Endocrine Fertility Parameters—Genomic Background and Their Genetic Relationship to Boar Taint in German Landrace and Large White

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Phenotypes
2.2. Boar Taint
2.3. Hormone Profiling
2.4. Statistical Analyses
2.5. Genotype Data
3. Results
3.1. Descriptive Summary
3.2. Variance Component Estimation
3.3. GWAS
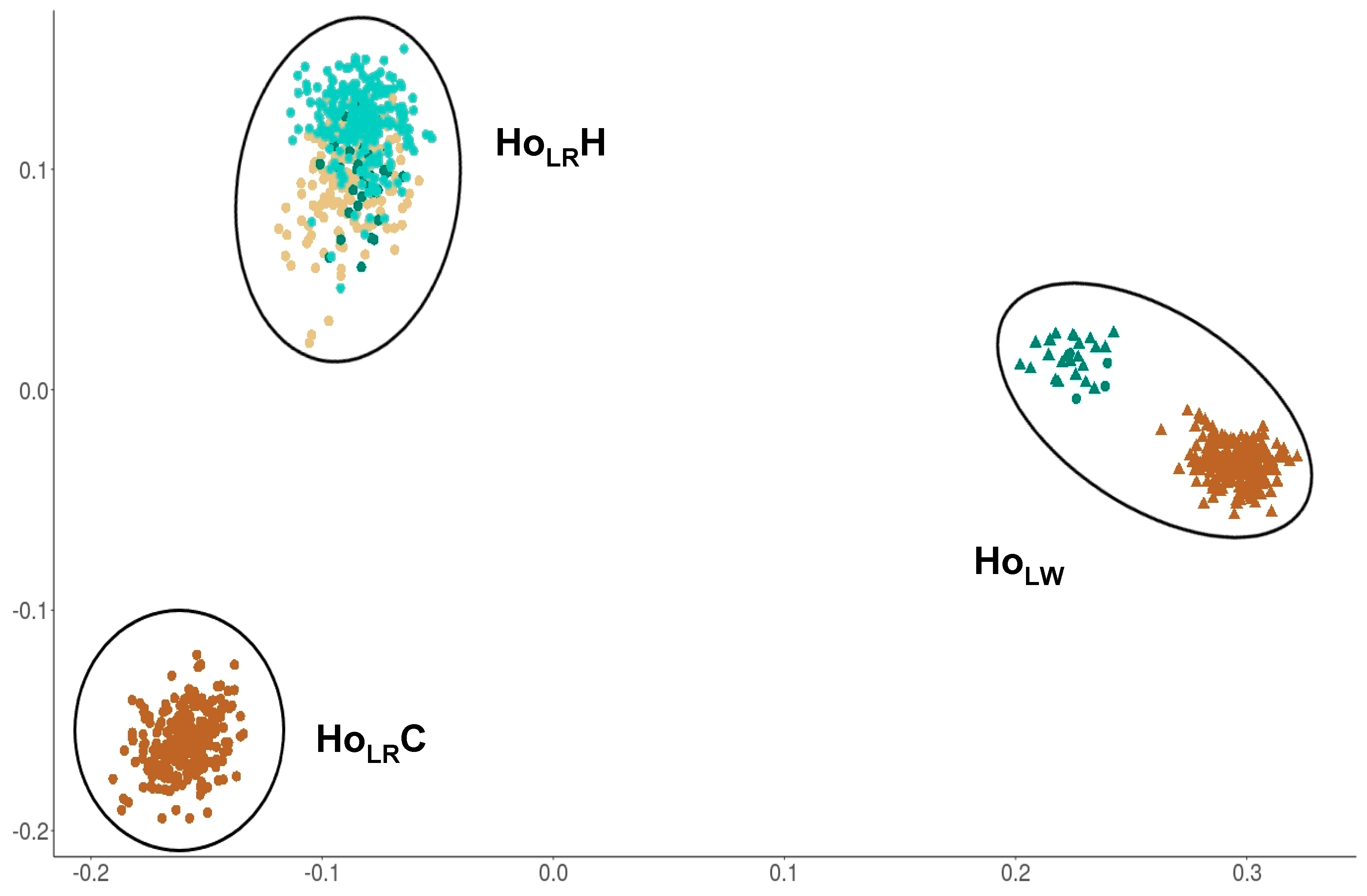
3.3.1. Genetic Structure
3.3.2. GWAS for Hormone Profiles
3.3.3. GWAS for AND and SKA
4. Discussion
4.1. Descriptive Summary for Boar Taint and Endocrine Parameters
4.2. Variance Component Estimation
4.3. GWAS
4.3.1. GWAS Hormones
4.3.2. GWAS Boar Taint Cluster BTLRH
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3’PUTR | 3’prime untranslated region |
| AKAP12 | A-Kinase Anchoring Protein12 |
| AND | Androstenone |
| BTLRC | Boar taint cluster Landrace, commercial breeding organization |
| BTLRH | Boar taint cluster Landrace, herd book organizations |
| BTLWC | Boar taint cluster Large White, commercialbreeding organization |
| BTLWH | Boar taint cluster Large White, herd book organizations |
| C | Commercial breeding organization |
| CORT | Cortisol |
| CYP2E1 | Cytochrome P450 Family 2 Subfamily E Member 1 |
| DDX24 | DEAD-Box Helicase 24 |
| DGV | Downstream gene variant |
| EFP | Endocrine fertility parameters |
| EST | 17-β estradiol |
| FSH | Follicle-stimulating hormone |
| GC | Genomic Control |
| GWAS | Genome-wide association analysis |
| GZMK | Granzyme K |
| H | Herd book organizations |
| h2 | Heritability |
| HoLRC | Hormone cluster Landrace, commercial breeding organization |
| HoLRH | Hormone cluster Landrace, herd book organizations |
| HoLW | Hormone cluster Large White from all organizations |
| HPA axis | Hypothalamic-pituitary-adrenal axis |
| IL6ST | interleukin-6 receptor subunit beta |
| intra-CV | Intra-variation coefficient |
| kg | Kilogram |
| LH | Luteinizing hormone |
| LR | Landrace |
| LW | Large White |
| MAF | Minor allele frequency |
| ml | Milliliter |
| n.m. | Not mapped |
| NCTEV | non coding transcript exon variant |
| ng | Nanogram |
| NR3C1 | Nuclear Receptor Subfamily 3 Group C Member 1 |
| PHACTR1 | Phosphatase And Actin Regulator 1 |
| PROG | Progesterone |
| QTL | Quantitative trait locus/loci |
| rg | Genetic correlation |
| rp | Phenotypic correlation |
| SIDA-HSPM-GC/MS | Stable isotope dilution analysis-headspacesolid-phase microextraction-gas chromatography/mass spectrometry |
| SKA | Skatole |
| SLC2A5 | Solute Carrier Family 2 Member 5 |
| SNPs | Single nucleotide polymorphism(s) |
| SSC | Susscrofachromosome |
| TEST | Testosterone |
| UGV | Upstream gene variant |
| VCE | Variance component estimation |
| VRK1 | VRK Serine/Threonine Kinase 1 |
References
- Merks, J.W.M.; Mathur, P.K.; Knol, E.F. New phenotypes for new breeding goals in pigs. Animal 2012, 6, 535–543. [Google Scholar] [CrossRef] [Green Version]
- Drittes Gesetz zur Änderung des Tierschutzgesetzes. Gesetz über Die Zusammenarbeit von Bundesregierung und Deutschem Bundestag in Angelegenheiten der Europäischen Union (EUZBBG); Bundesanzeiger Verlag: Köln, Germany, 2013; pp. 2182–2196. [Google Scholar]
- Lundström, K.; Matthews, K.R.; Haugen, J.-E. Pig meat quality from entire males. Animal 2009, 3, 1497–1507. [Google Scholar] [CrossRef] [Green Version]
- Stefanski, V.; Labussière, E.; Millet, S.; Weiler, U. Pork production with immunocastration: Welfare and environment. In Proceedings of the EAAP 69th Annual Meeting of the European Federation of Animal Science, Dubrovnik, Croatia, 27–31 August 2018; Wageningen Academic Publishers: Wageningen, The Netherlands, 2018; p. 229. [Google Scholar]
- Zamaratskaia, G.; Squires, E.J. Biochemical, nutritional and genetic effects on boar taint in entire male pigs. Animal 2009, 3, 1508–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, L.L. Reproductive Biology of Pigs. Anim. Ind. Rep. 2009. [Google Scholar] [CrossRef]
- Weiler, U.; Wesoly, R. Physiologische Aspekte der Androstenon- und Skatolbildung beim Eber. Züchtungskunde 2012, 84, 365–393. [Google Scholar]
- Squires, E.J. Possibilites for selection against boar taint. Acta Vet. Scand. 2006, 48, S8. [Google Scholar] [CrossRef]
- Bonneau, M. Compounds responsible for boar taint, with special emphasis on androstenone: A review. Livest. Prod. Sci. 1982, 9, 687–705. [Google Scholar] [CrossRef]
- Strathe, A.B.; Velander, I.H.; Mark, T.; Kadarmideen, H.N. Genetic parameters for androstenone and skatole as indicators of boar taint and their relationship to production and litter size traits in Danish Landrace. J. Anim. Sci. 2013, 91, 2587–2595. [Google Scholar] [CrossRef] [Green Version]
- Tajet, H.; Andresen, Ø.; Meuwissen, T.H. Estimation of genetic parameters of boar taint; skatole and androstenone and their correlations with sexual maturation. Acta Vet. Scand. 2006, 48, S9. [Google Scholar] [CrossRef]
- Windig, J.J.; Mulder, H.A.; Napel, J.T.; Knol, E.F.; Mathur, P.K.; Crump, R.E. Genetic parameters for androstenone, skatole, indole, and human nose scores as measures of boar taint and their relationship with finishing traits. J. Anim. Sci. 2012, 90, 2120–2129. [Google Scholar] [CrossRef]
- Brinke, I.; Große-Brinkhaus, C.; Roth, K.; Pröll-Cornelissen, M.J.; Henne, H.; Schellander, K.; Tholen, E. Genomic background and genetic relationships between boar taint and fertility traits in German Landrace and Large White. BMC Genet. 2020, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Doran, E.; Whittington, F.W.; Wood, J.D.; McGivan, J.D. Cytochrome P450IIE1 (CYP2E1) is induced by skatole and this induction is blocked by androstenone in isolated pig hepatocytes. Chem. Biol. Interact. 2002, 140, 81–92. [Google Scholar] [CrossRef]
- Moe, M.; Lien, S.; Aasmundstad, T.; Meuwissen, T.H.E.; Hansen, M.H.S.; Bendixen, C.; Grindflek, E. Association between SNPs within candidate genes and compounds related to boar taint and reproduction. BMC Genet. 2009, 10, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, P.K.; Napel, J.T.; Crump, R.E.; Mulder, H.A.; Knol, E.F. Genetic relationship between boar taint compounds, human nose scores, and reproduction traits in pigs. J. Anim. Sci. 2013, 91, 4080–4089. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, A.M.; Bastiaansen, J.W.M.; Harlizius, B.; Megens, H.-J.; Madsen, O.; Crooijmans, R.P.M.A.; Groenen, M.A.M. On the relationship between an Asian haplotype on chromosome 6 that reduces androstenone levels in boars and the differential expression of SULT2A1 in the testis. BMC Genet. 2014, 15, 4. [Google Scholar] [CrossRef] [Green Version]
- Gower, D.B. 16-Unsaturated C19 Steroids a Review of Their Chemistry, Biochemistry and Possible Physiological Role. J. Steroid Biochem. 1972, 3, 45–103. [Google Scholar] [CrossRef]
- Brooks, R.I.; Pearson, A.M. Steroid hormone pathways in the pig, with special emphasis on boar odor: A review. J. Anim. Sci. 1986, 62, 632–645. [Google Scholar] [CrossRef]
- Fischer, J.; Elsinghorst, P.W.; Bücking, M.; Tholen, E.; Petersen, B.; Wüst, M. Development of a candidate reference method for the simultaneous quantitation of the boar taint compounds androstenone, 3α-androstenol, 3β-androstenol, skatole, and indole in pig fat by means of stable isotope dilution analysis-headspace solid-phase microextraction-gas chromatography/mass spectrometry. Anal. Chem. 2011, 83, 6785–6791. [Google Scholar] [CrossRef]
- Hornung, R.W.; Reed, L.D. Estimation of Average Concentration in the Presence of Nondetectable Values. Appl. Occup. Environ. Hyg. 1990, 5, 46–51. [Google Scholar] [CrossRef]
- Meyer, K. WOMBAT: A tool for mixed model analyses in quantitative genetics by restricted maximum likelihood (REML). J. Zhejiang Univ. Sci. B 2007, 8, 815–821. [Google Scholar] [CrossRef] [Green Version]
- Aulchenko, Y.S.; Ripke, S.; Isaacs, A.; van Duijn, C.M. GenABEL: An R library for genome-wide association analysis. Bioinformatics 2007, 23, 1294–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devlin, B.; Roeder, K. Genomic Control for Association Studies. Biometrics 1999, 55, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Kendall, M.G.; Stuart, A.; Ord, J.K. The Advanced Theory of Statistics, 4th ed.; Distribution Theory; Griffin: London, UK, 1977; Volume 1, ISBN 0852642423. [Google Scholar]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G.; et al. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef] [PubMed]
- Bunter, K.L.; Bennett, C.; Luxford, B.G.; Graser, H.-U. Sire breed comparisons for meat and eating quality traits in Australian pig populations. Animal 2008, 2, 1168–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kress, K.; Weiler, U.; Schmucker, S.; Čandek-Potokar, M.; Vrecl, M.; Fazarinc, G.; Škrlep, M.; Batorek-Lukač, N.; Stefanski, V. Influence of Housing Conditions on Reliability of Immunocastration and Consequences for Growth Performance of Male Pigs. Animals 2019, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Colenbrander, B.; de Jong, F.H.; Wensing, C.J. Changes in serum testosterone concentrations in the male pig during development. J. Reprod. Fertil. 1978, 53, 377–380. [Google Scholar] [CrossRef]
- Cheon, Y.M.; Kim, H.K.; Yang, C.B.; Yi, Y.J.; Park, C.S. Effect of Season Influencing Semen Characteristics, Frozen-Thawed Sperm Viability and Testosterone Concentration in Duroc Boars. Asian-Aust. J. Anim. Sci. 2002, 15, 500–503. [Google Scholar] [CrossRef]
- Zoels, S.; Reiter, S.; Ritzmann, M.; Weiß, C.; Numberger, J.; Schütz, A.; Lindner, P.; Stefanski, V.; Weiler, U. Influences of Immunocastration on Endocrine Parameters, Growth Performance and Carcass Quality, as Well as on Boar Taint and Penile Injuries. Animals 2020, 10, 346. [Google Scholar] [CrossRef] [Green Version]
- Booth, W.D. A study of some major testicular steroids in the pig in relation to their effect on the development of male characteristics in the prepubertally castrated boar. J. Reprod. Fertil. 1980, 59, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Wettemann, R.P.; Johnson, R.K.; Omtvedt, I.T. The relationships between plasma progesterone and lh and the numbers of corpora lutea and embryos in purebred and crossbred gilts. J. Anim. Sci. 1980, 51, 1352–1357. [Google Scholar] [CrossRef] [Green Version]
- Henricks, D.M.; Guthrie, H.D.; Handlin, D.L. Plasma estrogen, progesterone and luteinizing hormone levels during the estrous cycle in pigs. Biol. Reprod. 1972, 6, 210–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoving, L.L.; Haen, S.M.; Laurenssen, B.F.A.; Peltoniemi, O.A.T.; Kemp, B.; Soede, N.M. Caudal vena cava progesterone and LH release patterns on Day 14 of gestation in primiparous sows. Reprod. Fertil. Dev. 2017, 29, 476–481. [Google Scholar] [CrossRef] [PubMed]
- van Wettere, W.H.E.J.; Toplis, P.; Miller, H.M. Effect of oral progesterone and caffeine at the end of gestation on farrowing duration and piglet growth and survival. Animal 2018, 12, 1638–1641. [Google Scholar] [CrossRef] [PubMed]
- Robic, A.; Larzul, C.; Bonneau, M. Genetic and metabolic aspects of androstenone and skatole deposition in pig adipose tissue: A review. Genet. Sel. Evol. 2008, 40, 129–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellier, P.; Le Roy, P.; Fouilloux, M.N.; Gruand, J.; Bonneau, M. Responses to restricted index selection and genetic parameters for fat androstenone level and sexual maturity status of young boars. Livest. Prod. Sci. 2000, 63, 265–274. [Google Scholar] [CrossRef]
- Engelsma, K.A.; Bergsma, R.; Harlizius, B.; Ducro-Steverink, D.W.B.; Knol, E.F. Genetic parameters for components of boar taint and their relation with carcass quality and female fertility. In Proceedings of the EAAP 58th Annual Meeting, Session 10, Poster 27, Dublin, Ireland, 26–29 August 2007. [Google Scholar]
- Zamaratskaia, G.; Babol, J.; Madej, A.; Squires, E.J.; Lundstrom, K. Age-related Variation of Plasma Concentrations of Skatole, Androstenone, Testosterone, Oestradiol-17beta, Oestrone Sulphate, Dehydroepiandrosterone Sulphate, Triiodothyronine and IGF-1 in Six Entire Male Pigs. Reprod. Domest. Anim. 2004, 39, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Grindflek, E.; Meuwissen, T.H.E.; Aasmundstad, T.; Hamland, H.; Hansen, M.H.S.; Nome, T.; Kent, M.; Torjesen, P.; Lien, S. Revealing genetic relationships between compounds affecting boar taint and reproduction in pigs. J. Anim. Sci. 2011, 89, 680–692. [Google Scholar] [CrossRef] [Green Version]
- Larzul, C.; Terenina, E.; Foury, A.; Billon, Y.; Louveau, I.; Merlot, E.; Mormede, P. The cortisol response to ACTH in pigs, heritability and influence of corticosteroid-binding globulin. Animal 2015, 9, 1929–1934. [Google Scholar] [CrossRef] [Green Version]
- Kadarmideen, H.N.; Janss, L.L.G. Population and systems genetics analyses of cortisol in pigs divergently selected for stress. Physiol. Genom. 2007, 29, 57–65. [Google Scholar] [CrossRef]
- Parois, S.P.; Prunier, A.; Mercat, M.J.; Merlot, E.; Larzul, C. Genetic relationships between measures of sexual development, boar taint, health, and aggressiveness in pigs. J. Anim. Sci. 2015, 93, 3749–3758. [Google Scholar] [CrossRef]
- Walker, S.; Robison, O.W.; Whisnant, C.S.; Cassady, J.P. Effect of divergent selection for testosterone production on testicular morphology and daily sperm production in boars. J. Anim. Sci. 2004, 82, 2259–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, W.H. Non-classical actions of testosterone and spermatogenesis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1557–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, T.; Lunstra, D.D.; Ford, J.J. Differential Pituitary and Gonadal Function of Chinese Meishan and European White Composite Boars: Effects of Gonadotropin-Releasing Hormone Stimulation, Castration, and Steroidal Feedback. Biol. Reprod. 1996, 54, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muráni, E.; Ponsuksili, S.; D’Eath, R.B.; Turner, S.P.; Kurt, E.; Evans, G.; Thölking, L.; Klont, R.; Foury, A.; Mormède, P.; et al. Association of HPA axis-related genetic variation with stress reactivity and aggressive behaviour in pigs. BMC Genet. 2010, 11, 74. [Google Scholar] [CrossRef] [Green Version]
- Muráni, E.; Reyer, H.; Ponsuksili, S.; Fritschka, S.; Wimmers, K. A substitution in the ligand binding domain of the porcine glucocorticoid receptor affects activity of the adrenal gland. PLoS ONE 2012, 7, e45518. [Google Scholar] [CrossRef] [Green Version]
- Gashaw, I.; Grümmer, R.; Klein-Hitpass, L.; Dushaj, O.; Bergmann, M.; Brehm, R.; Grobholz, R.; Kliesch, S.; Neuvians, T.P.; Schmid, K.W.; et al. Gene signatures of testicular seminoma with emphasis on expression of ets variant gene 4. Cell. Mol. Life Sci. 2005, 62, 2359–2368. [Google Scholar] [CrossRef]
- Désautés, C.; Bidanel, J.P.; Milant, D.; Iannuccelli, N.; Amigues, Y.; Bourgeois, F.; Caritez, J.C.; Renard, C.; Chevalet, C.; Mormède, P. Genetic linkage mapping of quantitative trait loci for behavioral and neuroendocrine stress response traits in pigs. J. Anim. Sci. 2002, 80, 2276–2285. [Google Scholar] [CrossRef]
- Ponsuksili, S.; Du, Y.; Murani, E.; Schwerin, M.; Wimmers, K. Elucidating molecular networks that either affect or respond to plasma cortisol concentration in target tissues of liver and muscle. Genetics 2012, 192, 1109–1122. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.R.; Ren, J.; Xing, Y.Y.; Guo, Y.M.; Wu, Y.B.; Yang, G.C.; Mao, H.R.; Huang, L.-S. A genome scan for quantitative trait loci affecting male reproductive traits in a White Duroc x Chinese Erhualian resource population. J. Anim. Sci. 2009, 87, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Maeda, A.; Goto, Y.; Matsuda-Minehata, F.; Cheng, Y.; Inoue, N.; Manabe, N. Changes in Expression of Interleukin-6 Receptors in Granulosa Cells During Follicular Atresia in Pig Ovaries. J. Reprod. Dev. 2007, 53, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Grindflek, E.; Lien, S.; Hamland, H.; Hansen, M.H.S.; Kent, M.; van Son, M.; Meuwissen, T.H.E. Large scale genome-wide association and LDLA mapping study identifies QTLs for boar taint and related sex steroids. BMC Genom. 2011, 12, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinhauser, C.B.; Landers, M.; Myatt, L.; Burghardt, R.C.; Vallet, J.L.; Bazer, F.W.; Johnson, G.A. Fructose Synthesis and Transport at the Uterine-Placental Interface of Pigs: Cell-Specific Localization of SLC2A5, SLC2A8, and Components of the Polyol Pathway. Biol. Reprod. 2016, 95, 108. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Qian, H.; Luo, Z.; Li, D.; Xu, H.; Chen, J.; He, P.; Zhou, X.; Zhang, T.; Chen, J.; et al. PHACTR1 gene polymorphism with the risk of coronary artery disease in Chinese Han population. Postgrad. Med. J. 2019, 95, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Edea, Z.; Kim, K.-S. A whole genomic scan to detect selection signatures between Berkshire and Korean native pig breeds. J. Anim. Sci. Technol. 2014, 56, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mörlein, D.; Lungershausen, M.; Steinke, K.; Sharifi, A.R.; Knorr, C. A single nucleotide polymorphism in the CYP2E1 gene promoter affects skatole content in backfat of boars of two commercial Duroc-sired crossbred populations. Meat Sci. 2012, 92, 739–744. [Google Scholar] [CrossRef]
- Skinner, T.M.; Doran, E.; McGivan, J.D.; Haley, C.S.; Archibald, A.L. Cloning and mapping of the porcine cytochrome-p450 2E1 gene and its association with skatole levels in the domestic pig. Anim. Genet. 2005, 36, 417–422. [Google Scholar] [CrossRef]
- Squires, E.J.; Lundström, K. Relationship Between Cytochrome P450IIE1 in Liver and Levels of Skatole and Its Metabolites in Intact Male Pigs. J. Anim. Sci. 1997, 75, 2506–2511. [Google Scholar] [CrossRef]
- Wiercinska, P.; Lou, Y.; Squires, E.J. The roles of different porcine cytochrome P450 enzymes and cytochrome b5A in skatole metabolism. Animal 2012, 6, 834–845. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Trait | Cluster | Number of Animals | Number of Markers |
|---|---|---|---|
| AND | BTLRC * | 1293 | 38,411 |
| BTLWC * | 1317 | 39,302 | |
| BTLRH | 735 | 43,644 | |
| SKA | BTLRC * | 1293 | 38,411 |
| BTLWC * | 1317 | 39,302 | |
| BTLRH | 735 | 43,644 | |
| CORT | HoLRC | 254 | 40,176 |
| HoLW | 271 | 40,972 | |
| HoLRH | 434 | 44,095 | |
| TEST | HoLRC | 252 | 40,176 |
| HoLW | 267 | 40,972 | |
| HoLRH | 423 | 44,095 | |
| EST | HoLRC | 251 | 40,176 |
| HoLW | 265 | 40,972 | |
| HoLRH | 415 | 44,095 | |
| LH | HoLRC | 254 | 40,176 |
| HoLW | 272 | 40,972 | |
| HoLRH | 417 | 44,095 | |
| FSH | HoLRC | 254 | 40,176 |
| HoLW | 272 | 40,972 | |
| HoLRH | 417 | 44,095 |
| Trait | Sex | Landrace | Large White | ||
|---|---|---|---|---|---|
| N | Mean ± SD | N | Mean ± SD | ||
| Age | female | 353 | 4.85 ± 3.44 | 138 | 4.83 ± 2.05 |
| male | 357 | 5.11 ± 2.49 | 148 | 5.09 ± 1.58 | |
| CORT | female | 353 | 3.34 ± 2.59 | 137 | 3.49 ± 2.71 |
| male | 357 | 3.53 ± 2.84 | 148 | 3.65 ± 3.03 | |
| male + female | 710 | 3.44 ± 2.75 | 285 | 3.58 ± 2.91 | |
| TEST | female | 344 | −0.71 ± 0.51 | 133 | −1.90 ± −2.12 |
| male | 353 | 2.44 ± 2.25 | 148 | 2.08 ± 2.02 | |
| male + female | 697 | 1.81 ± 2.17 | 281 | 1.46 ± 1.91 | |
| EST | female | 340 | −1.35 ± −1.35 | 132 | −1.56 ± −1.90 |
| male | 346 | 0.46 ± 0.41 | 147 | −0.03 ± 0.14 | |
| male + female | 686 | −0.07 ± 0.24 | 279 | −0.50 ± −0.08 | |
| LH | female | 343 | 1.95 ± 1.45 | 138 | 1.97 ± 0.89 |
| male | 357 | 1.03 ± 0.43 | 148 | 0.95 ± 0.03 | |
| male + female | 700 | 1.59 ± 1.34 | 286 | 1.58 ± 1.10 | |
| FSH | female | 348 | 7.20 ± 6.64 | 138 | 7.21 ± 6.13 |
| male | 357 | 7.25 ± 6.92 | 148 | 7.27 ± 6.64 | |
| male + female | 705 | 7.22 ± 6.80 | 286 | 7.24 ± 6.46 | |
| AND | male | 2136 | 7.41 ± 7.32 | 1639 | 7.11 ± 7.01 |
| SKA | male | 2136 | 5.40 ± 5.53 | 1639 | 4.54 ± 4.88 |
| Trait | Breed | AND | SKA | CORT | TEST | EST | LH | FSH | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AND | LR | 0.52 | (0.07) | 0.42 | (0.11) | −0.18 | (0.28) | 0.62 | (0.91) | 0.49 | (0.33) | 0.11 | (0.20) | 0.30 | (0.19) |
| LW | 0.44 | (0.07) | 0.57 | (0.12) | 0.08 | (0.26) | 0.83 | (0.34) | 0.46 | (0.27) | 0.32 | (n.E.) | 0.30 | (0.25) | |
| SKA | LR | 0.34 | 0.40 | (0.06) | −0.21 | (0.30) | 0.93 | (n.E.) | 0.95 | (0.40) | −0.16 | (0.21) | −0.14 | (0.20) | |
| LW | 0.27 | 0.34 | (0.07) | 0.38 | (0.30) | 0.27 | (0.36) | 0.03 | (0.30) | 0.45 | (n.E.) | 0.01 | (0.29) | ||
| CORT | LR | −0.01 | −0.01 | 0.11 | (0.08) | −0.35 | (n.E.) | 0.01 | (0.60) | −0.27 | (0.42) | 0.03 | (0.37) | ||
| LW | 0.02 | 0.02 | 0.35 | (0.17) | −0.03 | (0.48) | −0.26 | (0.41) | −0.42 | (n.E.) | −0.58 | (0.38) | |||
| TEST | LR | 0.32 | 0.28 | −0.02 | 0.03 | (0.08) | 0.89 | (n.E.) | 0.11 | (0.86) | 0.11 | (0.72) | |||
| LW | 0.52 | 0.29 | 0.12 | 0.23 | (0.18) | 0.58 | (0.34) | 0.01 | (n.E.) | −0.06 | (0.45) | ||||
| EST | LR | 0.36 | 0.41 | 0.06 | 0.65 | 0.09 | (0.08) | −0.22 | (0.42) | −0.13 | (0.42) | ||||
| LW | 0.47 | 0.27 | 0.04 | 0.76 | 0.42 | (0.25) | −0.04 | (n.E.) | 0.17 | (0.39) | |||||
| LH | LR | 0.10 | −0.03 | 0.07 | −0.32 | −0.16 | 0.28 | (0.10) | 0.91 | (0.17) | |||||
| LW | 0.15 | 0.12 | 0.07 | 0.24 | 0.15 | 0.04 | (0.19) | 0.66 | (n.E.) | ||||||
| FSH | LR | 0.14 | 0.07 | −0.08 | 0.14 | 0.17 | 0.42 | 0.28 | (0.09) | ||||||
| LW | 0.20 | 0.03 | −0.04 | 0.15 | 0.13 | 0.52 | 0.37 | (0.17) | |||||||
| CORT | TEST | EST | FSH | LH | |
|---|---|---|---|---|---|
| Genetic correlation | 0.80 (n.E.) | 0.91 (n.E.) | 0.42 (n.E.) | 0.75 (±0.59) | 0.88 (±0.43) |
| Trait | Cluster | SNP Name | SSC | Position (Mb) | MAF | Variant | Variance by SNP | SNP Effect (± SE) | Effect Allele | Gene Symbol |
|---|---|---|---|---|---|---|---|---|---|---|
| CORT | HoLRC | ALGA0106239 | 2 | 144.8 | 0.15 | Intron | 0.088 | −0.314 (±0.06) | A | NR3C1 |
| DRGA0017574 | 2 | 144.8 | 0.15 | Intron | 0.088 | −0.314 (±0.06) | T | NR3C1 | ||
| FBF0920 * | 7 | 115.2 | 0.24 | - | 0.087 | 0.274 (±0.05) | - | - | ||
| CASI0004483 * | 7 | 115.3 | 0.17 | Intron | 0.115 | 0.374 (±0.06) | A | DDX24 | ||
| ALGA0045097 | 7 | 115.6 | 0.51 | Intron | 0.071 | 0.202 (±0.04) | A | alpha-1-antiproteinase-like | ||
| FBF0965 | 7 | 115.6 | 0.51 | - | 0.069 | 0.199 (±0.04) | - | - | ||
| HoLW | FBF0971 * | 7 | 115.6 | 0.18 | - | 0.123 | 0.375 (±0.06) | - | - | |
| H3GA0023283 | 7 | 115.6 | 0.72 | Intron | 0.077 | −0.233 (±0.05) | G | SERPINA11 | ||
| FBF0974 | 7 | 115.6 | 0.72 | - | 0.077 | −0.233 (±0.05) | - | - | ||
| MARC0043760* | 7 | 115.6 | 0.78 | Intron | 0.109 | −0.326 (±0.06) | G | SERPINA11 | ||
| FBF0973 | 7 | - | 0.71 | - | 0.076 | −0.231 (±0.05) | - | - | ||
| HoLRH | DIAS0003615 | 18 | 33.3 | 0.35 | n.m. | 0.038 | −0.137 (±0.03) | G | - | |
| TEST | HoLRC | ASGA0001286 | 1 | 14.9 | 0.57 | Intron | 0.081 | 0.183 (±0.04) | A | AKAP12 |
| DRGA0000172 | 1 | 15.0 | 0.57 | Intron | 0.081 | 0.183 (±0.04) | T | AKAP12 | ||
| ALGA0001286 | 1 | 15.0 | 0.43 | intergenic | 0.082 | −0.184 (±0.04) | C | - | ||
| ASGA0001297 | 1 | 150.6 | 0.43 | intergenic | 0.082 | −0.184 (±0.04) | C | - | ||
| ALGA0044414 | 7 | 106.5 | 0.04 | intergenic | 0.084 | 0.487 (±0.10) | T | - | ||
| INRA0028035 | 7 | 113.1 | 0.04 | n.m. | 0.084 | 0.487 (±0.10) | C | - | ||
| HoLW | ASGA0073034 | 16 | 33.9 | 0.40 | intergenic | 0.074 | 0.250 (±0.05) | G | - | |
| ASGA0073036 | 16 | 34.0 | 0.40 | intergenic | 0.074 | 0.250 (±0.05) | A | - | ||
| MARC0056521 | 16 | 34.2 | 0.39 | UGV | 0.079 | 0.263 (±0.06) | G | Granzyme K | ||
| ALGA0116942 | 16 | 34.7 | 0.37 | Intron | 0.069 | 0.242 (±0.05) | C | PLPP1 | ||
| ASGA0073065 | 16 | 35.0 | 0.37 | Intron | 0.068 | 0.241 (±0.05) | G | IL31RA | ||
| ALGA0090291 | 16 | 35.1 | 0.37 | Intron | 0.068 | 0.238 (±0.05) | T | IL6ST | ||
| ASGA0096589 | 16 | 35.3 | 0.37 | Intron | 0.068 | 0.241 (±0.05) | C | ANKRD55 | ||
| SIRI0000852 | 16 | 37.0 | 0.41 | n.m. | 0.078 | 0.249 (±0.05) | A | - | ||
| EST | HoLRH | MARC0051573 | 6 | 69.5 | 0.39 | Intron | 0.044 | −0.274 (±0.06) | T | SLC2A5 |
| ASGA0075694 | 17 | 19.6 | 0.10 | NCTEV | 0.048 | 0.445 (±0.10) | T | ENSSSCG00000048560 | ||
| LH | HoLRH | ALGA0038510 | 7 | 9.5 | 0.21 | Intron | 0.043 | −0.159 (±0.04) | A | PHACTR1 |
| FSH | HoLRC | MARC0079871 | 10 | 58.1 | 0.01 | intergenic | 0.069 | 0.628 (±0.14) | C | - |
| HoLW | DBNP0002208 | 7 | 117.9 | 0.19 | Intron | 0.065 | 0.146 (±0.03) | C | VRK1 | |
| SKA | BTLRH | M1GA0020074 | 14 | 140.5 | 0.69 | UGV | 0.035 | −0.233 (±0.04) | A | LRRC27 |
| MARC0028756 | 14 | 140.6 | 0.69 | Intron | 0.035 | −0.233 (±0.04) | A | LRRC27 | ||
| M1GA0020080 | 14 | 140.6 | 0.27 | UGV | 0.035 | 0.244 (±0.05) | C | LRRC27, PWWP2B | ||
| M1GA0020121 | 14 | 140.9 | 0.27 | 3’PUTR | 0.035 | 0.242 (±0.05) | T | CFAP46 | ||
| M1GA0020138 | 14 | 141.0 | 0.27 | DGV | 0.033 | 0.238 (±0.05) | G | ENSSSCG00000047411 | ||
| ALGA0083389 | 14 | 141.1 | 0.27 | DGV | 0.034 | 0.239 (±0.05) | T | ADGRA1 | ||
| INRA0048622 | 14 | 141.1 | 0.27 | Intron | 0.034 | 0.239 (±0.05) | T | KNDC1 | ||
| ASGA0068302 | 14 | 141.2 | 0.27 | UGV | 0.034 | 0.239 (±0.05) | G | ADAM8, TUBGCP2 | ||
| H3GA0043620 | 14 | 141.2 | 0.27 | intergenic | 0.034 | 0.239 (±0.05) | G | - | ||
| ASGA0068308 | 14 | 141.3 | 0.27 | Intron | 0.034 | 0.239 (±0.05) | G | CALY | ||
| H3GA0043634 | 14 | 141.3 | 0.27 | 3’PUTR | 0.034 | 0.239 (±0.05) | T | ECHS1 | ||
| H3GA0043632 | 14 | 141.3 | 0.27 | Intron | 0.034 | 0.239 (±0.05) | A | MTG1 | ||
| INRA0048614 | 14 | 152.9 | 0.27 | n.m. | 0.034 | 0.239 (±0.05) | G | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brinke, I.; Große-Brinkhaus, C.; Roth, K.; Pröll-Cornelissen, M.J.; Klein, S.; Schellander, K.; Tholen, E. Endocrine Fertility Parameters—Genomic Background and Their Genetic Relationship to Boar Taint in German Landrace and Large White. Animals 2021, 11, 231. https://doi.org/10.3390/ani11010231
Brinke I, Große-Brinkhaus C, Roth K, Pröll-Cornelissen MJ, Klein S, Schellander K, Tholen E. Endocrine Fertility Parameters—Genomic Background and Their Genetic Relationship to Boar Taint in German Landrace and Large White. Animals. 2021; 11(1):231. https://doi.org/10.3390/ani11010231
Chicago/Turabian StyleBrinke, Ines, Christine Große-Brinkhaus, Katharina Roth, Maren Julia Pröll-Cornelissen, Sebastian Klein, Karl Schellander, and Ernst Tholen. 2021. "Endocrine Fertility Parameters—Genomic Background and Their Genetic Relationship to Boar Taint in German Landrace and Large White" Animals 11, no. 1: 231. https://doi.org/10.3390/ani11010231
APA StyleBrinke, I., Große-Brinkhaus, C., Roth, K., Pröll-Cornelissen, M. J., Klein, S., Schellander, K., & Tholen, E. (2021). Endocrine Fertility Parameters—Genomic Background and Their Genetic Relationship to Boar Taint in German Landrace and Large White. Animals, 11(1), 231. https://doi.org/10.3390/ani11010231