Metataxonomic and Histopathological Study of Rabbit Epizootic Enteropathy in Mexico


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
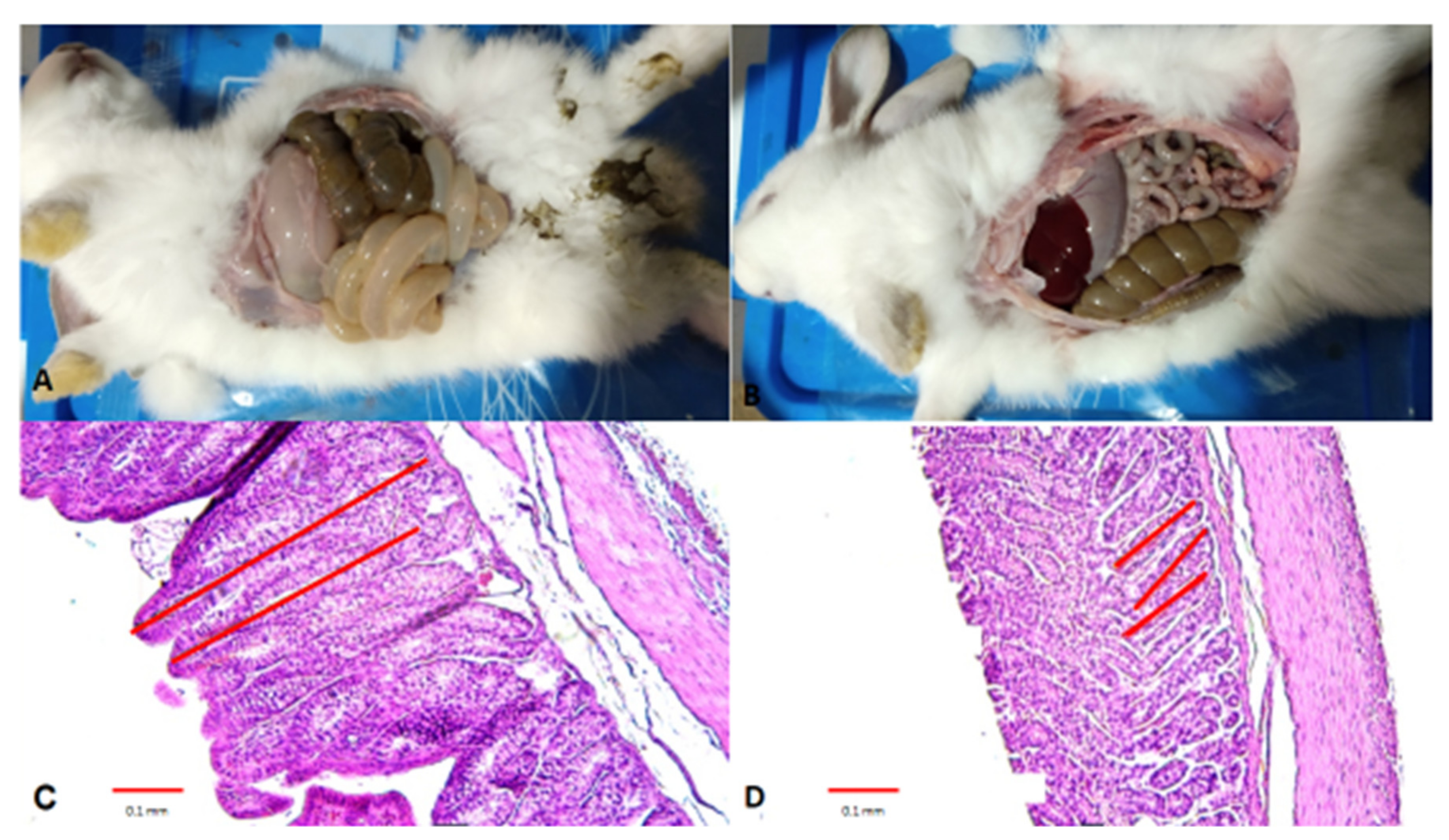
3.1. Histopathological Study
3.2. Metataxonomic Study
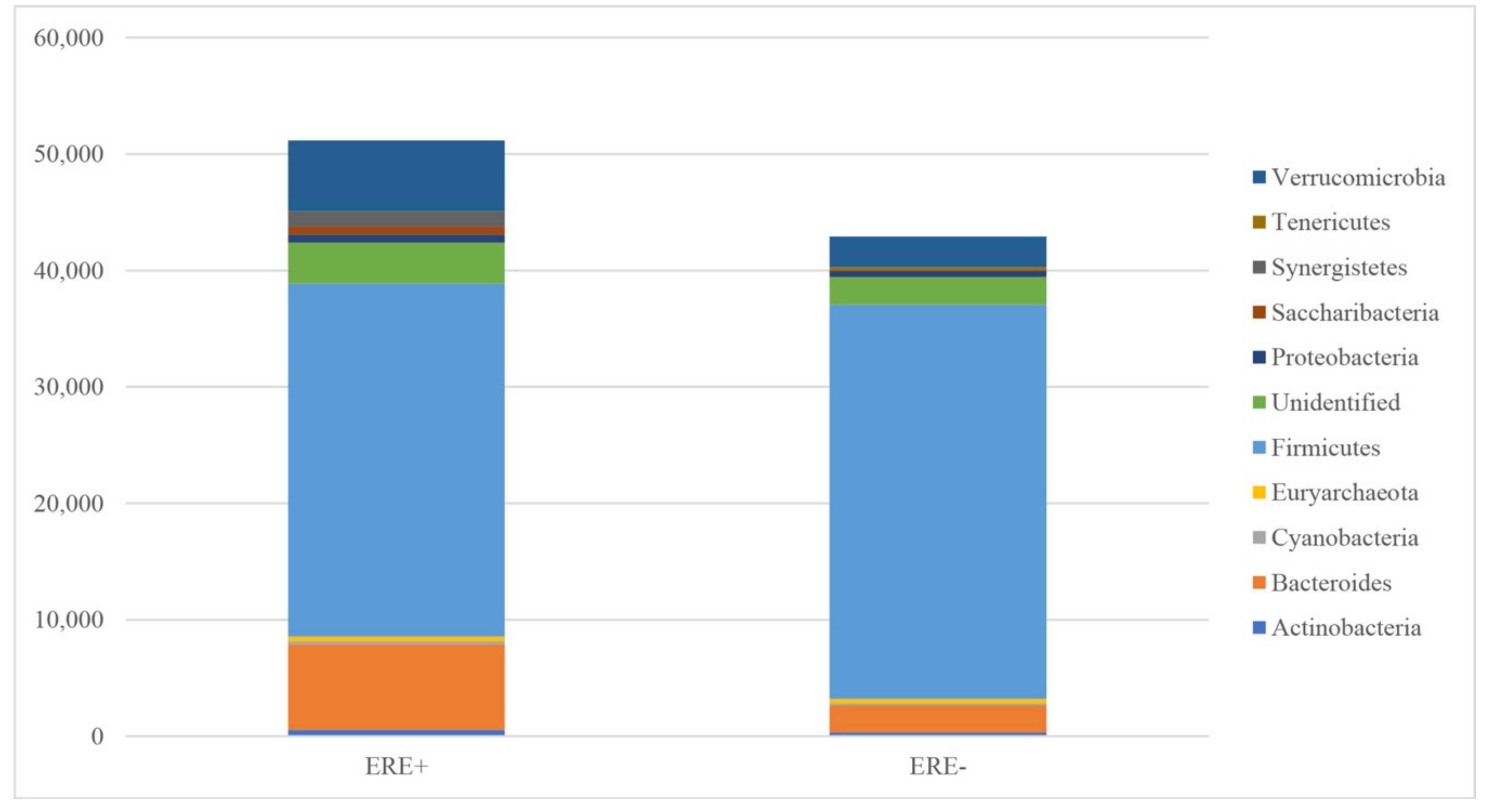
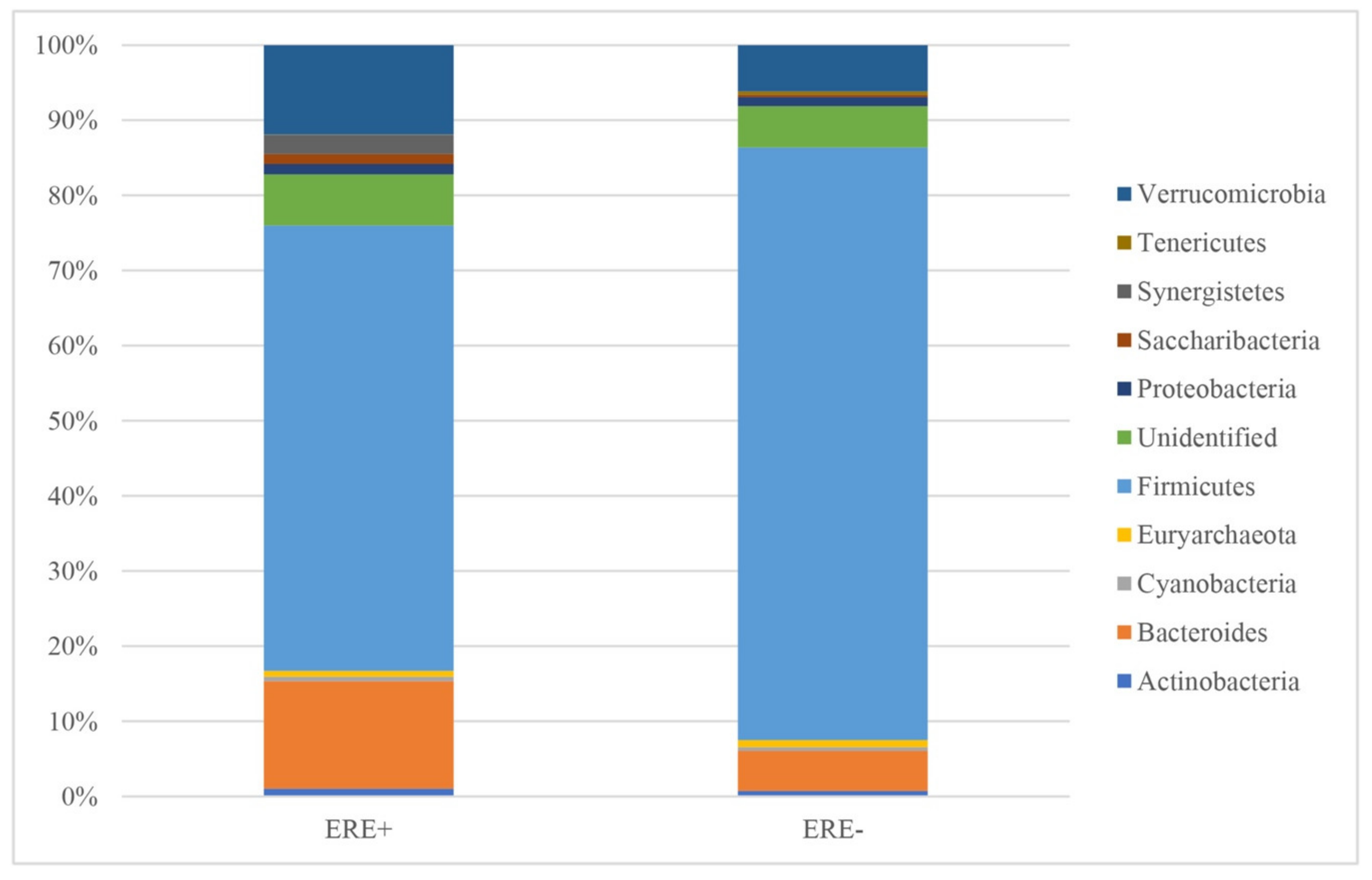
3.2.1. Bacterial Abundance
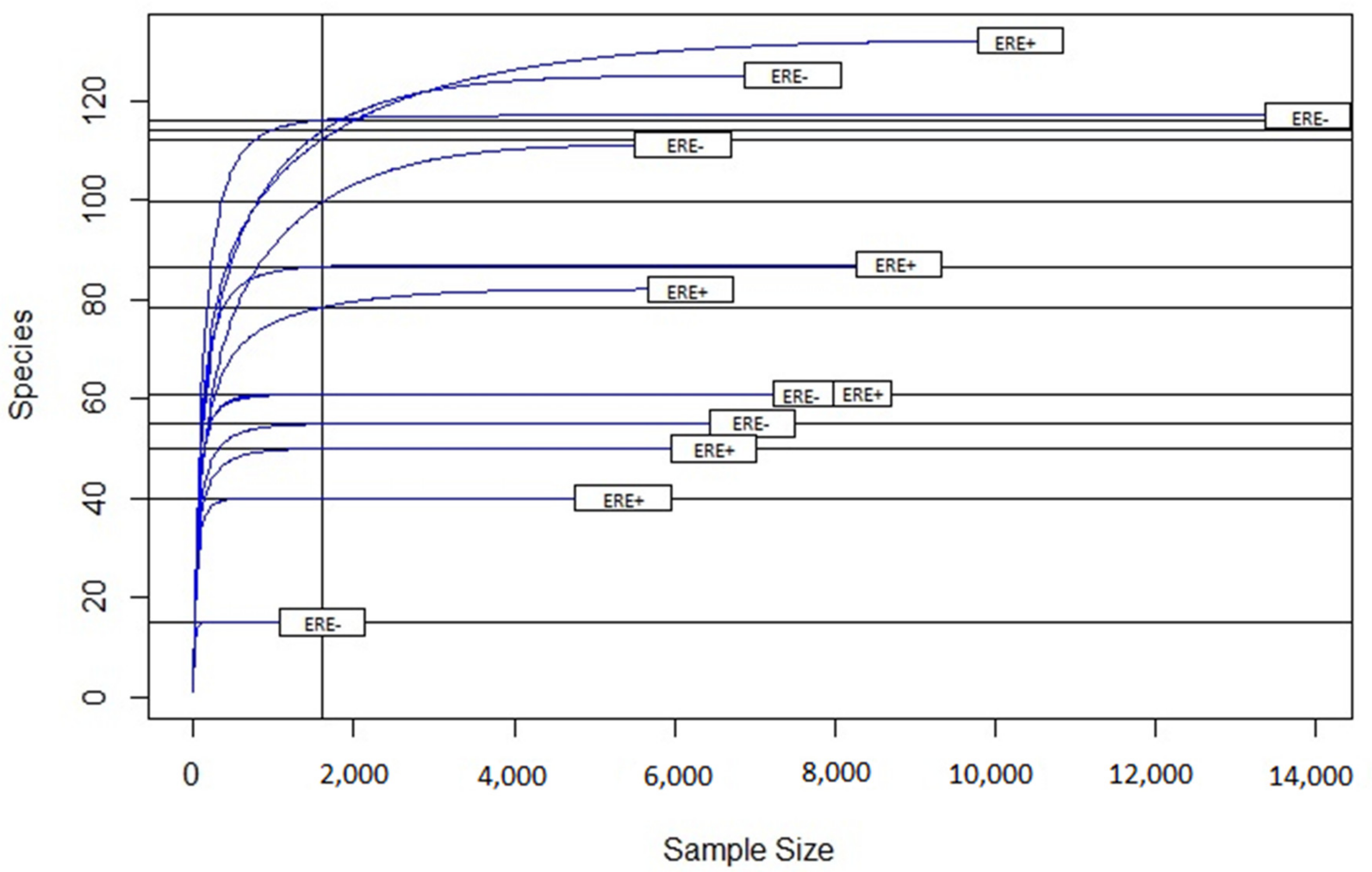
3.2.2. Bacterial Diversity
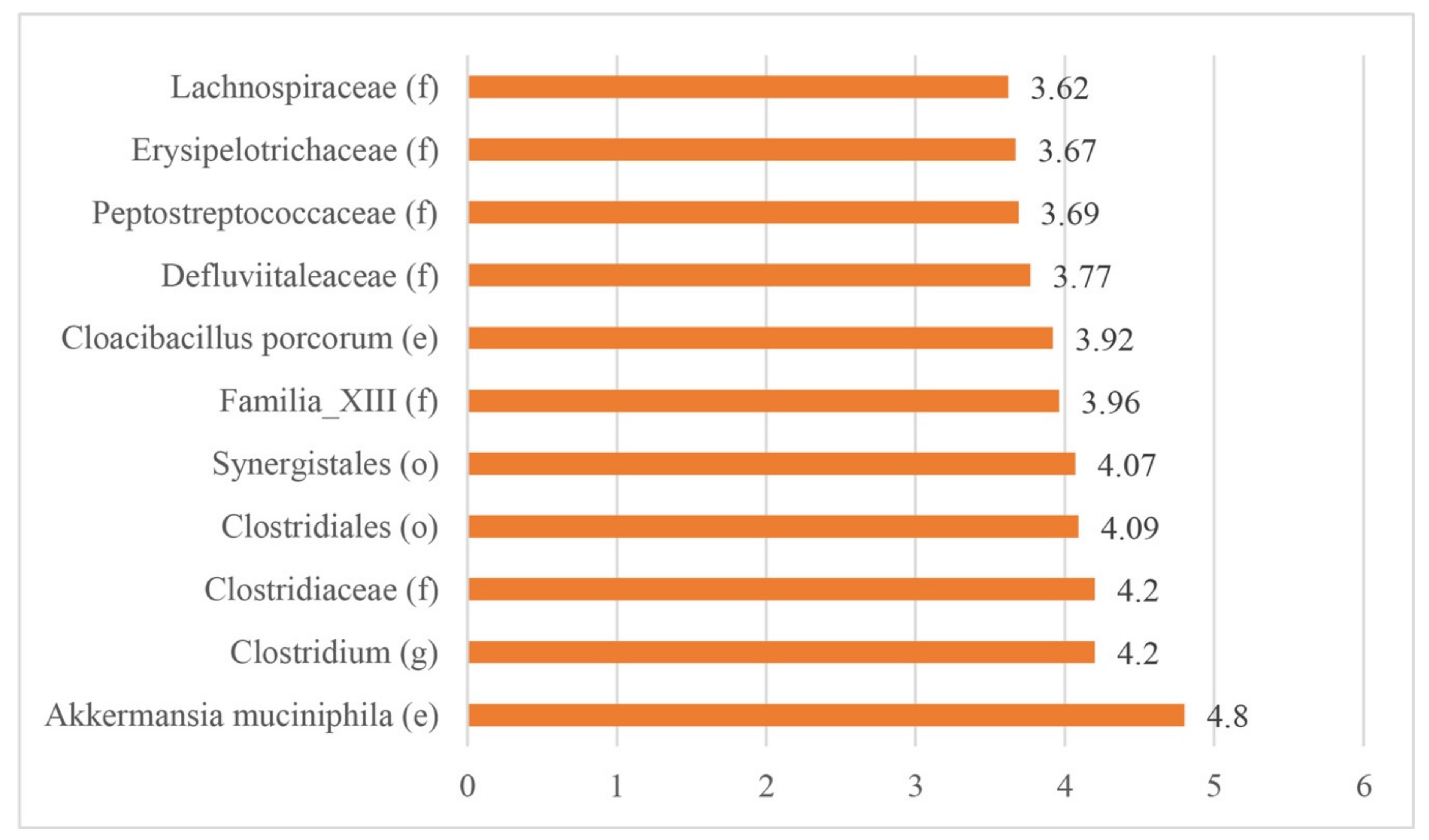
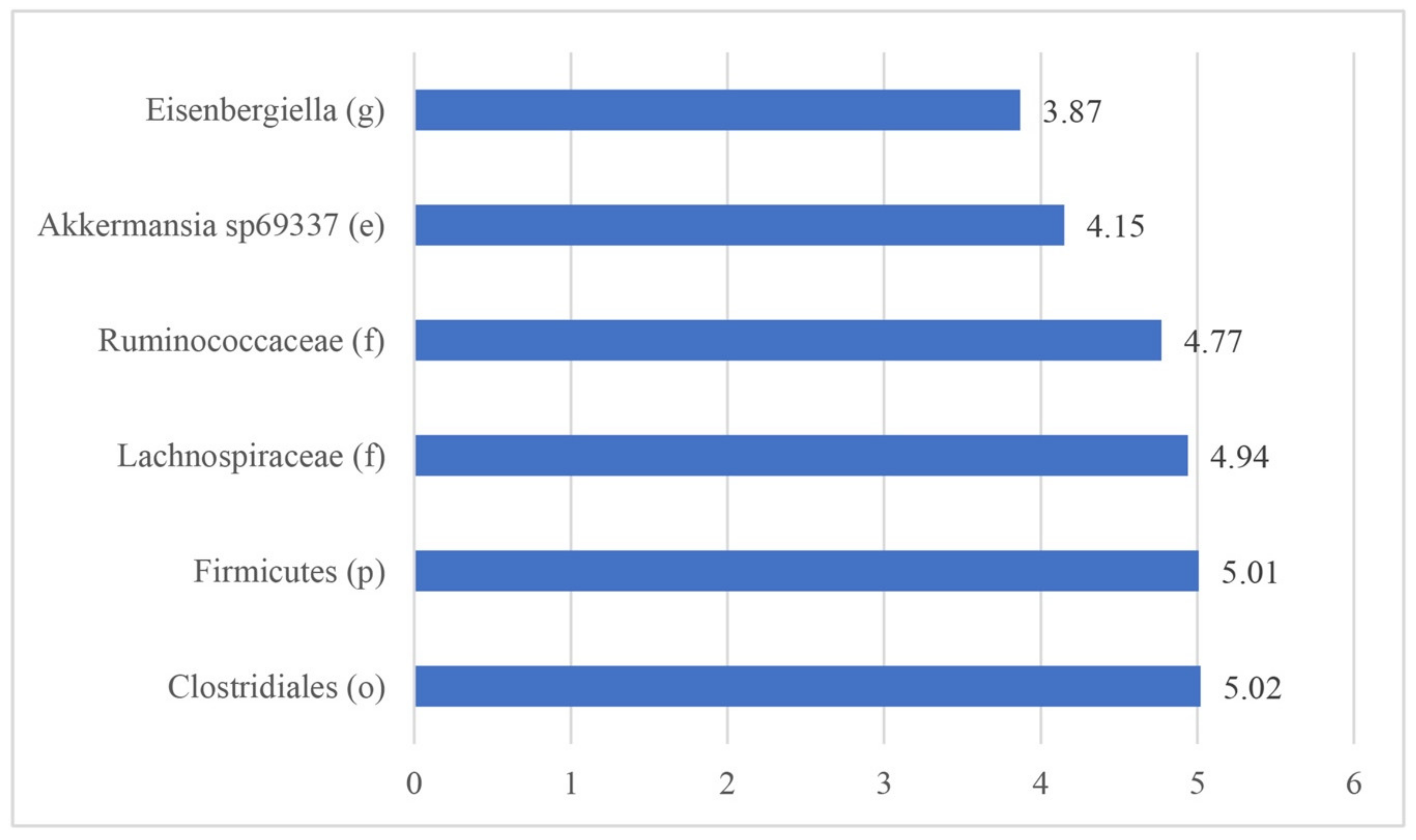
3.2.3. Biomarkers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Licois, D.; Vautherot, J.F.; Coudert, P.; Dambrine, G. Modele de Reproduction Experimentale de L’enterocolite Epizootique Chez Des Lapins Eops. World Rabbit Sci. 1998, 6, 349–353. [Google Scholar] [CrossRef] [Green Version]
- Bouquin, S.; Jobert, J.L.; Larour, G.; Balaine, L.; Eono, F.; Boucher, S.; Huneau, A.; Michel, V. Risk Factors for an Acute Expression of Epizootic Rabbit Enteropathy Syndrome in Rabbits after Weaning in French Kindling-to-Finish Farms. Livest. Sci. 2009, 125, 283–290. [Google Scholar] [CrossRef]
- Lebas, F.; Coudert, P.; de Rochambeau, H.; Thebault, R.G. El Conejo Cría y Patología. Collecion FAO: Producción y Sanidad Animal; FAO: Rome, Italy, 1996; Volume 19, ISSN 1014-6423. [Google Scholar]
- De Rozas Ruiz de Gauna, A.M.P.; Carabaño, R.; García, J.; Rosell, J.M.; Díaz Cano, J.V.; Barbé Garcia, J.; Pascual Amorós, J.J.; Badiola Saíz, J.I. Etiopatogenia de la enteropatía epizoótica del conejo. Boletín de Cunicultura 2005, 139, 167–174. [Google Scholar]
- Rodríguez Serrano, T.M. La ERE, enteropatía epizoótica del conejo. Boletín de Cunicultura 2004, 135, 39–46. [Google Scholar]
- Pérez, C.; Pérez, A.; Coscelli, A.; Prieto, A.; Díaz-Baños, P. Identificación de infecciones digestivas en conejos: Orientaciones para optimizar la prevención y el control. Boletín de cunicultura 2015, 176, 14–18. [Google Scholar]
- Pérez, J.E. Revisión y conclusiones de la enteropatía epizoótica del conejo. Cunicultura 2013, 38, 9–12. [Google Scholar]
- Abecia, L.; Fondevila, M.; Balcells, J.; McEwan, N.R. The effect of lactating rabbit does on the development of the caecal microbial community in the pups they nurture. J. Appl. Microbiol. 2007, 103, 557–564. [Google Scholar] [CrossRef]
- Licois, D.; Wyers, M.; Coudert, P. Epizootic Rabbit Enteropathy: Experimental transmission and clinical characterization. Vet. Res. 2005, 36, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Dewrée, R.; Meulemans, L.; Lassence, C.; Desmecht, D.; Ducatelle, R.; Mast, J.; Licois, D.; Vindevogel, H.; Marlier, D. Experimentally induced epizootic rabbit enteropathy: Clinical, histopathological, ultrastructural, bacteriological and haematological findings. World Rabbit Sci. 2007, 15, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Licois, D.; Coudert, P.; Cerén, N.; Vautherot, J.-F. Epizootic Enterocolitis of the rabbit: Review of current research. World Rabbit Sci. 2000, 8 (Suppl. 1), 187–194. [Google Scholar]
- Fernández, G. Enfermedades Infecciosas que cursan con procesos digestivos en conejos. Boletín de Cunicultura Lagomorpha 2006, 144, 23–40. [Google Scholar]
- Haligur, M.; Ozmen, O.; Demir, N. Pathological and Ultrastructural Studies on Mucoid Enteropathy in New Zealand Rabbits. J. Exot. Pet Med. 2009, 18, 224–228. [Google Scholar] [CrossRef]
- Allen, A.L.; Bryant, U.K. What’s Up Doc? Can. Vet. J. 2009, 50, 1297–1299. [Google Scholar] [PubMed]
- Bäuerl, C.; Collado, M.C.; Zúñiga, M.; Blas, E.; Pérez Martínez, G. Changes in Cecal Microbiota and Mucosal Gene Expression Revealed New Aspects of Epizootic Rabbit Enteropathy. PLoS ONE 2014, 9, e105707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huybens, N.; Horueix, J.; Licois, D.; Mainil, J.; Marlier, D. Pyrosequencing of epizootic rabbit enteropathy inocula and rabbit caecal samples. Vet. J. 2013, 196, 109–110. [Google Scholar] [CrossRef]
- Abecia, L.; Rodríguez-Romero, N.; Martínez-Fernández, G.; Martínez-Vallespín, B.; Fondevila, M. Pyrosequencing study of cecal bacterial community of rabbit does and kits from a farm affected by epizootic rabbit enteropathy. World Rabbit Sci. 2017, 25, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.X.; Zou, H.W.; Liu, S.Q.; Wang, L.Z.; Xue, B.; Wu, D.; Tian, G.; Cai, J.; Yan, T.H.; Wang, Z.S.; et al. The underlying microbial mechanism of epizootic rabbit enteropathy triggered by a low fiber diet. Sci. Rep. 2018, 8, 12489. [Google Scholar] [CrossRef]
- Park, S.-J.; Kim, J.; Lee, J.-S.; Rhee, S.-K.; Kim, H. Characterization of the fecal microbiome in different swine groups by high-throughput sequencing. Anaerobe 2014, 28, 157–162. [Google Scholar] [CrossRef]
- Borsanelli, A.C.; Lappin, D.F.; Viora, L.; Bennett, D.; Dutra, I.S.; Brandt, B.W.; Riggio, M.P. Microbiomes associated with bovine periodontitis and oral health. Vet. Microbiol. 2018, 218, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution simple inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Norma Oficial Mexicana NOM-062-ZOO-1999. Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio. Sección 9. Available online: http://www.fmvz.unam.mx/fmvz/principal/archivos/062ZOO.PDF (accessed on 25 January 2019).
- Gómez Soto, J.G.; Reis de Souza, T.C.; Mariscal Landin, G.; Aguilera Barreyro, A.; Bernal Santos, M.G.; Escobar Garcia, K. Gastrointestinal morphophysiology and presence of kafirins in ileal digesta in growing pigs fed sorghum-based diets. J. Appl. Anim. Res. 2018, 46, 618–625. [Google Scholar] [CrossRef] [Green Version]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach, 2nd ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Community Ecology Package. 2019. Available online: https://github.com/vegandevs/vegan (accessed on 23 June 2019).
- The R Foundation. Introduction to R. Disponible, 2019. Available online: https://www.r-project.org/about.html (accessed on 20 May 2020).
- Rodríguez-De Lara, R.; Cedillo-Peláez, C.; Constantino, F.; Fallas-López, M.; Cobos-Peralta, M.A.; Gutiérrez-Olvera, C.; Juárez-Acevedo, M.; Miranda-Romero, L.A. Studies on the evolution, pathology, and immunity of commercial fattering rabbits affected with epizootic outbreaks of diarrhoeas in México: A case report. Research in Veterinary Sci. 2008, 84, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Aslam, S.; Durrani, A.Z.; Rizwan, M.; Kashif, M. Comparative Efficacy of metronidazole in combination colistinsulphate and probiotics against Clostridium perfringens and their effects on histopathological features of visceral organs in broiler. Veterinaria 2016, 4, 11–17. [Google Scholar]
- Butler, D.; Torres-Torres, S.; Pahud, B.; Myers, A.; Bass, J.A.; St Peter, S.D.; McCulloh, R. Clostridium difficile ileitis in pediatric inflammatory bowel disease. J. Pediatric Gastroenterol. Nutr. 2017, 64, e156–e158. [Google Scholar] [CrossRef]
- Djukivic, A.; García-Garcera, M.; Martínez-Paredes, E.; Issac, S.; Artacho, A.; Martínez, J.; Ubeda, C. Gut colonization by a novel Clostridium species is associated with the onset of epizootic rabbit enteropathiy. Vet. Res. 2018, 49, 213. [Google Scholar] [CrossRef] [Green Version]
- Crowley, E.J.; Kink, J.M.; Wilkinson, T.; Worgan, H.J.; Huson, K.M.; Rose, M.T.; McEwan, N.R. Comparison of the microbial population in rabbits and guinea pigs by next generation sequencing. PLoS ONE 2017, 12, e0165779. [Google Scholar] [CrossRef] [Green Version]
- Read, T.; Fortun-Lamothe, L.; Pascal, G.; Le Boulch, M.; Cauquil, L.; Gabinaud, B.; Banneller, C.; Balmisse, E.; Destombes, N.; Bouchez, O.; et al. Diversity and co-occurrence pattern analysis of cecal microbiota establishment at the onset of solid feeding in young rabbits. Front. Microbiol. 2019, 10, 973. [Google Scholar] [CrossRef]
- Fouts, D.E.; Szpakowski, S.; Purushe, J.; Torralba, M.; Waterman, R.C.; MacNeil, M.D.; Alexander, L.J.; Nelson, K.E. Next Generation Sequencing to Define Prokaryotic and Fungal Diversity in the Bovine Rumen. PLoS ONE 2012, 7, e48289. [Google Scholar] [CrossRef]
- Gruninger, R.J.; Sensen, C.W.; McAllister, T.A.; Forster, R.J. Diversity of Rumen Bacteria in Canadian Cervids. PLoS ONE 2014, 9, e89682. [Google Scholar] [CrossRef]
- Zhou, X.; Li, Y.; Liu, S.; Yang, Q.; Su, X.; Zhou, L.; Tang, M.; Fu, R.; Li, J.; Huang, Q. Ultra-deep sequencing enables high-fidelity recovery of biodiversity for bulk arthropod samples without PCR amplification. Gigascience 2013, 2, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaura, E. Next-generation Sequencing Approaches to Understanding the Oral Microbiome. Adv. Dent. Res. 2016, 24, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Ebbert, M.T.W.; Wadsworth, M.E.; Staley, L.A.; Hoyt, K.L.; Pickett, B.; Miller, J.; Duce, J.; Kauwe, J.S.K.; Ridge, P.G. Evaluating the necessity of PCR duplicate removal from next-generation sequencing data and a comparison of approaches. BMC Bioinform. 2016, 17, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aird, D.; Ross, M.G.; Chen, W.-S.; Danielsson, M.; Fennell, T.; Russ, C.; Jaffe, D.B.; Nusbaum, C.; Gnirke, A. Analyszing and minimizing PCR amplification bias in Illumina sequencing libraries. Genome Biol. 2011, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kebschull, J.M.; Zador, A.M. Sources of PCR-induced distortions in high-throughput sequencing data sets. Nucleic Acids Res. 2015, 43, e143. [Google Scholar] [CrossRef] [Green Version]
- Tilak, M.K.; Botero-Castro, F.; Galtier, N.; Nabholz, B. Illumina library preparation for sequencing the GC-rich frection of heterogeneous genomic DNA. Genome Biol. Evol. 2018, 10, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Boisot, P.; Duperray, J.; Guyonvarch, A.; Richard, A.; Licois, D.; Coudert, P. Evaluation of the effectiveness of soluble bacitracin (Bacivet s®) in drinking water compared to bacitracin in the feed (Albac®), during an experimental reproduction of epizootic rabbit enteropathy syndrome. México. In Proceedings of the 8th World Rabbit Congress, Puebla, Mexico, 7–10 September 2004; pp. 457–462. [Google Scholar]
- Abecia, L.; Fondevila, M.; Balcells, J.; Edwards, J.E.; Newbold, C.J.; McEwan, N.R. Molecular profiling of bacterial species in the rabbit caecum. FEMS Microbiol. Lett. 2005, 244, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, S.; Dal Bosco, A.; Combes, S.; Moscati, L.; Crotti, S.; Cartoni, A.; Cotozzolo, E.; Castellini, C. Dehydrated alfalfa and fresh grass supply in young rabbits: Effect on performance and cecal microbiota biodiversity. Animals 2019, 9, 341. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Liu, S.; Wang, Z.; Wang, L.; Xue, B.; Zou, H.; Tian, G.; Cai, J.; Peng, Q. Effects of particle size of ground alfalfa on cecal bacteria and archaea populations of rabbits. PeerJ 2019, 7, e7910. [Google Scholar] [CrossRef]
- De Blas, J.C.; Astillero, J.R.; Chamorro, S.; Corujo, A.; García-Alonso, J.; García-Rebollar, P.; García-Ruiz, A.I.; Menoyo, D.; Nicodemus, N.; Romero, C.; et al. Efectos de la nutrición y el manejo sobre el desarrollo de patologías digestivas de gazapos en un entorno de enteropatía epizoótica. 2007. Salud intestinal en gazapos. FEDNA. pp. 109–124. Available online: http://fundacionfedna.org/sites/default/files/07CAP_VI.pdf (accessed on 26 March 2020).
- Chamorro, S.; De Blas, C.; Grant, G.; Badiola, I.; Menoyo, D.; Carabaño, R. Effect of dietary supplementation of glutamine-arginine on intestinal health in twenty-five-day-old weaned rabbits. J. Anim. Sci. 2010, 88, 170–180. [Google Scholar] [CrossRef]
- Macchione, I.G.; Lopetuso, L.R.; Ianiro, G.; Napoli, M.; Gibiino, G.; Rizzatti, G.; Petito, V.; Gasbarrini, A.; Scaldaferri, F. Akkermansia muciniphila: Key player in metabolic and gastrointestinal disorders. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8075–8083. [Google Scholar] [CrossRef] [PubMed]
- López-Siles, M.; Enrich-Capó, N.; Aldeguer, X.; Sabat-Mir, M.; Duncan, S.H.; García-Gil, L.J.; Martínez-Medina, M. Alterations in the Abundance and Co-occurrence of Akkermansia muciniphila and Faecalibacterium prausnitzii in the Colonic Mucosa of Inflammatory Bowel Disease Subjects. Front. Cell. Infect. Microbiol. 2018, 8, 281. [Google Scholar] [CrossRef] [PubMed]
- Earley, H.; Lennon, G.; Balfe, A.; Coffey, J.C.; Winter, D.C.; O´Connell, P.R. The abundance of Akkermansia muciniphila and its relationship with sulphated colonic mucins in health and ulcerative colitis. Sci. Rep. Nat. Re. 2019, 9, 15683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, M.; Belzer, C.; M de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia miciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2016, 23, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Bian, X.; Wu, W.; Yang, L.; Lv, L.; Wang, Q.; Li, Y.; Ye, J.; Fang, D.; Wu, J.; Jiang, X.; et al. Administration of Akkermansia muciniphila Ameliorates Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice. Front. Microbiol. 2019, 10, 2259. [Google Scholar] [CrossRef] [Green Version]
- Vo, N.; Tsai, T.C.; Maxwell, C.; Carbonero, F. Early exposure to agricultural soil accelerates the maturation of the early-life pig gut microbiota. Anaerobe 2017, 45, 31–39. [Google Scholar] [CrossRef]
- Looft, T.; Levine, U.Y.; Stanton, T.B. Cloacibacillus porcorum sp. nov., a mucin-degrading bacterium from the swine intestinal tract and emended description of the genus Cloacibacillus. Int. J. Syst. Evol. Microbiol. 2013, 63, 1960–1996. [Google Scholar] [CrossRef]
- Eshar, D.; Weese, J.S. Molecular analysis of the microbiota in hard feces from healthy rabbits (Oryctolagus cuniculus) medicated with long term oral meloxicam. BMC Vet. Res. 2014, 10, 62. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Depth of Crypts (µm) | |||
|---|---|---|---|---|
| ERE+ | ERE− | SEM | p Value | |
| Ileo-ceco-colic valve | 199 | 187 | 16.21 | 0.73 |
| Cecum | 136 | 121 | 5.55 | 0.22 |
| Colon | 173 | 308 | 10.51 | 0.0004 |
| Phylum | ERE+ | % | ERE− | % | p Value |
|---|---|---|---|---|---|
| Actinobacteria | 538 ± 108.5 | 1.05 | 312 ± 217.9 | 0.73 | 0.138 |
| Bacteroides | 7327 ± 2934.8 | 14.32 | 2300 ± 1020.4 | 5.36 | 0.137 |
| Cyanobacteria | 286 ± 88.3 | 0.56 | 214 ± 58.1 | 0.50 | 0.508 |
| Euryarchaeota | 400 ± 97.3 | 0.78 | 387 ± 194.2 | 0.90 | 0.951 |
| Firmicutes | 30320 ± 4856.3 | 59.26 | 33848 ± 2811.8 | 78.88 | 0.544 |
| Unidentified | 3504 ± 1402.1 | 6.85 | 2366 ± 260.9 | 5.51 | 0.443 |
| Proteobacteria | 697 ± 225.8 | 1.36 | 521 ± 58.5 | 1.22 | 0.468 |
| Saccharibacteria | 683 ± 224.3 | 1.34 | 121 ± 95.3 | 0.28 | 0.044 |
| Synergistetes | 1287 ± 680.6 | 2.52 | - | - | 0.088 |
| Tenericutes | 39 ± 18.3 | 0.08 | 196 ± 47.3 | 0.46 | 0.012 |
| Verrucomicrobia | 6075 ± 1334.6 | 11.87 | 2642 ± 1012.8 | 6.16 | 0.068 |
| Sequences and Index. | ERE+ | ERE− | p-Value |
|---|---|---|---|
| Number of sequences | 115,534 | 99,863 | |
| Number of operational taxonomic units (OTU) | 317 | 279 | |
| Number of identified genera | 76 | 81 | |
| Shannon Index | 6.24 (0.18) | 6.54 (0.04) | 0.001 |
| Simpson Index | 0.95 (0.04) | 0.97 (0.01) | 0.001 |
| Simpson Inverse Index | 30.12 (0.09) | 43.54 (0.06) | 0.001 |
| Fisher Index | 66.20 (2.19) | 81.40 (0.80) | 0.001 |
| Chao1 Index | 304.98 (6.70) | 275.86 (2.50) | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puón-Peláez, X.-H.D.; McEwan, N.R.; Gómez-Soto, J.G.; Álvarez-Martínez, R.C.; Olvera-Ramírez, A.M. Metataxonomic and Histopathological Study of Rabbit Epizootic Enteropathy in Mexico. Animals 2020, 10, 936. https://doi.org/10.3390/ani10060936
Puón-Peláez X-HD, McEwan NR, Gómez-Soto JG, Álvarez-Martínez RC, Olvera-Ramírez AM. Metataxonomic and Histopathological Study of Rabbit Epizootic Enteropathy in Mexico. Animals. 2020; 10(6):936. https://doi.org/10.3390/ani10060936
Chicago/Turabian StylePuón-Peláez, Xiao-Haitzi Daniel, Neil Ross McEwan, José Guadalupe Gómez-Soto, Roberto Carlos Álvarez-Martínez, and Andrea Margarita Olvera-Ramírez. 2020. "Metataxonomic and Histopathological Study of Rabbit Epizootic Enteropathy in Mexico" Animals 10, no. 6: 936. https://doi.org/10.3390/ani10060936
APA StylePuón-Peláez, X. -H. D., McEwan, N. R., Gómez-Soto, J. G., Álvarez-Martínez, R. C., & Olvera-Ramírez, A. M. (2020). Metataxonomic and Histopathological Study of Rabbit Epizootic Enteropathy in Mexico. Animals, 10(6), 936. https://doi.org/10.3390/ani10060936