0.66 | 0.17 |
| d0–42 | 95.1 | 102.6 | 101.4 | 100.6 | 95.2 | 98.3 | 102.1 | 99.9 | 101.1 | 1.46 | 0.08 | 0.05 | 0.0002 | 0.1 |
| FCR | ||||||||||||||
| d0–10 | 1.42 | 1.33 | 1.34 | 1.43 | 1.43 | 1.33 | 1.38 | 1.37 | 1.38 | 0.042 | 0.1 | 0.29 | 0.47 | 0.73 |
| d0–42 | 1.71 | 1.68 | 1.69 | 1.67 | 1.75 | 1.70 | 1.71 | 1.68 | 1.71 | 0.018 | 0.13 | 0.01 | 0.42 | 0.96 |
| Items | Experimental Treatments | p-Value | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NC | OA2 + AC (A) | OA3 + MCFA + AC (B) | SEM | |||||||||||
| 0.5 g/kg | 1 g/kg | 2 g/kg | 4 g/kg | 1 g/kg | 2 g/kg | 4 g/kg | 8 g/kg | Linear | Quadratic | |||||
| A | B | A | B | |||||||||||
| Villus height, VH (μm) | 803.7 | 872.1 | 895.0 | 873.7 | 773.5 | 909.9 | 847.4 | 843.6 | 850.8 | 36.15 | 0.20 | 0.92 | 0.29 | 0.56 |
| Crypt depth, CD(μm) | 160.9 | 149.5 | 148.8 | 146.0 | 181.9 | 148.1 | 147.1 | 158.2 | 152.2 | 6.28 | 0.002 | 0.85 | <0.0001 | 0.69 |
| Ratio VH:CD | 5.0 | 5.8 | 6.1 | 6.0 | 4.3 | 6.2 | 5.8 | 5.4 | 5.7 | 0.27 | 0.002 | 0.81 | <0.0001 | 0.46 |
| Goblet cells Density/100 μm of villus height | 21.7 | 22.2 | 21.1 | 20.8 | 29.1 | 19.5 | 19.3 | 22.6 | 23.2 | 1.53 | 0.003 | 0.17 | 0.05 | 0.55 |
| Index | NC | PC | OA2 + AC | OA3 + MCFA + AC | SEM | p-Value |
|---|---|---|---|---|---|---|
| Shannon | 0.91 b | 0.93 a,b | 0.94 a | 0.95 a | 0.006 | 0.009 |
| Simpson | 3.01 b | 3.14 a,b | 3.31 a | 3.36 a | 0.074 | 0.008 |
| Invsimpson | 12.43 b | 15.60 a,b | 19.26 a | 19.82 a | 1.502 | 0.004 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelli, N.; Pérez, J.F.; Vilarrasa, E.; Cabeza Luna, I.; Melo-Duran, D.; D’Angelo, M.; Solà-Oriol, D. Targeted-Release Organic Acids and Essential Oils Improve Performance and Digestive Function in Broilers under a Necrotic Enteritis Challenge. Animals 2020, 10, 259. https://doi.org/10.3390/ani10020259
Abdelli N, Pérez JF, Vilarrasa E, Cabeza Luna I, Melo-Duran D, D’Angelo M, Solà-Oriol D. Targeted-Release Organic Acids and Essential Oils Improve Performance and Digestive Function in Broilers under a Necrotic Enteritis Challenge. Animals. 2020; 10(2):259. https://doi.org/10.3390/ani10020259
Chicago/Turabian StyleAbdelli, Nedra, José Francisco Pérez, Ester Vilarrasa, Irene Cabeza Luna, Diego Melo-Duran, Matilde D’Angelo, and David Solà-Oriol. 2020. "Targeted-Release Organic Acids and Essential Oils Improve Performance and Digestive Function in Broilers under a Necrotic Enteritis Challenge" Animals 10, no. 2: 259. https://doi.org/10.3390/ani10020259
APA StyleAbdelli, N., Pérez, J. F., Vilarrasa, E., Cabeza Luna, I., Melo-Duran, D., D’Angelo, M., & Solà-Oriol, D. (2020). Targeted-Release Organic Acids and Essential Oils Improve Performance and Digestive Function in Broilers under a Necrotic Enteritis Challenge. Animals, 10(2), 259. https://doi.org/10.3390/ani10020259