A Potential Atypical Case of Rabbit Haemorrhagic Disease in a Dwarf Rabbit

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
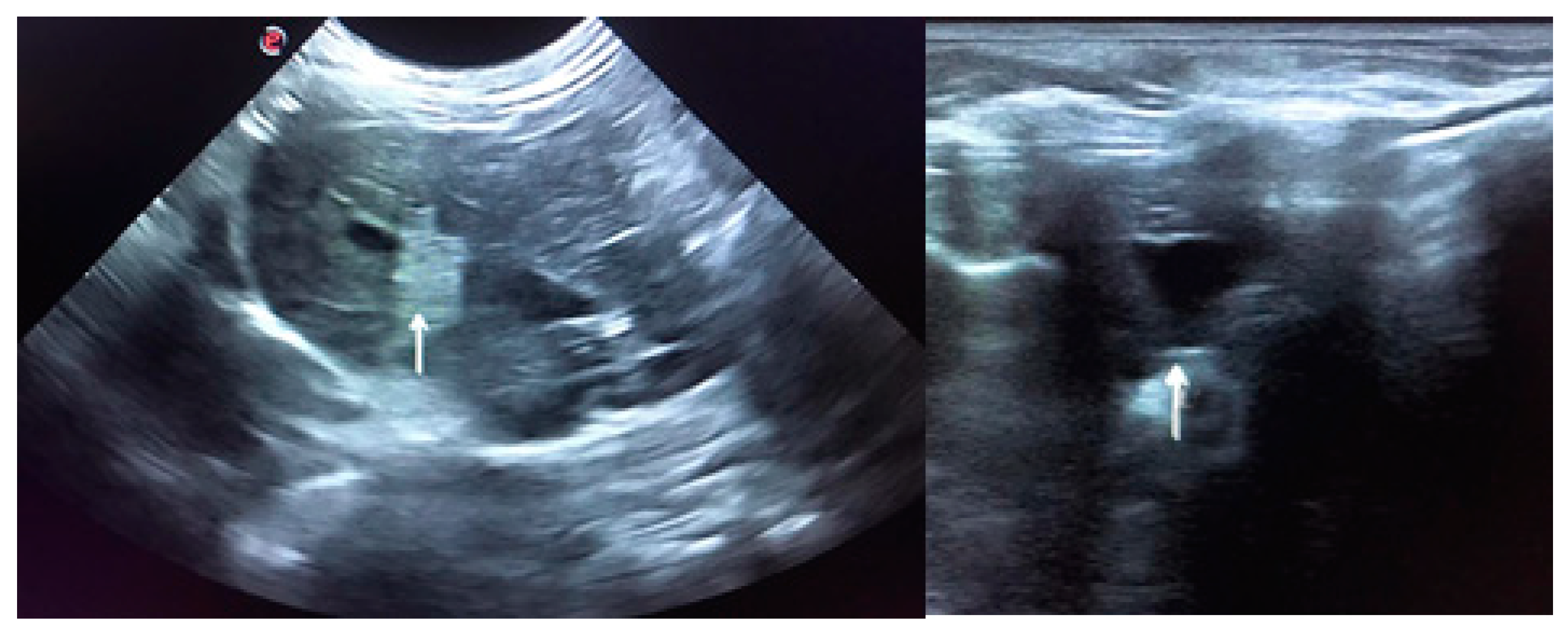
2. Clinical Presentation
3. Material and Methods
3.1. Necropsy and Histopathology
3.2. Molecular Analysis
3.3. Phylogenetic Analysis
4. Results
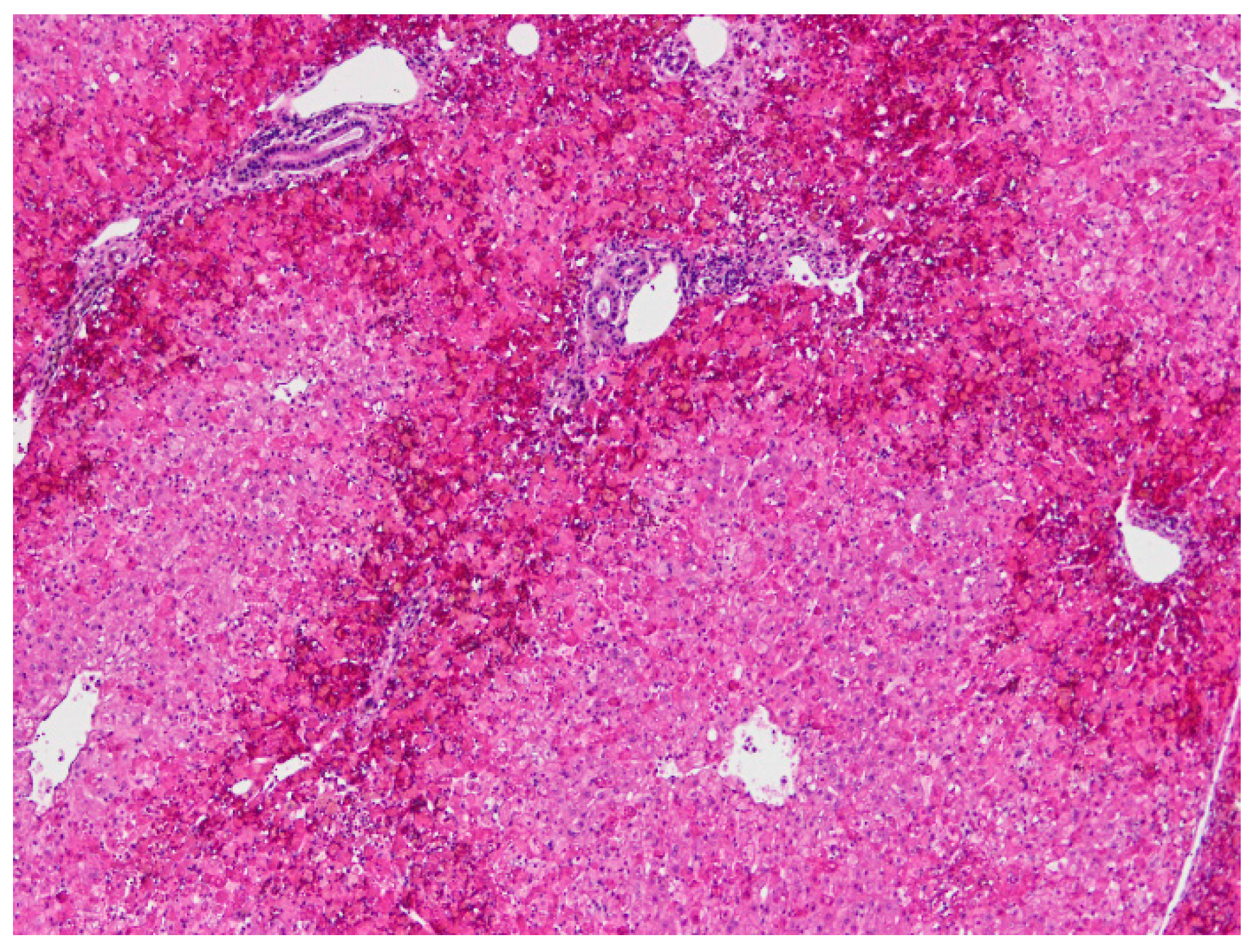
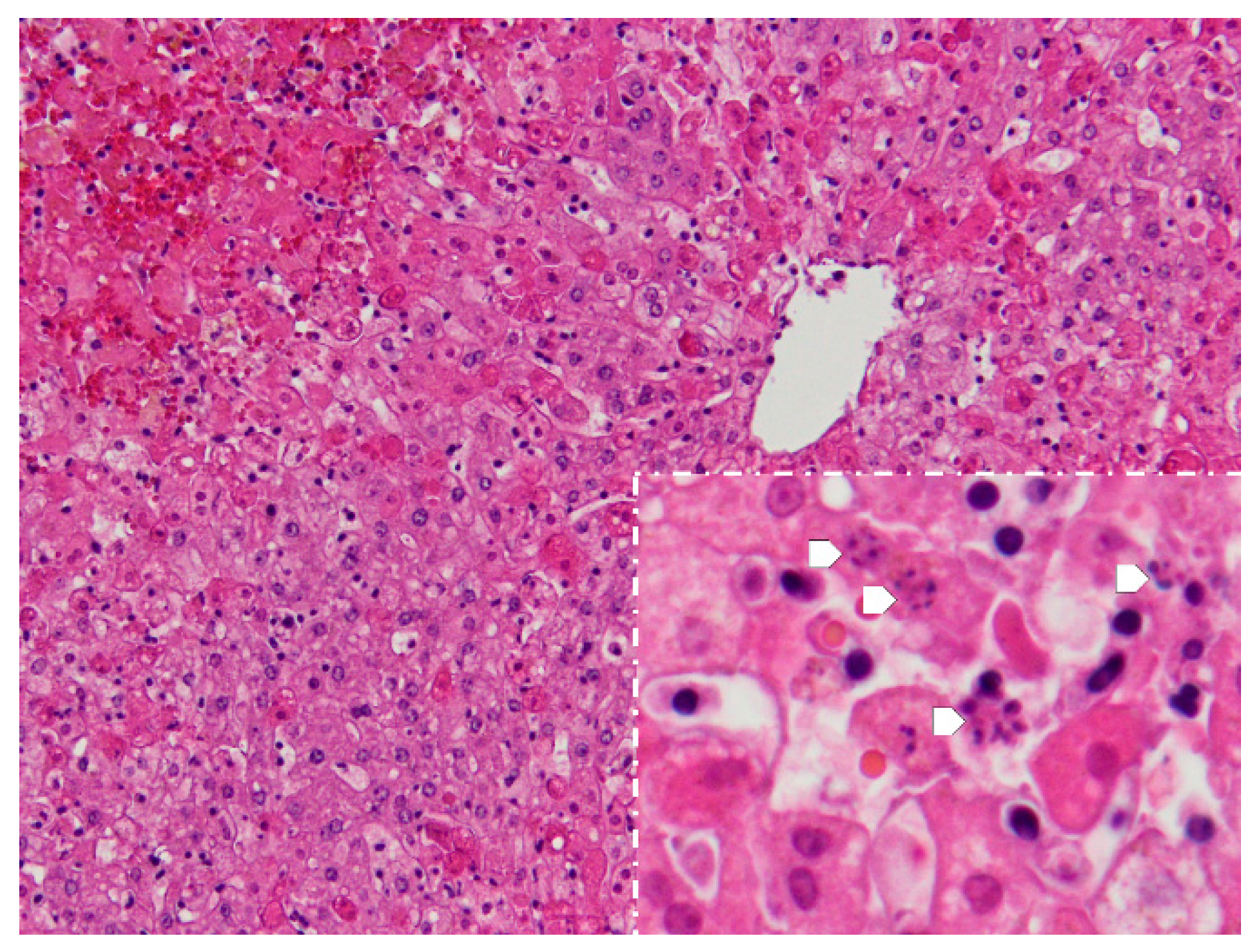
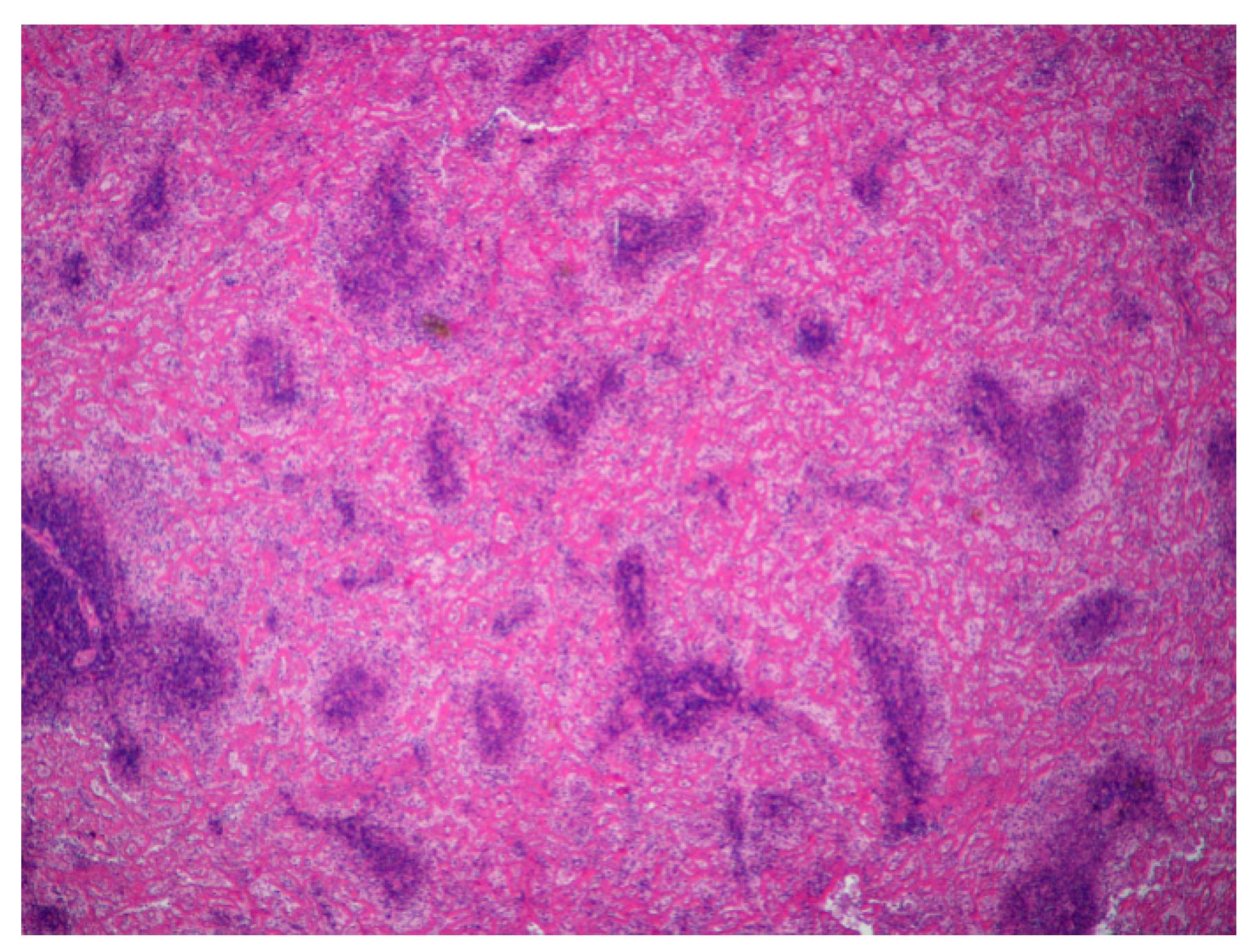
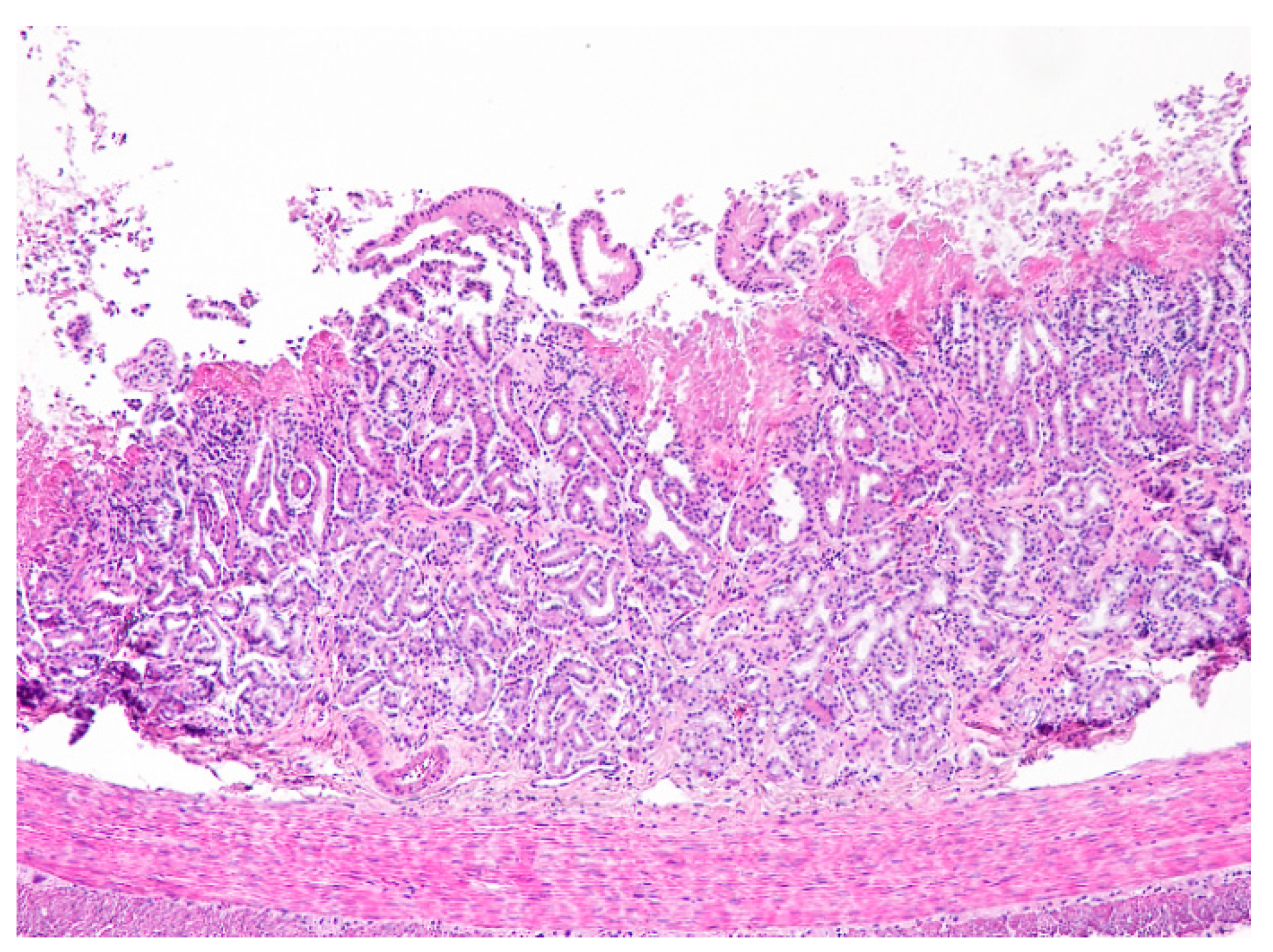
4.1. Necropsy and Histopathology
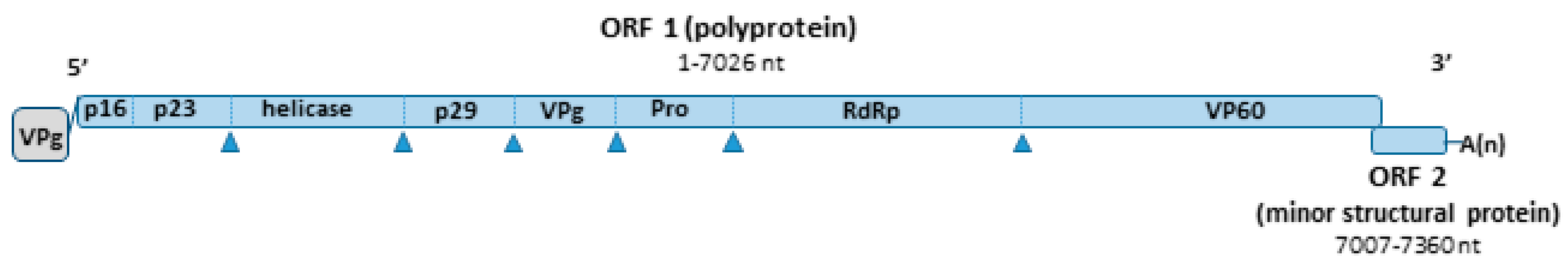
4.2. Virology
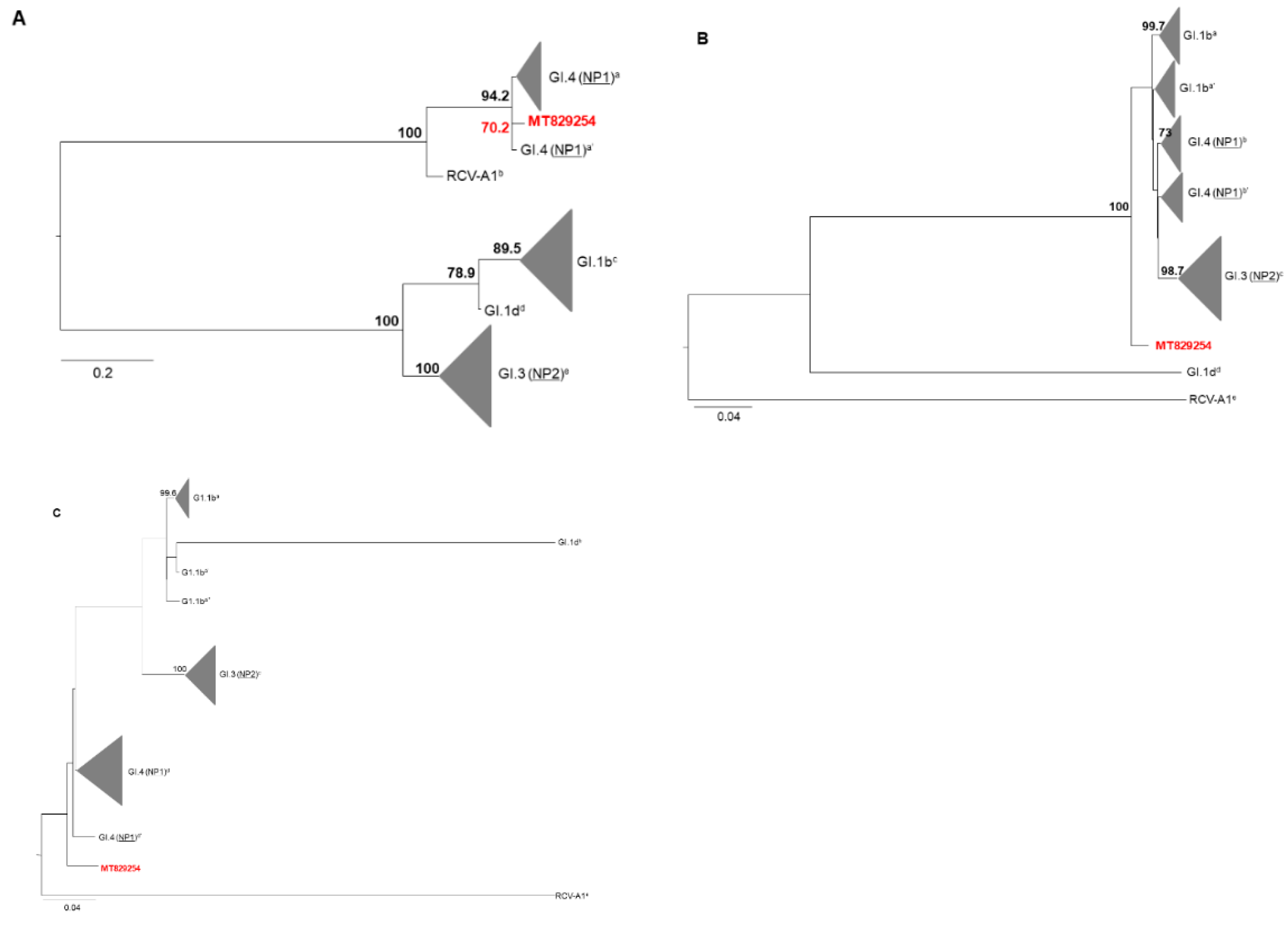
4.3. Phylogenetic Analysis
5. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.-S.; Le Gall-Reculé, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Célio, A.P.; et al. Proposal for a unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Lees, A.C.; Bell, D.J. A conservation paradox for the 21st century: The European wild rabbit Oryctolagus cuniculus, an invasive alien and an endangered native species. Mamm. Rev. 2008, 38, 304–320. [Google Scholar] [CrossRef]
- Neimanis, A.; Larsson Pettersson, U.; Huang, N.; Gavier-Widén, D.; Strive, T. Elucidation of the pathology and tissue distribution of Lagovirus europaeus GI.2/RHDV2 (rabbit haemorrhagic disease virus 2) in young and adult rabbits (Oryctolagus cuniculus). Vet. Res. 2018, 49, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE, 2019: Rabbit Haemorrhagic Disease (Technical Disease Card). 1–7. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/RHD.pdf (accessed on 3 October 2020).
- Ramiro-Ibáñez, F.; Martín-Alonso, J.M.; García Palencia, P.; Parra, F.; Alonso, C. Macrophage tropism of rabbit hemorrhagic disease virus is associated with vascular pathology. Virus Res. 1999, 60, 21–28. [Google Scholar]
- Villafuerte, R.; Calvete, C.; Blanco, J.C.; Lucientes, J. Incidence of viral hemorrhagic disease in wild rabbit populations in Spain. Mammalia 1995, 651–660. [Google Scholar] [CrossRef]
- Le Gall-Recule, G.; Zwingelstein, F.; Boucher, S.; Le Normand, B.; Plassiart, G.; Portejoie, Y.; Decors, A.; Bertagnoli, S.; Guerin, J.-L.; Marchandeau, S. Detection of a new variant of rabbit haemorrhagic disease virus in France. Vet. Rec. 2011, 168, 137–138. [Google Scholar] [CrossRef] [Green Version]
- Silvério, D.; Lopes, A.M.; Melo-ferreira, J.; Magalhaes, M.J.; Ponterroso, P.; Serronha, A.; Maio, E.; Alves, P.C.; Esteves, P.J.; Abrantes, J. Insights into the evolution of the new variant rabbit haemorrhagic disease virus (GI. 2) and the identification of novel recombinant strains. Transbound. Emerg. Dis. 2018, 65, 983–992. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guérin, J.L.; Lemaitre, E.; et al. Emergence of a new lagovirus related to rabbit haemorrhagic disease virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef] [Green Version]
- Harcourt-Brown, N.; Silkstone, M.; Whitbread, T.J.; Harcourt-Brown, F.M. RHDV2 epidemic in UK pet rabbits. Part 1: Clinical features, gross post mortem and histopathological findings. J. Small Anim. Pract. 2020, 61, 419–427. [Google Scholar] [CrossRef]
- Abade dos Santos, F.A.; Carvalho, C.; Nuno, O.; Correia, J.J.; Henriques, M.; Peleteiro, M.C.; Fevereiro, M.; Duarte, M.D. Detection of rabbit Haemorrhagic disease virus 2 during the wild rabbit (Oryctolagus cuniculus) eradication from the Berlengas archipelago, Portugal. BMC Vet. Res. 2017, 13, 336. [Google Scholar] [CrossRef] [Green Version]
- Abade dos Santos, F.A. Quadro Anatomo-Histopatológico e Diagnóstico Molecular da Doença Hemorrágica Viral em Coelho-Bravo; Faculty of Veterinary Medicine, University of Lisbon: Lisbon, Portugal, 2018. [Google Scholar]
- Barthold, W.S.; Griffey, S.M.; Percy, D.; Rabbit, H. Pathology of Laboratory Rodents and Rabbits; Wiley-Blackwell: Hoboken, NJ, USA, 2016; ISBN 9781118824245. [Google Scholar]
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Deptuła, W. The importance of liver lesions and changes to biochemical and coagulation factors in the pathogenesis of RHD. Acta Biochim. Pol. 2015, 62, 169–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plassiart, G.; Guelfi, J.-F.; Ganiere, J.-P.; Wang, B.; Andre-Fontaine, G.; Wyers, M. Hematological Parameters and Visceral Lesions Relationships in Rabbit Viral Hemorrhagic Disease. J. Vet. Med. Ser. B 1992, 39, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, Y.S.; Itakura, C. Fibrin(ogen)-related antigens in rabbits experimentally infected with rabbit haemorrhagic disease virus. Res. Vet. Sci. 1997, 63, 123–127. [Google Scholar] [CrossRef]
- Ueda, K.; Park, J.H.; Ochiai, K.; Itakura, C. Disseminated intravascular coagulation (DIC) in rabbit haemorrhagic disease. Jpn. J. Vet. Res. 1992, 40, 133–141. [Google Scholar] [PubMed]
- Rocchi, M.S.; Dagleish, M.P. Diagnosis and prevention of rabbit viral haemorrhagic disease 2. In Pract. 2018, 40, 11–16. [Google Scholar] [CrossRef]
- Soliman, M.; Rhaman, M.; Samy, M.; Mehana, O.; Nasef, S. Molecular, Clinical and Pathological Studies on Viral Rabbit Hemorrhagic Disease. Alex. J. Vet. Sci. 2016, 48, 20. [Google Scholar] [CrossRef] [Green Version]
- Duarte, M.D.; Carvalho, C.L.; Abade dos Santos, F.A.; Gomes, J.; Alves, P.C.; Esteves, P.J.; Abrantes, J.; Lopes, A.M.; Lopes, P.M.; Serronha, A.; et al. +Coelho: Avaliação Ecossanitária das Populações Naturais de Coelho Bravo Visando o Controlo da Doença Hemorrágica Viral. Master’s Thesis, University of Lisbon, Lisboa, Portugal, 2018. [Google Scholar]
- Montbrau, C.; Padrell, M.; Ruiz, M.C. Efficacy and Safety of a New Inactivated Vaccine against the Rabbit Haemorrhagic Disease Virus 2-Like Variant (RHDV-2). 2012, Volume 1984. Available online: https://www.hipra.com/wcm/connect/hipra/d0be8fc3-788a-4910-a47e-64709166ad1c/Efficacy+and+safety+of+a+new+inactivated+vaccine+against+RHDV-2.pdf?MOD=AJPERES&CACHEID=ROOTWORKSPACE.Z18_GG50HI40O8ABD0Q8OC940F2000-d0be8fc3-788a-4910-a47e-64709166ad1c-lAmNnR1 (accessed on 25 September 2020).
- Office International des Epizooties. OIE Rabbit Haemorrhagic Disease. In OIE Terrestrial Manual 2018; Office International des Epizooties: Paris, France, 2018; pp. 1389–1406. Available online: www.oie.int (accessed on 15 September 2020).
- Fisher, P.; Graham, J. Chapter 10—Rabbits. In Exotic Animal Formulary, 5th ed.; Carpenter, J.W., Marion, C.J., Eds.; Elsevier Saunders: St. Louis, MI, USA, 2018; pp. 494–531. ISBN 978-0-323-44450-7. [Google Scholar]
- Tham, K.M.; Barnes, S.M.; Hunter, S.N. Polymerase chain reaction amplification and gene sequence analysis of a calicivirus from a feral rabbit. Virus Genes 1999, 18, 235–242. [Google Scholar] [CrossRef]
- Duarte, M.D.; Carvalho, C.L.; Barros, S.C.; Henriques, A.M.; Ramos, F.; Fagulha, T.; Luís, T.; Duarte, E.L.; Fevereiro, M. A real time Taqman RT-PCR for the detection of rabbit hemorrhagic disease virus 2 (RHDV2). J. Virol. Methods 2015, 219, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Duarte, M.D.; Barros, S.C.; Henriques, A.M.; Fagulha, M.T.; Ramos, F.; Luís, T.; Fevereiro, M. Development and validation of a real time PCR for the detection of myxoma virus based on the diploid gene M000. 5L/R. J. Virol. Methods 2014, 196, 219–224. [Google Scholar] [CrossRef]
- R Development Core Team R. A Language and Environment for Statistical Computing; R Found. Stat. Comput.: Vienna, Austria, 2008; ISBN 3-900051-07-0. [Google Scholar]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Tavaré, S.; Miura, R.M. Lectures on mathematics in the life sciences. Am. Math. Soc. 1986, 57–86. [Google Scholar]
- Carvalho, C.L.; Duarte, E.L.; Monteiro, M.; Botelho, A.; Albuquerque, T.; Fevereiro, M.; Henriques, A.M.; Barros, S.S.; Duarte, M.D. Challenges in the rabbit haemorrhagic disease 2 (RHDV2) molecular diagnosis of vaccinated rabbits. Vet. Microbiol. 2017, 198, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrantes, J.; Droillard, C.; Lopes, A.M.; Lemaitre, E.; Lucas, P.; Blanchard, Y.; Marchandeau, S.; Esteves, P.J.; Le Gall-Reculé, G. Recombination at the emergence of the pathogenic rabbit haemorrhagic disease virus Lagovirus europaeus/GI.2. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Blanco-Aguiar, J.; Martín-Alonso, A.; Leitão, M.; Foronda, P.; Mendes, M.; Gonçalves, D.; Abrantes, J.; Esteves, P.J. Full genome sequences are key to disclose RHDV2 emergence in the Macaronesian islands. Virus Genes 2018, 54, 1–4. [Google Scholar] [CrossRef]
- Dalton, K.P.; Arnal, J.L.; Benito, A.A.; Chacón, G.; Martín Alonso, J.M.; Parra, F. Conventional and real time RT-PCR assays for the detection and differentiation of variant rabbit hemorrhagic disease virus (RHDVb) and its recombinants. J. Virol. Methods 2018, 251, 118–122. [Google Scholar] [CrossRef]
- Bonvehí, C.; Ardiaca, M.; Montesinos, A.; Juan, C.; Gómez, A.; Teso, B.; Barbero, S.; Ekei, S. Clinicopathologic findings of naturally occurring Rabbit Hemorrhagic Disease Virus 2 infection in pet rabbits. Vet. Clin. Pathol. 2019, 48, 89–95. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haematology | February | March | June | Reference Values | |
|---|---|---|---|---|---|
| Carpenter, 2018 | Idexx Lab., 2017 | ||||
| Haematocrit (%) | 30.9 | 36.3 | 37.6 | 30–50 | 29.4–40.9 |
| Haemoglobin (g/dL) | 10.7 | 11.8 | 13.3 | 8–17.5 | 9.8–13.2 |
| Erythrocytes (×106/μL) | 5.21 | 5.67 | 6.45 | 4–8 | 4.45–6.71 |
| MCV (fL) | 59.3 | 64.0 | 58.3 | 58–75 | 58.1–69.6 |
| MCH (pg) | 20.5 | 20.8 | 20.6 | 17.5–23.5 | 18.9–22.1 |
| MCHC (g/dL) | 34.6 | 32.5 | 35.4 | 29–37 | 31.6–33.6 |
| Reticulocytes (%) | 2.4 | 5.4 | 1.4 | 2–4 | - |
| Reticulocytes (K/μL) | 122.4 | 306.7↑ | 88.4 | - | 69.5–242.7 |
| Platelets (103/μL) | 40↓ | 370 | 97↓ | 290–650 | 219–521 |
| WBC (103/μL) | 2.53↓ | 5.15 | 1.86↓ | 5–12 | 4.54–10.22 |
| Heterophils (%) | 28.1↓ | 28.8↓ | 47.9 | 35–55 | - |
| Lymphocytes (%) | 64.4↑ | 60.0 | 41.9 | 25–60 | - |
| Monocytes (%) | 4.7 | 5.4 | 4.8 | 2–10 | - |
| Eosinophils (%) | 0.8 | 0.8 | 0.0 | 0–5 | - |
| Basophils (%) | 2.0 | 5.0 | 5.4 | 2–8 | - |
| Heterophils (K/μL) | 0.71↓ | 1.48 | 0.89↓ | - | 0.96–3.34 |
| Lymphocytes (K/μL) | 1.63 | 3.09 | 0.78↓ | - | 1.49–5.21 |
| Monocytes (K/μL) | 0.12↓ | 0.28↓ | 0.09↓ | - | 0.31–0.99 |
| Eosinophils (K/μL) | 0.02↓ | 0.04↓ | 0.00↓ | - | 0.05–2.12 |
| Basophils (K/μL) | 0.05↓ | 0.26↓ | 0.10↓ | - | 0.56–2.12 |
| Biochemistry | |||||
| Glucose | 194↑ | - | 164↑ | 75–150 | 75–145 |
| Creatinine (CREA) | 1.0 | - | 1.2 | 0.5–2.6 | 0.8–1.8 |
| BUN | 12 | - | 22 | 15–50 | 10–24 |
| BUN/CREA | 12 | - | 18 | - | - |
| PHOS | - | - | 3.7 | 2.3–6.9 | 1.2–4.9 |
| CA | - | - | 11.7 | 8–14.8 | 5.6–12.0 |
| TP | 6.6 | - | 7.7↑ | 5.4–7.5 | 5.5–7.2 |
| ALB | 4.4 | - | 4.4 | 2.5–5 | 2.7–4.6 |
| GLOB | 2.2 | - | 3.3↑ | 1.5–3.5 | 1.5–2.8 |
| ALB/GLOB | 2.0 | - | 1.3 | - | - |
| ALT | 240↑ | - | 586↑ | 14–80 | 31–53 |
| ALKP | 99 | - | 104 | 4–70 | 70–145 |
| GGT | - | - | 2 | - | - |
| TBIL | - | - | 0.9 | 0–0.75 | 0.3–0.8 |
| CHOL | - | - | 30 | 12–16 | 35–53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abade dos Santos, F.A.; Magro, C.; Carvalho, C.L.; Ruivo, P.; Duarte, M.D.; Peleteiro, M.C. A Potential Atypical Case of Rabbit Haemorrhagic Disease in a Dwarf Rabbit. Animals 2021, 11, 40. https://doi.org/10.3390/ani11010040
Abade dos Santos FA, Magro C, Carvalho CL, Ruivo P, Duarte MD, Peleteiro MC. A Potential Atypical Case of Rabbit Haemorrhagic Disease in a Dwarf Rabbit. Animals. 2021; 11(1):40. https://doi.org/10.3390/ani11010040
Chicago/Turabian StyleAbade dos Santos, Fábio A., Carolina Magro, Carina L. Carvalho, Pedro Ruivo, Margarida D. Duarte, and Maria C. Peleteiro. 2021. "A Potential Atypical Case of Rabbit Haemorrhagic Disease in a Dwarf Rabbit" Animals 11, no. 1: 40. https://doi.org/10.3390/ani11010040
APA StyleAbade dos Santos, F. A., Magro, C., Carvalho, C. L., Ruivo, P., Duarte, M. D., & Peleteiro, M. C. (2021). A Potential Atypical Case of Rabbit Haemorrhagic Disease in a Dwarf Rabbit. Animals, 11(1), 40. https://doi.org/10.3390/ani11010040