Model-Based Distribution and Abundance of Three Delphinidae in the Mediterranean


 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
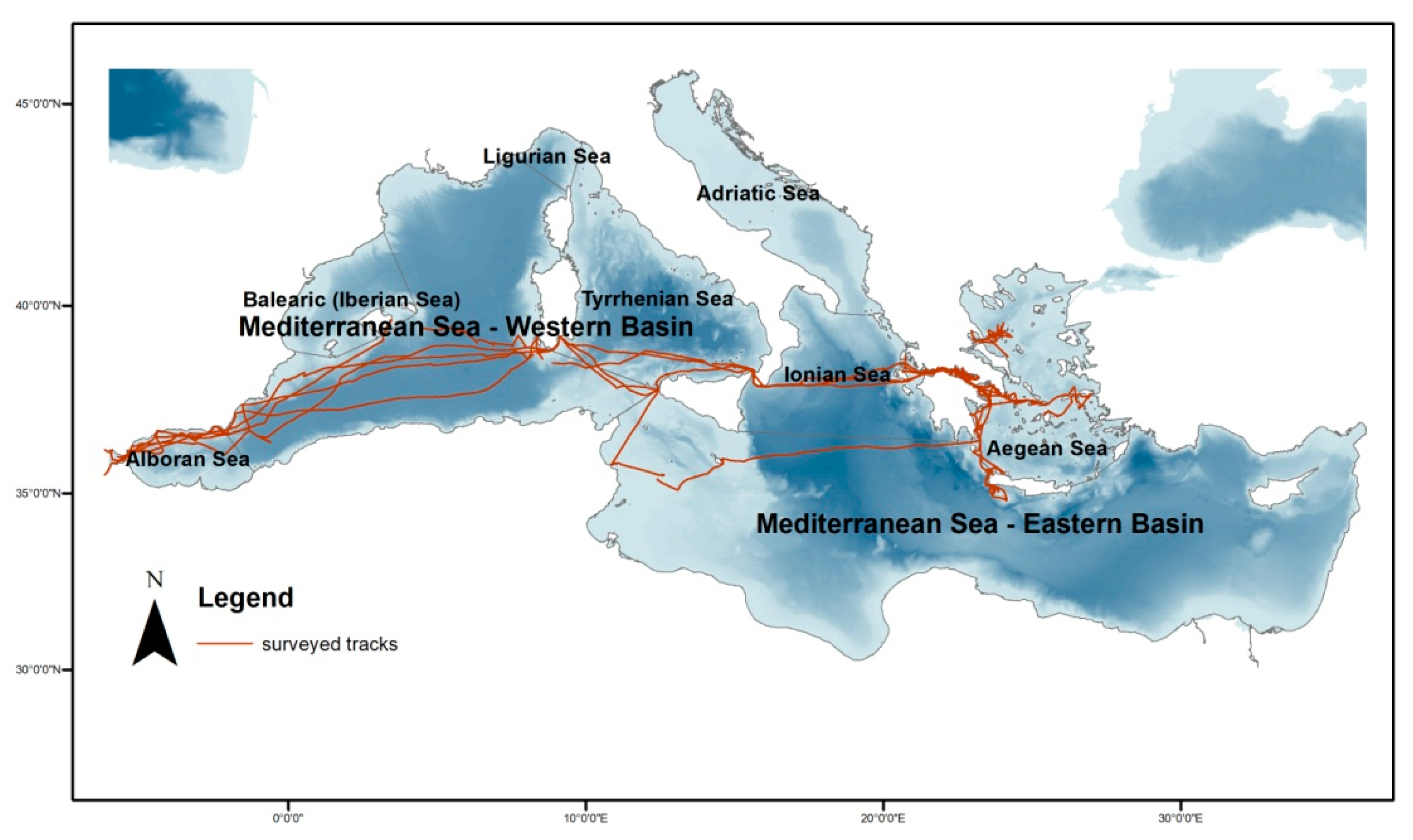
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis and Modeling Framework
3. Results
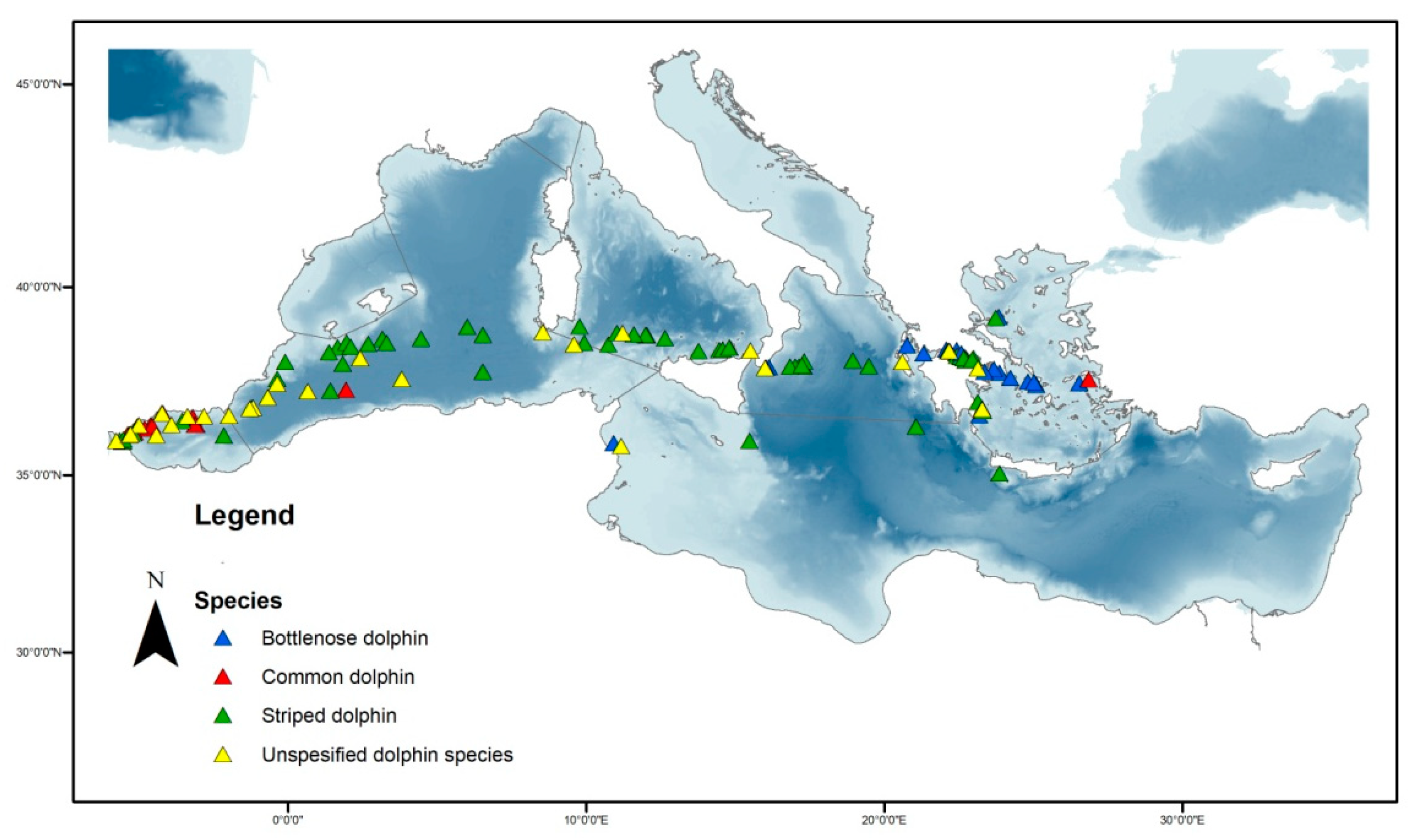
3.1. Three Main Dolphin Species Occurrence
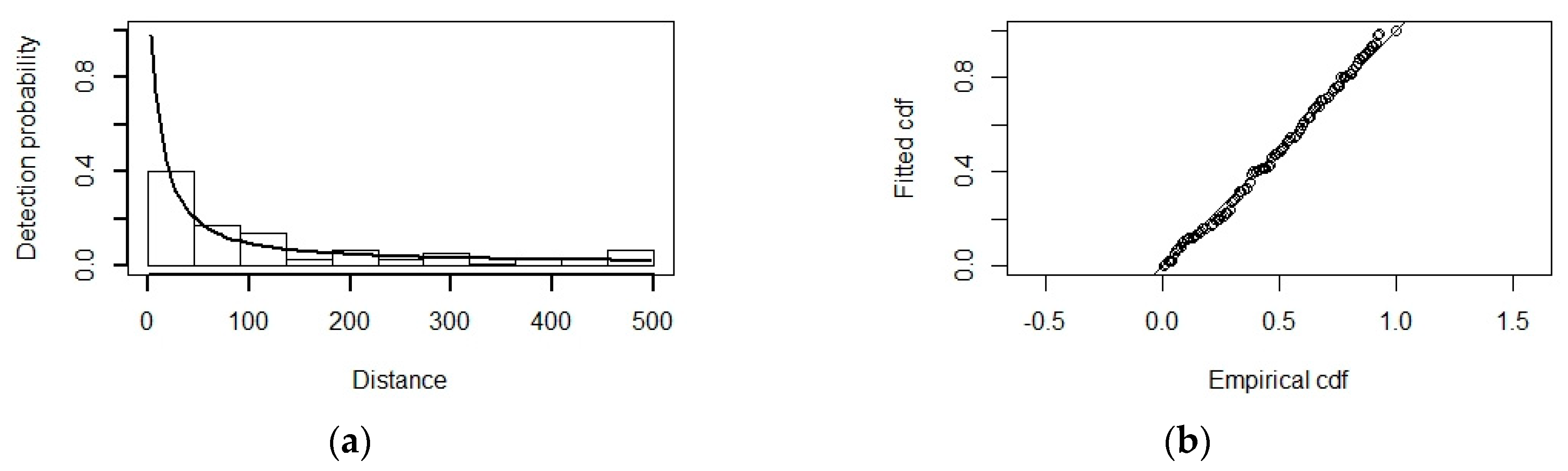
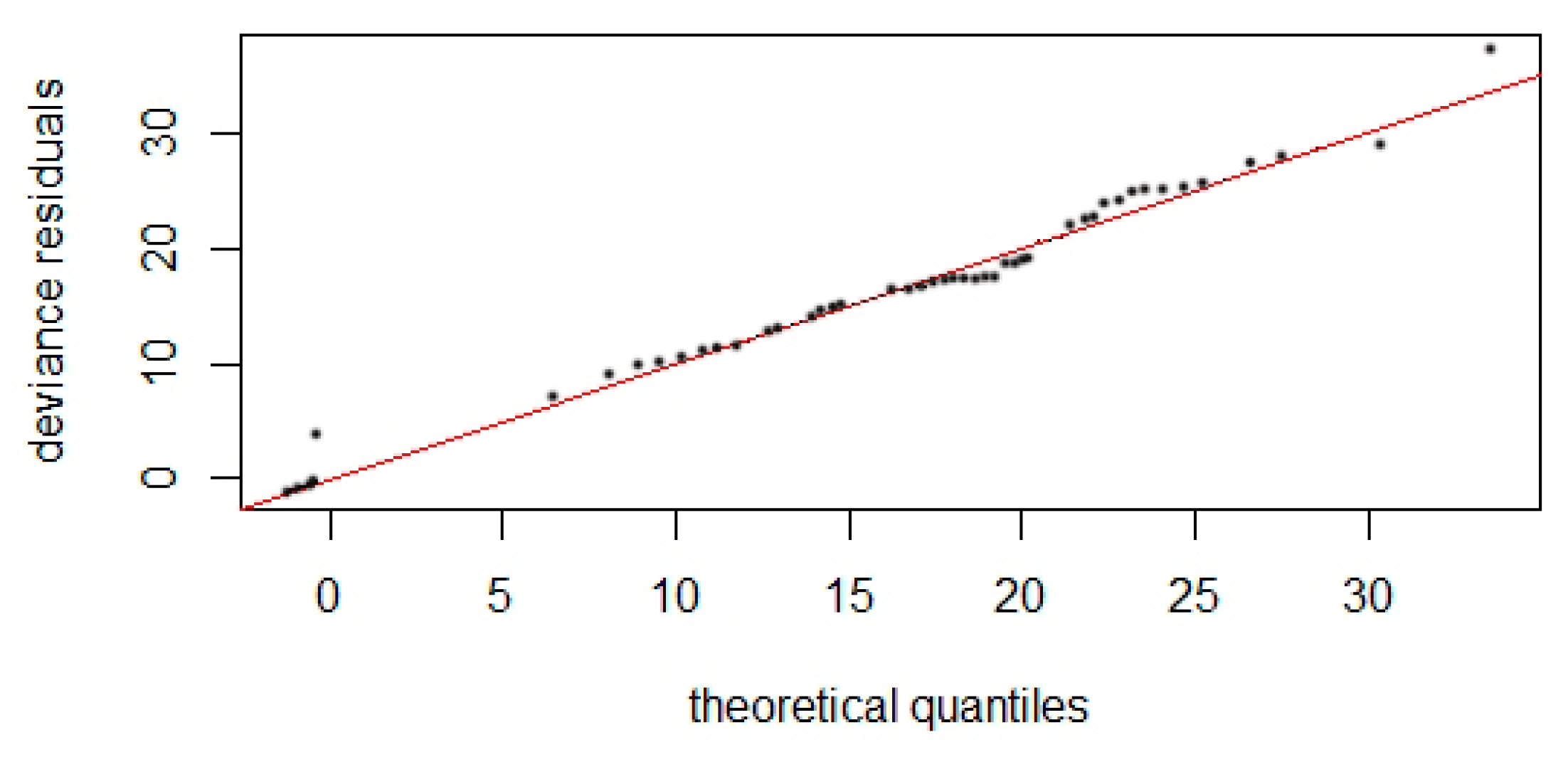
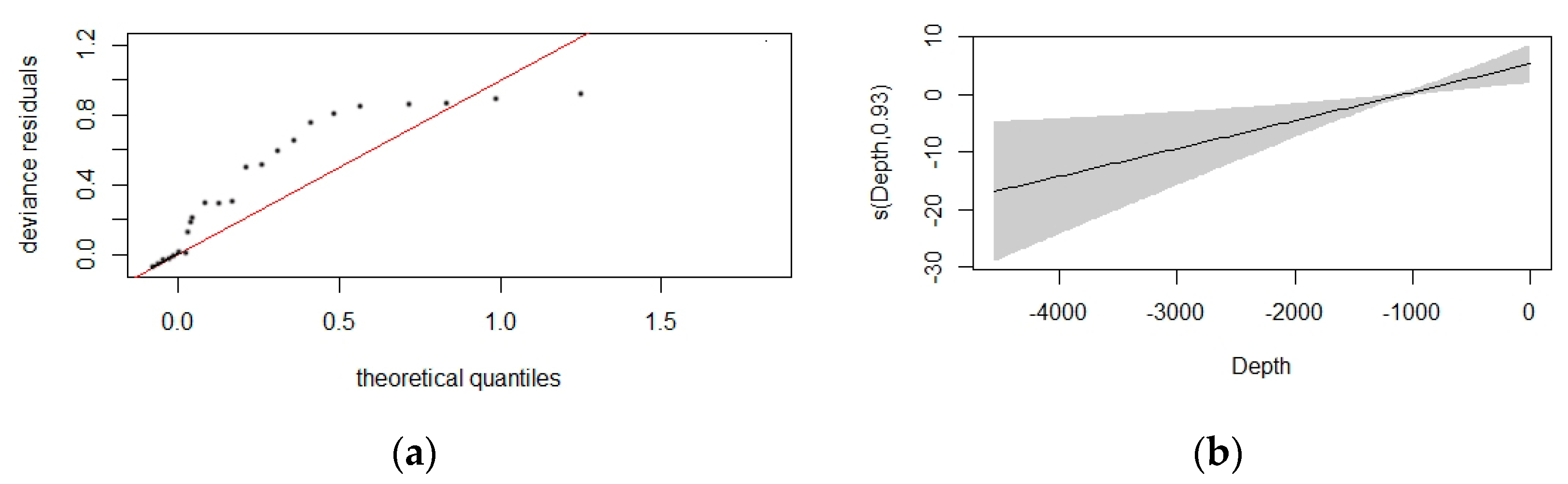
3.2. Detection Function
3.3. Density Surface Models and Predictions
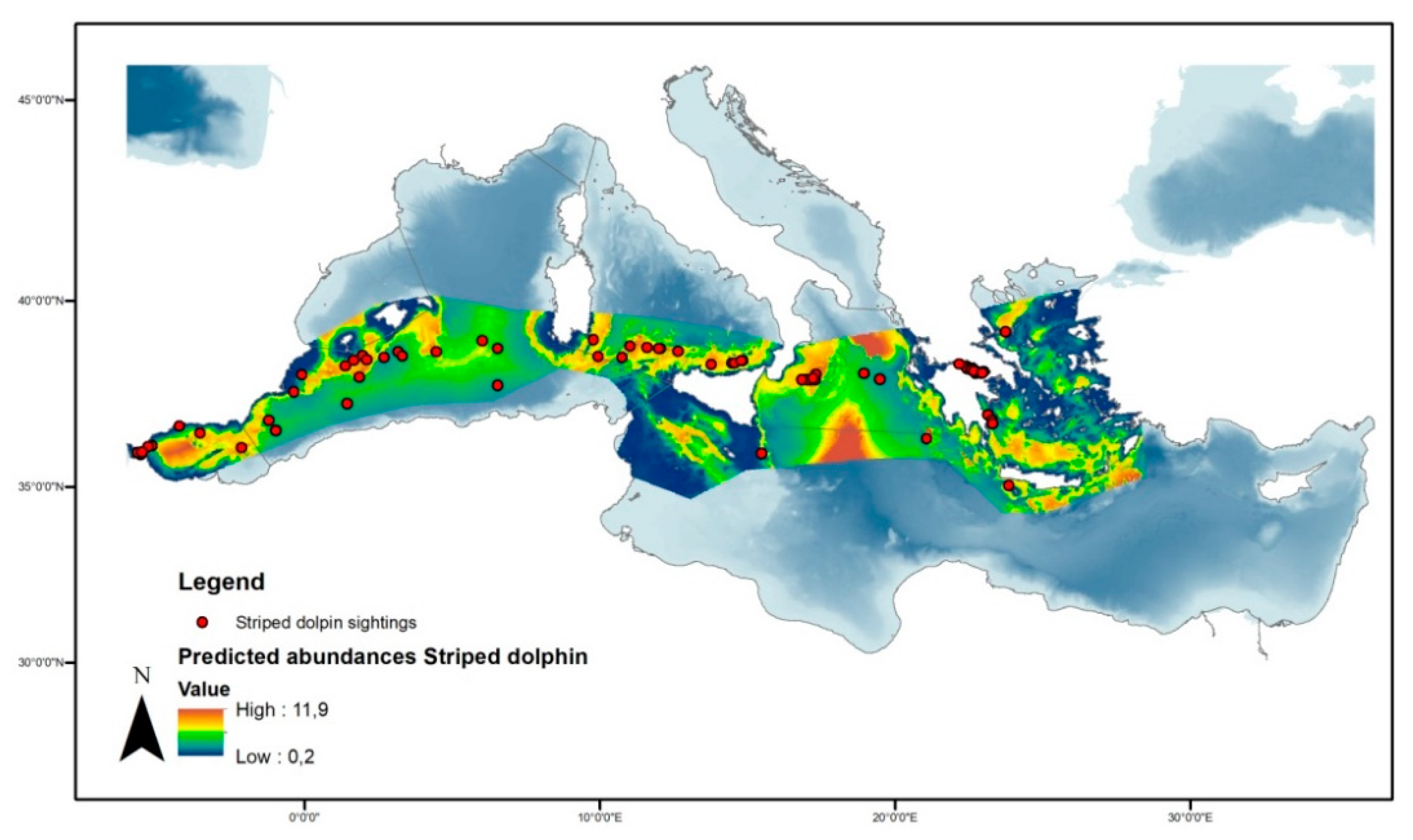
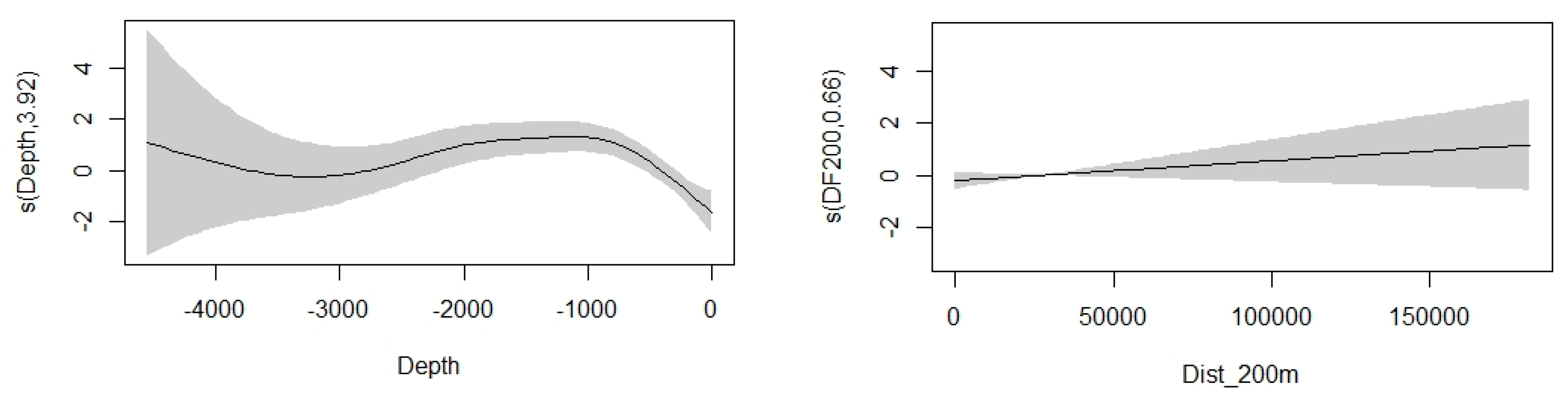
3.3.1. Striped Dolphin
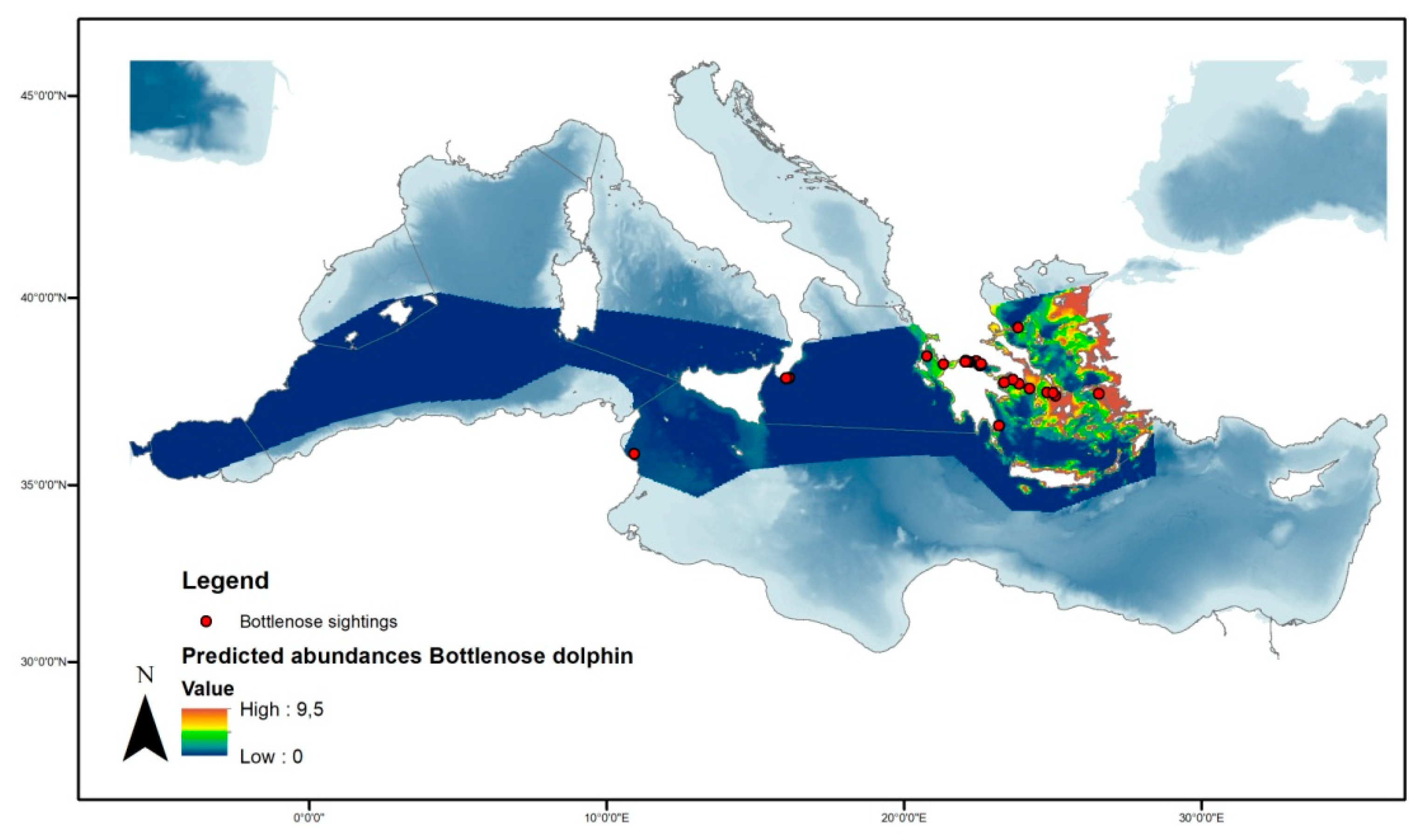
3.3.2. Bottlenose Dolphin
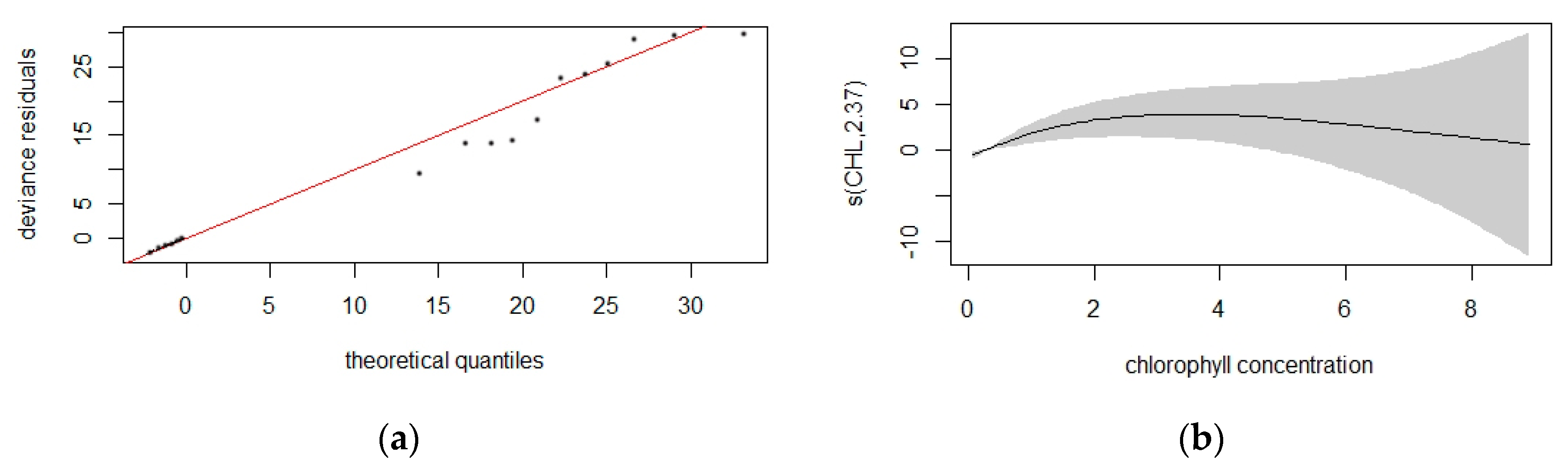
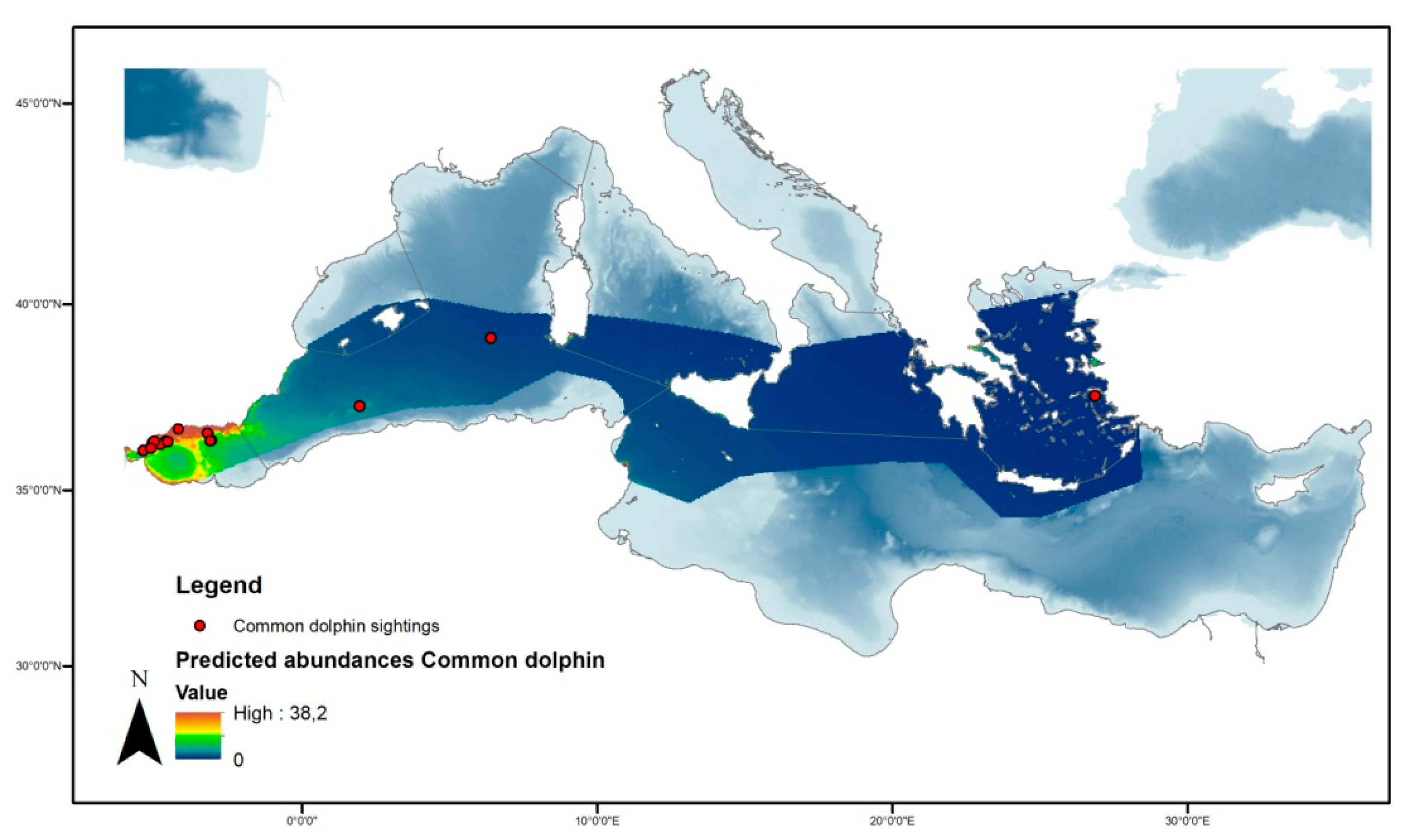
3.3.3. Common Dolphin
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Natoli, A.; Birkun, A.; Aguilar, A.; Lopez, A.; Hoelzel, A.R. Habitat structure and the dispersal of male and female bottlenose dolphins (Tursiops truncatus). Proc. R. Soc. B Biol. Sci. 2005, 272, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- Gaspari, S.; Azzelino, A.; Airoldi, S.; Hoelzel, A.R. Social kin associations and genetic structuring of striped dolphin populations (Stenella coeruleoalba) in the Mediterranean Sea. Mol. Ecol. 2007, 16, 2922–2933. [Google Scholar] [CrossRef] [PubMed]
- Gaspari, S.; Holcer, D.; Mackelworth, P.; Fortuna, C.; Frantzis, A.; Genov, T.; Vighi, M.; Natali, C.; Rako, N.; Banchi, E.; et al. Population genetic structure of common bottlenose dolphins (Tursiops truncatus) in the Adriatic Sea and contiguous regions: Implications for international conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 25, 212–222. [Google Scholar] [CrossRef]
- Gkafas, G.; Exadactylos, A.; Rogan, E.; Raga, J.; Reid, R.; Hoelzel, R. Biogeography and temporal progression during the evolution of striped dolphin population structure in European waters. J. Biogeogr. 2017, 44, 2681–2691. [Google Scholar] [CrossRef]
- Notarbartolo di Sciara, G.; Venturino, M.C.; Zanardelli, M.; Bearzi, G.; Borsani, J.F.; Cavalloni, B. Cetaceans in the central Mediterranean Sea: Distribution and sighting frequencies. Boll. Zool. 1993, 60, 131–138. [Google Scholar] [CrossRef]
- Bearzi, G.; Reeves, R.R.; Notarbartolo di Sciara, G.; Politi, E.; Cañadas, A.; Frantzis, A.; Mussi, B. Ecology, status and conservation of short-beaked common dolphins Delphinus delphis in the Mediterranean Sea. Mammal Rev. 2003, 33, 224–252. [Google Scholar] [CrossRef] [Green Version]
- Bearzi, G.; Holcer, D.; Notarbartolo di Sciara, G. The role of historical dolphin takes and habitat degradation in shaping the present status of northern Adriatic cetaceans. Aquat. Conserv. Mar. Freshw. Ecosyst. 2004, 14, 363–379. [Google Scholar] [CrossRef]
- Evans, P.G.H.; Hammond, P.S. Monitoring cetaceans in European waters. Mammal Rev. 2004, 34, 131–156. [Google Scholar] [CrossRef]
- Bearzi, G.; Politi, E.; Agazzi, S.; Bruno, S.; Costa, M.; Bonizzoni, S. Occurrence and present status of coastal dolphins (Delphinus delphis and Tursiops truncatus) in the eastern Ionian Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 243–257. [Google Scholar] [CrossRef]
- Gannier, A. Summer distribution and relative abundance of delphinids in the Mediterranean Sea. Rev. Ecol. 2005, 60, 223–238. [Google Scholar]
- Carlucci, R.; Fanizza, C.; Cipriano, G.; Paoli, C.; Russo, T. Modeling the spatial distribution of the striped dolphin (Stenella coeruleoalba) and common bottlenose dolphin (Tursiops truncatus) in the Gulf of Taranto (northern Ionian Sea, central-eastern Mediterranean Sea). Ecol. Indic. 2016, 69, 707–721. [Google Scholar] [CrossRef]
- Giménez, J.; Cañadas, A.; Ramírez, F.; Afán, I.; García-Tiscar, S.; Fernández-Maldonado, C.; Castillo, J.J.; de Stephanis, R. Intra- and interspecific niche partitioning in striped and common dolphins inhabiting the southwestern Mediterranean Sea. Mar. Ecol. Prog. Ser. 2017, 567, 199–210. [Google Scholar] [CrossRef]
- Bonizzoni, S.; Furey, N.B.; Santostasi, N.L.; Eddy, L.; Valavanis, V.D.; Bearzi, G. Modelling dolphin distribution within an Important Marine Mammal Area in Greece to support spatial management planning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1665–1680. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IUCN. Final Report of the Workshop: First IMMA Regional Workshop for the Mediterranean, Chania, Greece, 24–28 October 2017; Marine Mammal Protected Area Task Force of the International Union for Conservation of Nature: Gland. Available online: http://www.marinemammalhabitat.org/download/report-regional-workshop-medi-terranean-important-marine-mammal-areas/ (accessed on 25 September 2019).
- Millot, C.; Taupier-Letage, I. Circulation in the Mediterranean Sea. In The Hand-Book of Environmental Chemistry. The Natural Environment and the Biological Cycles; Springer: Berlin, Germany, 2005; pp. 29–66. Volume 5, Part K. [Google Scholar] [CrossRef]
- Curry, B.E. Advances in Marine Biology 70; Academic Press: London, UK, 2015; p. 318. [Google Scholar]
- Notarbartolo di Sciara, G.; Birkun, A., Jr. Conserving Whales, Dolphins and Porpoises in the Mediterranean and Black Seas: An ACCOBAMS Status Report, 2010; ACCOBAMS: Monaco City, Monaco, 2010; p. 212. [Google Scholar]
- Bearzi, G.; Fortuna, C.; Reeves, R. Tursiops truncatus (Mediterranean subpopulation). In The IUCN Red List of Threatened Species 2012; e.T16369383A16369386; IUCN: Gland, Switzerland, 2012. [Google Scholar] [CrossRef]
- Cañadas, A.; Sagarminaga, R.; Garcia-Tiscar, S.A. Cetacean distribution related with depth and slope in the Mediterranean waters off southern Spain. Deep-Sea Research Part I. Oceanogr. Res. Pap. 2002, 49, 2053–2073. [Google Scholar] [CrossRef]
- Hammond, P.S.; Bearzi, G.; Bjørge, A.; Forney, K.; Karczmarski, L.; Kasuya, T.; Wilson, B. Stenella coeruleoalba. In IUCN Red List of Threatened Species 2008; Version 2010.4; IUCN: Gland, Switzerland, 2010. [Google Scholar]
- Azzellino, A.; Fossi, M.C.; Gaspari, S.; Lanfredi, C.; Lauriano, G.; Marsili, L.; Panigada, S.; Podestà, M. An index based on the biodiversity of cetacean species to assess the environmental status of marine ecosystems. Mar. Environ. Res. 2014, 100, 94–111. [Google Scholar] [CrossRef] [PubMed]
- ACCOBAMS. Resolution 6.13. Comprehensive cetacean population estimates and distribution in the ACCOBAMS area (monitoring of cetacean distribution, abundance and accobams survey initiative). 2016. Available online: http://www.accobams.org/ (accessed on 20 August 2019).
- IUCN. Initial Guidance on the Use of Selection Criteria for the Identification of Important Marine Mammal Areas (IMMAs); Marine Mammal Protected Area Task Force of the International Union for Conservation of Nature: Gland, Switzerland, 2016; Available online: http://www.marinemammalhabitat.org/download/imma-guidance-document-october-2016/ (accessed on 1 September 2019).
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Advanced Distance Sampling: Estimating Abundance of Biological Population; Oxford University Press: New York, NY, USA, 2004. [Google Scholar]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Hedley, S.L.; Buckland, S.T. Spatial models for line transect sampling. J. Agric. Biol. Environ. Stat. 2004, 9, 181–199. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Redfern, J.V.; Ferguson, M.C.; Becker, A.E.; Hyrenbach, K.D.; Good, C.; Barlow, J.; Kaschner, K.; Baumgartner, M.F.; Forne, K.A.; Balance, L.T.; et al. Techniques for cetacean-habitat modeling. Mar. Ecol. Prog. Ser. 2006, 310, 271–295. [Google Scholar] [CrossRef]
- Ready, J.; Kaschner, K.; South, A.B.; Eastwood, P.D.; Rees, T.; Rius, J.; Agbayani, E.; Kullander, S.; Froese, R. Predicting the distributions of marine organisms at the global scale. Ecol. Model. 2010, 221, 467–478. [Google Scholar] [CrossRef]
- Dambach, J.; Rödder, D. Applications and future challenges in marine species distribution modeling. Aquat. Conserv. Mar. Freshw. Ecosyst. 2011, 21, 92–100. [Google Scholar] [CrossRef]
- Miller, D.L.; Burt, M.L.; Rexstad, A.E.; Thomas, L. Spatial models for distance sampling data: Recent developments and future directions. Methods Ecol. Evol. 2013, 4, 1001–1010. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.E.; Glegg, G.A.; Howell, K.L. Species distribution modelling to support marine conservation planning: The next steps. Mar. Policy 2014, 45, 330–332. [Google Scholar] [CrossRef]
- Sillero, N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Model. 2011, 8, 1343–1346. [Google Scholar] [CrossRef]
- Guisan, A.; Lehmann, A.; Ferrier, S.; Austin, M.; Overton, J.M.C.; Aspinall, R.; Hastie, T. Making better biogeographical predictions of species’ distributions. J. Appl. Ecol. 2006, 43, 386–392. [Google Scholar] [CrossRef]
- Thomas, L.; Buckland, S.T.; Rexstad, A.E.; Laake, J.L.; Strindberg, S.; Hedley, S.L.; Bishop, J.R.; Marques, T.A.; Burnham, K.P. Distance software: Design and analysis of distance sampling surveys for estimating population size. J. Appl. Ecol. 2010, 47, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Buckland, S.T.; Rexstad, A.E.; Marques, T.A.; Oedekoven, C.S. Distance Sampling: Methods and Applications; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Miller, D.L.; Rexstad, E.; Burt, L.; Bravington, M.V.; Hedley, S. Density Surface Modelling of Distance Sampling Data. 2019. Version 2.2.17. Available online: https://cran.r-project.org/web/packages/dsm/index.html (accessed on 1 October 2019).
- Bouchet, P.J.; Miller, D.L.; Roberts, J.J.; Mannocci, L.; Harris, C.M.; Thomas, L. From Here and Now to There and Then: Practical Recommendations for Extrapolating Cetacean Density Surface Models to Novel Conditions; Technical report 2019-01 v1.0; Centre for Research into Ecological & Environmental Modelling (CREEM), University of St Andrews: St Andrews, UK, 2019; p. 59. [Google Scholar]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L. Distance Sampling: Estimating Abundance of Biological Populations; Chapman & Hall: London, UK, 1993. [Google Scholar]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- NASA Goddard Space Flight Center, Ocean Ecology Laboratory, Ocean Biology Processing Group. Sea-viewing Wide Field-of-view Sensor (SeaWiFS) Ocean Color Data; NASA OB.DAAC: Greenbelt, MD, USA, 2014. [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- EMODnet Bathymetry Consortium. EMODnet Digital Bathymetry (DTM). 2018. Available online: https://doi.org/10.12770/18ff0d48-b203-4a65-94a9-5fd8b0ec35f6 (accessed on 14 September 2018). [CrossRef]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: London, UK, 2017. [Google Scholar]
- Borchers, D.L.; Buckland, S.T.; Goedhart, P.W.; Clarke, E.D.; Hedley, S.L. Horvitz-Thompson estimators for double-platform line transect surveys. Biometrics 1998, 54, 1221–1237. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: Berlin, Germany, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Miller, D.L.; Rexstad, E.; Thomas, L.; Marshall, L.; Laake, J.L. Distance Sampling in R. J. Stat. Softw. 2019, 89, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Candy, S. Modelling catch and effort data using generalised linear models, the Tweedie distribution, random vessel effects and random stratum-by-year effects. CCAMLR Sci. 2004, 11, 59–80. [Google Scholar]
- Roberts, J.J.; Best, B.D.; Mannocci, L.; Fujioka, E.; Halpin, P.N.; Palka, D.L.; Garrison, L.P.; Mullin, K.D.; Cole, T.V.N.; Khan, C.B.; et al. Habitat-based cetacean density models for the U.S. Atlantic and Gulf of Mexico. Sci. Rep. 2016, 6, 22615. [Google Scholar] [CrossRef] [PubMed]
- Redfern, J.V.; Moore, T.J.; Fiedler, P.C.; de Vos, A.; Brownell, R.L., Jr.; Forney, K.A.; Becker, A.E.; Ballance, L.T. Predicting cetacean distributions in data-poor marine ecosystems. Divers. Distrib. 2017, 23, 394–408. [Google Scholar] [CrossRef]
- Whitt, A.D.; Powell, J.A.; Richardson, A.G.; Bosyk, J.R. Abundance and distribution of Marine mammals in nearshore waters off New Jersey, USA. J. Cetacean Res. Manag. 2015, 15, 45–59. [Google Scholar]
- Cañadas, A.; Hammond, P.S. Abundance and habitat preferences of the short-beaked common dolphin Delphinus delphis in the southwestern Mediterranean: Implications for conservation. Endanger. Species Res. 2008, 4, 309–331. [Google Scholar] [CrossRef]
- Becker, E.; Foley, D.G.; Forney, K.; Barlow, J.; Redfern, J.; Gentemann, C.L. Forecasting cetacean abundance patterns to enhance management decisions. Endanger. Species Res. 2012, 16, 97–112. [Google Scholar] [CrossRef] [Green Version]
- Redfern, J.V.; McKenna, M.F.; Moore, T.; Calambokidis, J.; DeAngelis, M.L.; Becker, A.E.; Barlow, J.; Forney, K.A.; Fiedler, P.C.; Chivers, S.J. Assessing the risk of ships striking large whales in marine spatial planning. Conserv. Biol. 2013, 27, 292–302. [Google Scholar] [CrossRef]
- Karamitros, G.; Gkafas, G.A.; Giantsis, I.A.; Martsikalis, P.; Kavouras, M.; Exadactylos, A. Design-based and model-based estimations of distribution and abundance of dolphin populations in Gulf of Corinth, Hellas. In Proceedings of the 16th International Conference on Environmental Science and Technology (CEST 2019), Rhodes, Greece, 4–7 September 2019. [Google Scholar]
- Moura, A.E.; Natoli, A.; Rogan, E.; Hoelzel, A.R. Atypical panmixia in a European dolphin species (Delphinus delphis): Implications for the evolution of diversity across oceanic boundaries. J. Evol. Biol. 2013, 26, 63–75. [Google Scholar] [CrossRef]
- Giannoulaki, M.; Markoglou, E.; Valavanis, V.D.; Alexiadou, P.; Cucknell, A.C.; Frantzis, A. Linking small pelagic fish and cetacean distribution to model suitable habitat for coastal dolphin species, Delphinus delphis and Tursiops truncatus, in the Greek Seas (Eastern Mediterranean). Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 436–451. [Google Scholar] [CrossRef]
- Becker, E.; Forney, K.; Redfern, J.; Barlow, J.; Jacox, M.; Roberts, J.; Palacios, D. Predicting cetacean abundance and distribution in a changing climate. Divers. Distrib. 2019, 25, 626–643. [Google Scholar] [CrossRef]
- Bearzi, G. Delphinus delphis. In The IUCN Red List of Threatened Species 2012; e.T6336A16236707; IUCN: Gland, Switzerland, 2012. [Google Scholar]
- Bosc, E.; Bricaud, A.; Antoine, D. Seasonal and interannual variability in algal biomass and primary production in the Mediterranean Sea, as derived from 4 years of SeaWiFS observations. Glob. Biogeochem. Cycles 2004, 18. [Google Scholar] [CrossRef] [Green Version]
- Norabartolo di Sciara, G. Guida dei Mammiferi Marini del Mediterraneo; Muzzio editore: Padova, Italy, 1994. [Google Scholar]
- MacLeod, C.D.; Weir, C.R.; Begoña Santos, M.; Dunn, T.E. Temperature-based summer habitat partitioning between white-beaked and common dolphins around the United Kingdom and Republic of Ireland. J. Mar. Biol. Assoc. UK 2008, 88, 1193–1198. [Google Scholar] [CrossRef] [Green Version]
- Cañadas, A.; Sagarminaga, R.; De Stephanis, R.; Urquiola, E.; Hammond, P.S. Habitat preference modelling as a conservation tool: Proposals for marine protected areas for cetaceans in southern Spanish waters. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 495–521. [Google Scholar] [CrossRef]
- Azzellino, A.; Panigada, S.; Lanfredi, C.; Zanardelli, M.; Airoldi, S.; Notarbartolo di Sciara, G. Predictive habitat models for managing marine areas: Spatial and temporal distribution of marine mammals within the Pelagos Sanctuary (Northwestern Mediterranean sea). Ocean Coast. Manag. 2012, 67, 63–74. [Google Scholar] [CrossRef]
- Brotons, J.M.; Grau, A.M.; Rendell, L. Estimating the impact of interactions between bottlenose dolphins and artisanal fisheries around the Balearic Islands. Mar. Mammal Sci. 2008, 24, 112–127. [Google Scholar] [CrossRef]
- Hoelzel, A.R. Evolution of population structure in marine mammals. In Population Genetics for Animal Conservation; Bertorelle, G., Bruford, M.W., Hauffe, H.C., Rizzoli, A., Vernesi, C., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 294–319. [Google Scholar]
- Natoli, A.; Canadas, A.; Vaquero, C.; Politi, E.; Fernandez-Navarro, P.; Hoelzel, A.R. Conservation genetics of the short-beaked common dolphin (Delphinus delphis) in the Mediterranean Sea and in the eastern North Atlantic Ocean. Conserv. Genet. 2008, 9, 1479–1487. [Google Scholar] [CrossRef]
- Moura, A.E.; Nielsen, S.C.A.; Vilstrup, J.T.; Moreno-Mayar, J.V.; Gilbert, M.T.P.; Gray, H.W.I.; Natoli, A.; Möller, L.; Hoelzel, A.R. Recent diversification of a marine genus (Tursiops spp.) tracks habitat preference and environmental change. Syst. Biol. 2013, 62, 865–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Name Used | Source | Spatial Resolution | Unit |
|---|---|---|---|---|
| Depth | Depth | GEBCO | 1/16 arc | m |
| Slope % | Slope | GIS calculations | 1/16 arc | Degree |
| Distance from coast | DFC | GIS calculations | - | m |
| Distance from 200 m isobaths | DF200 | GIS calculations | - | m |
| Sea surface temperature | SST | MODIS (NASA (b), 2013) | 4 km | °C |
| Chlorophyll-a | CHL | MODIS (NASA (b), 2013) | 4 km | mg/m3 |
| Longitude/latitude | x/y | Europe Lambert Conformal Conic | - | m |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karamitros, G.; Gkafas, G.A.; Giantsis, I.A.; Martsikalis, P.; Kavouras, M.; Exadactylos, A. Model-Based Distribution and Abundance of Three Delphinidae in the Mediterranean. Animals 2020, 10, 260. https://doi.org/10.3390/ani10020260
Karamitros G, Gkafas GA, Giantsis IA, Martsikalis P, Kavouras M, Exadactylos A. Model-Based Distribution and Abundance of Three Delphinidae in the Mediterranean. Animals. 2020; 10(2):260. https://doi.org/10.3390/ani10020260
Chicago/Turabian StyleKaramitros, Grigorios, Georgios A. Gkafas, Ioannis A. Giantsis, Petros Martsikalis, Menelaos Kavouras, and Athanasios Exadactylos. 2020. "Model-Based Distribution and Abundance of Three Delphinidae in the Mediterranean" Animals 10, no. 2: 260. https://doi.org/10.3390/ani10020260
APA StyleKaramitros, G., Gkafas, G. A., Giantsis, I. A., Martsikalis, P., Kavouras, M., & Exadactylos, A. (2020). Model-Based Distribution and Abundance of Three Delphinidae in the Mediterranean. Animals, 10(2), 260. https://doi.org/10.3390/ani10020260