Population Density, Size Structure, and Reproductive Cycle of the Comestible Sea Urchin Sphaerechinus granularis (Echinodermata: Echinoidea) in the Pagasitikos Gulf (Aegean Sea)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Field Sampling
2.2. Population Density and Size Structure
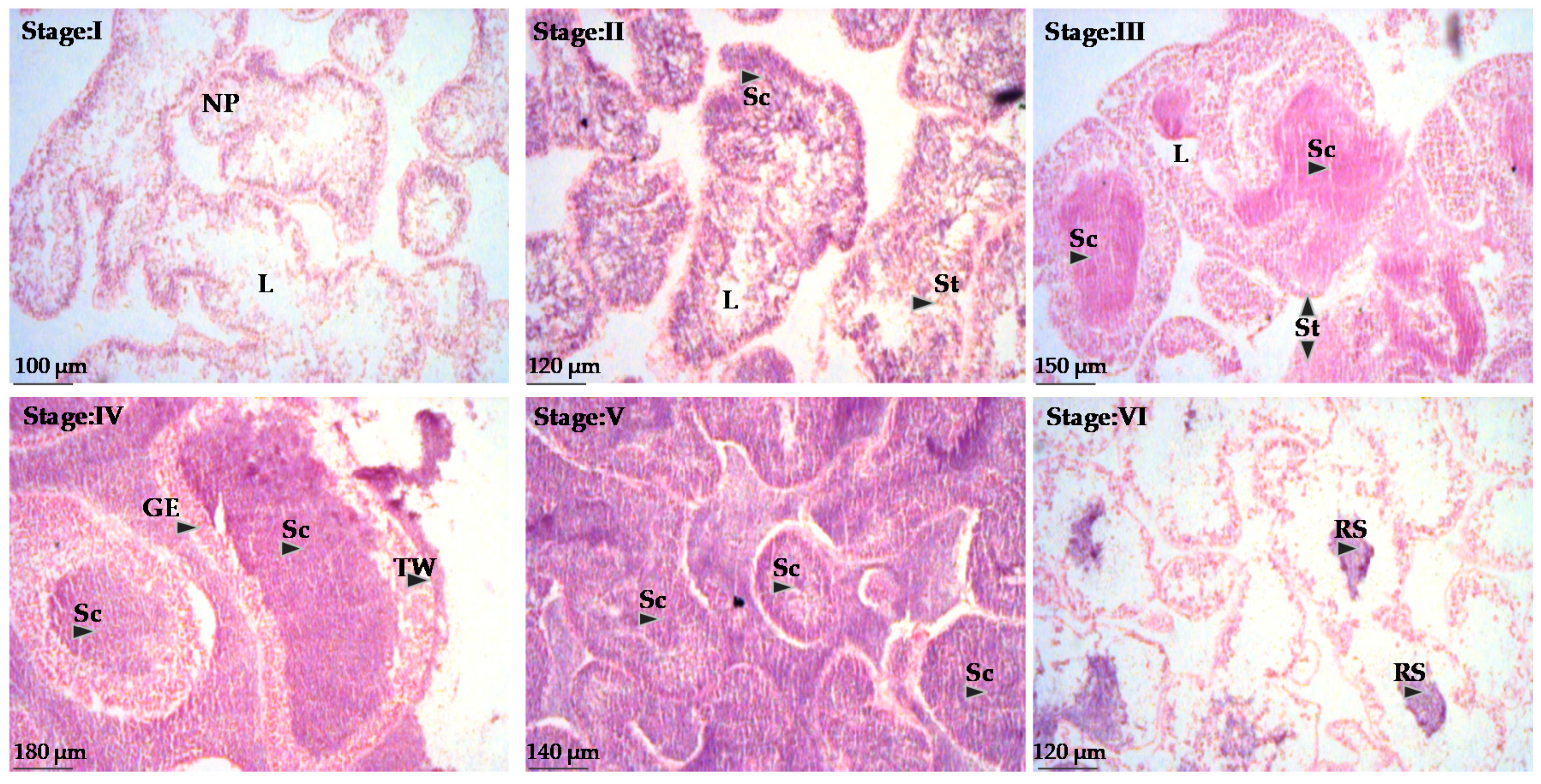
2.3. Reproductive Cycle
3. Results
3.1. Environmental Parameters
3.2. Population Density and Size Structure
3.3. Reproductive Cycle
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martínez-Pita, I.; Sánchez-Espaňa, A.I.; García, F.J. Gonadal growth and reproduction in the sea urchin Sphaerechinus granularis (Lamarck 1816) (Echinodermata: Echinoidea) in southern Spain. Sci. Mar. 2008, 72, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Palacín, C.; Turon, X.; Ballesteros, M.; Giribet, G.; López, S. Stock evaluation of three littoral echinoid species on the Catalan coast (north-western Mediterranean). P.S.Z.N. Mar. Ecol. 1998, 19, 163–177. [Google Scholar] [CrossRef]
- Sartoretto, S.; Francour, P. Quantification of bioerosion by Sphaerechinus granularis on “coralligene” concretions of the western Mediterranean. J. Mar. Biol. Assoc. UK 1997, 77, 565–568. [Google Scholar] [CrossRef]
- Verlaque, M. Preliminary data on some Posidonia feeders. Rap. Com. Int. Mer. Médit. 1981, 27, 201–202. [Google Scholar]
- Guillou, M.; Lumingas, L.J.L. The reproductive cycle of the “blunt” sea urchin. Aquac. Int. 1998, 6, 147–160. [Google Scholar] [CrossRef]
- Cherbonnier, G. Faune Marine des Pyrénées Orientales. Fasc. 2; Echinodermes, Université de Paris et Laboratoire Arago: Paris, France, 1958. (In French) [Google Scholar]
- Antoniadou, C.; Vafidis, D. Population ecology of common sea urchins (Arbacia lixula, Paracentrotus lividus, Sphaerechinus granularis) on algal-dominated rocky shores in the Aegean Sea. In Echinoderms: Ecology, Habitats and Reproductive Biology, 1st ed.; Whitmore, E., Ed.; NOVA Science: New York, NY, USA, 2014; pp. 146–166. [Google Scholar]
- Ziegenhorn, M.A. Sea urchins covering behaviour: A comparative review. In Sea Urchin from Environment to Aquaculture and Biomedicine, 1st ed.; Agnelo, M., Ed.; Intech Open: London, UK, 2017; pp. 19–33. [Google Scholar] [CrossRef] [Green Version]
- Guillou, M.; Michel, C. Reproduction and growth of Sphaerechinus granularis (Echinodermata: Echinoidea) in southern Brittany. J. Mar. Biol. Assoc. UK 1993, 73, 179–192. [Google Scholar] [CrossRef]
- Guillou, M.; Lumingas, L.J.L. Variation in the reproductive strategy of the sea urchin Sphaerechinus granularis (Echinodermata: Echinoidea) related to food availability. J. Mar. Biol. Assoc. UK 1999, 79, 131–136. [Google Scholar] [CrossRef]
- Fenaux, L. Modalitésιs de ponte chez l’oursin Sphaerechinus granularis. Revue Ges. Hydrobiol. 1972, 57, 551–558. [Google Scholar] [CrossRef]
- Koukouras, A.; Sinis, A.I.; Bobori, D.; Kazantzidis, S.; Kitsos, M.S. The echinoderm, (Deuterostomia) fauna of the Aegean Sea, and comparisons with those of the neighboring seas. J. Biol. Res. 2007, 7, 67–92. [Google Scholar]
- Korres, G.; Triantafyllou, G.; Petihakis, G.; Raitsos, D.E.; Hoteit, I.; Pollani, A.; Colella, S.; Tsiaras, K. A data assimilation tool for the Pagasitikos Gulf ecosystem dynamics: Methods and benefits. J. Mar. Syst. 2012, 94, 102–117. [Google Scholar] [CrossRef]
- Lolas, A.; Antoniadou, C.; Vafidis, D. Spatial variation of molluscan fauna associated with Cystoseira assemblages from a semi-enclosed gulf in the Aegean Sea. Reg. Stud. Mar. Sci. 2018, 19, 17–24. [Google Scholar] [CrossRef]
- Antoniadou, C.; Vafidis, D. Population structure and morphometric relationships of Paracentrotus lividus (Echinodermata: Echinoidae) in the south Aegean Sea. Cah. Biol. Mar. 2009, 50, 293–301. [Google Scholar]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sampling: Estimating Abundance of Biological Populations, 1st ed.; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Buckland, S.T.; Rexstad, E.A.; Marques, T.A.; Oedekoven, C.S. Distance Sampling: Methods and Applications, 1st ed.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Thomas, L.; Buckland, S.T.; Rexstad, E.A.; Laake, J.L.; Strindberg, S.; Hedley, S.L.; Bishop, J.R.B.; Marques, T.A.; Burnham, K.P. Distance software: Design and analysis of distance sampling surveys for estimating population size. J. Appl. Ecol. 2010, 47, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsanevakis, S. Estimating abundance of endangered marine benthic species using Distance Sampling through SCUBA diving: The Pinna nobilis (Mollusca: Bivalvia) example. In Endangered Species: New Research, 1st ed.; Columbus, A.M., Kuznetsov, L., Eds.; NOVA Science: New York, NY, USA, 2009; pp. 81–115. [Google Scholar]
- Thanopoulou, Z.; Sini, M.; Vatikiotis, K.; Katsoupis, C.; Dimitrakopoulos, P.G.; Katsanevakis, S. How many fish? Comparison of two underwater visual sampling methods for monitoring fish communities. Peer J. 2018, 6, e5066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underwood, A.J. Experiments in Ecology. Their Logical Design and Interpretation Using Analysis of Variance, 2nd ed.; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Byrne, M. Annual reproductive cycles of the commercial sea urchin Paracentrotus lividus from an exposed intertidal and a sheltered subtidal habitat on the west coast of Ireland. Mar. Biol. 1990, 104, 275–289. [Google Scholar] [CrossRef]
- Soualili, D.L.; Guillou, M.; Semroud, R. Age and growth of the echinoid Sphaerechinus granularis from the Algerian coast. J. Mar. Biol. Assoc. UK 1999, 79, 1139–1140. [Google Scholar] [CrossRef]
- Marques, T.A.; Buckland, S.T.; Borchers, D.L.; Rexstad, E.A.; Thomas, L. Distance sampling. In International Encyclopedia of Statistical Science, 1st ed.; Lovric, M., Ed.; Springer: Berlin, Germany, 2011; pp. 398–400. [Google Scholar]
- Trenzado, C.E.; Hidalgo, F.; Villanueva, M.; Furné, M.; Díaz-Casado, M.E.; Merino, R.; Sanz, A. Study of the enzymatic digestive profile in three species of Mediterranean sea urchins. Aquaculture 2012, 344, 174–180. [Google Scholar] [CrossRef]
- Ruitton, S.; Verlaque, M.; Aubin, G.; Boudouresque, C.F. Grazing on Caulerpa racemosa var. cylindracea (Caulerpales, Chlorophyta) in the Mediterranean Sea by herbivorous fishes and sea urchins. Vie Milieu 2006, 56, 33–41. [Google Scholar]
- Jordana, E.; Guillou, M.; Lumingas, L.J.L. Age and growth of the sea urchin Sphaerechinus granularis in southern Brittany. J. Mar. Biol. Assoc. UK 1997, 77, 1199–1212. [Google Scholar] [CrossRef]
- Guillou, M.; Lumingas, L.J.L.; Michel, C. The effect of feeding or starvation on resource allocation to body components during the reproductive cycle of the sea urchin Sphaerechinus granularis (Lamarck). J. Exp. Mar. Biol. Ecol. 2000, 245, 183–196. [Google Scholar] [CrossRef]
- Lozano, J.; Galera, J.; López, S.; Turon, X.; Palacín, C.; Morera, G. Biological cycles and recruitment of Paracentrotus lividus (Echinodermata: Echinoidea) in two contrasting habitats. Mar. Ecol. Prog. Ser. 1995, 122, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Guettaf, M.; San Martin, G.A.; Francour, P. Interpopulation variability of the reproductive cycle of Paracentrotus lividus (Echinodermata: Echinoidea) in the south-western Mediterranean. J. Mar. Biol. Assoc. UK 2000, 80, 899–907. [Google Scholar] [CrossRef]
- Vafidis, D.; Antoniadou, C.; Kyriakouli, K. Reproductive Cycle of the Edible Sea Urchin Paracentrotus lividus (Echinodermata: Echinoidae) in the Aegean Sea. Water 2019, 11, 1029. [Google Scholar] [CrossRef] [Green Version]
- Pearse, J.S.; Cameron, R.A. Echinodermata: Echinoidea. In Reproduction of Marine Invertebrates, 1st ed.; Giese, A.C., Pearse, J.S., Pearse, V.B., Eds.; The Boxwood Press: Pacific Grove, CA, USA, 1991; Volume 6, pp. 513–662. [Google Scholar]
- Carey, N.; Harianto, J.; Byrne, M. Sea urchins in a high-CO2 world: Partitioned effects of body size, ocean warming and acidification on metabolic rate. J. Exp. Biol. 2016, 219, 1178–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month/Parameter | T | S | pH | O2 | ||||
|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | |
| January | 13.96 | 13.17 | 36.99 | 38.21 | 8.27 | 8.33 | 5.21 | 4.01 |
| February | 13.28 | 13.46 | 37.67 | 38.03 | 8.28 | 8.27 | 3.04 | 5.11 |
| March | 13.31 | 13.53 | 37.78 | 38.46 | 8.23 | 8.29 | 4.76 | 6.49 |
| April | 15.34 | 15.94 | 38.33 | 37.94 | 8.28 | 8.28 | 6.62 | 6.49 |
| May | 21.21 | 19.92 | 37.54 | 37.54 | 8.29 | 8.26 | 5.88 | 2.48 |
| June | 25.04 | 26.45 | 37.47 | 36.64 | 8.26 | 8.24 | 6.61 | 5.71 |
| July | 27.08 | 27.12 | 36.46 | 36.18 | 8.26 | 8.23 | 5.44 | 2.45 |
| August | 27.67 | 26.78 | 36.12 | 36.76 | 8.26 | 8.31 | 2.06 | 5.02 |
| September | 24.28 | 25.03 | 36.84 | 36.49 | 8.29 | 8.29 | 4.92 | 5.92 |
| October | 21.79 | 22.04 | 36.42 | 36.84 | 8.31 | 8.35 | 5.22 | 5.54 |
| November | 18.31 | 17.95 | 36.71 | 36.99 | 8.34 | 8.39 | 2.48 | 3.67 |
| December | 16.01 | 15.64 | 36.87 | 36.21 | 8.31 | 8.24 | 2.99 | 4.97 |
| Station | Original Field Data Individuals/100 m2 | Distance Software Data Individuals/100 m2 |
|---|---|---|
| S1 | 5.5 | 9.2 |
| S2 | 4.5 | 7.8 |
| Mean value | 5.0 | 8.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vafidis, D.; Antoniadou, C.; Ioannidi, V. Population Density, Size Structure, and Reproductive Cycle of the Comestible Sea Urchin Sphaerechinus granularis (Echinodermata: Echinoidea) in the Pagasitikos Gulf (Aegean Sea). Animals 2020, 10, 1506. https://doi.org/10.3390/ani10091506
Vafidis D, Antoniadou C, Ioannidi V. Population Density, Size Structure, and Reproductive Cycle of the Comestible Sea Urchin Sphaerechinus granularis (Echinodermata: Echinoidea) in the Pagasitikos Gulf (Aegean Sea). Animals. 2020; 10(9):1506. https://doi.org/10.3390/ani10091506
Chicago/Turabian StyleVafidis, Dimitrios, Chryssanthi Antoniadou, and Vassiliki Ioannidi. 2020. "Population Density, Size Structure, and Reproductive Cycle of the Comestible Sea Urchin Sphaerechinus granularis (Echinodermata: Echinoidea) in the Pagasitikos Gulf (Aegean Sea)" Animals 10, no. 9: 1506. https://doi.org/10.3390/ani10091506
APA StyleVafidis, D., Antoniadou, C., & Ioannidi, V. (2020). Population Density, Size Structure, and Reproductive Cycle of the Comestible Sea Urchin Sphaerechinus granularis (Echinodermata: Echinoidea) in the Pagasitikos Gulf (Aegean Sea). Animals, 10(9), 1506. https://doi.org/10.3390/ani10091506