
Prandial Correlations and Structure of the Ingestive Behavior of Pigs in Precision Feeding Programs


 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source
2.2. Diets and Feeding
2.3. Data Collection and Management
2.4. Definition of the Meal Criterion
2.5. Statistical Analysis
3. Results
3.1. General Description and Meal Criterion
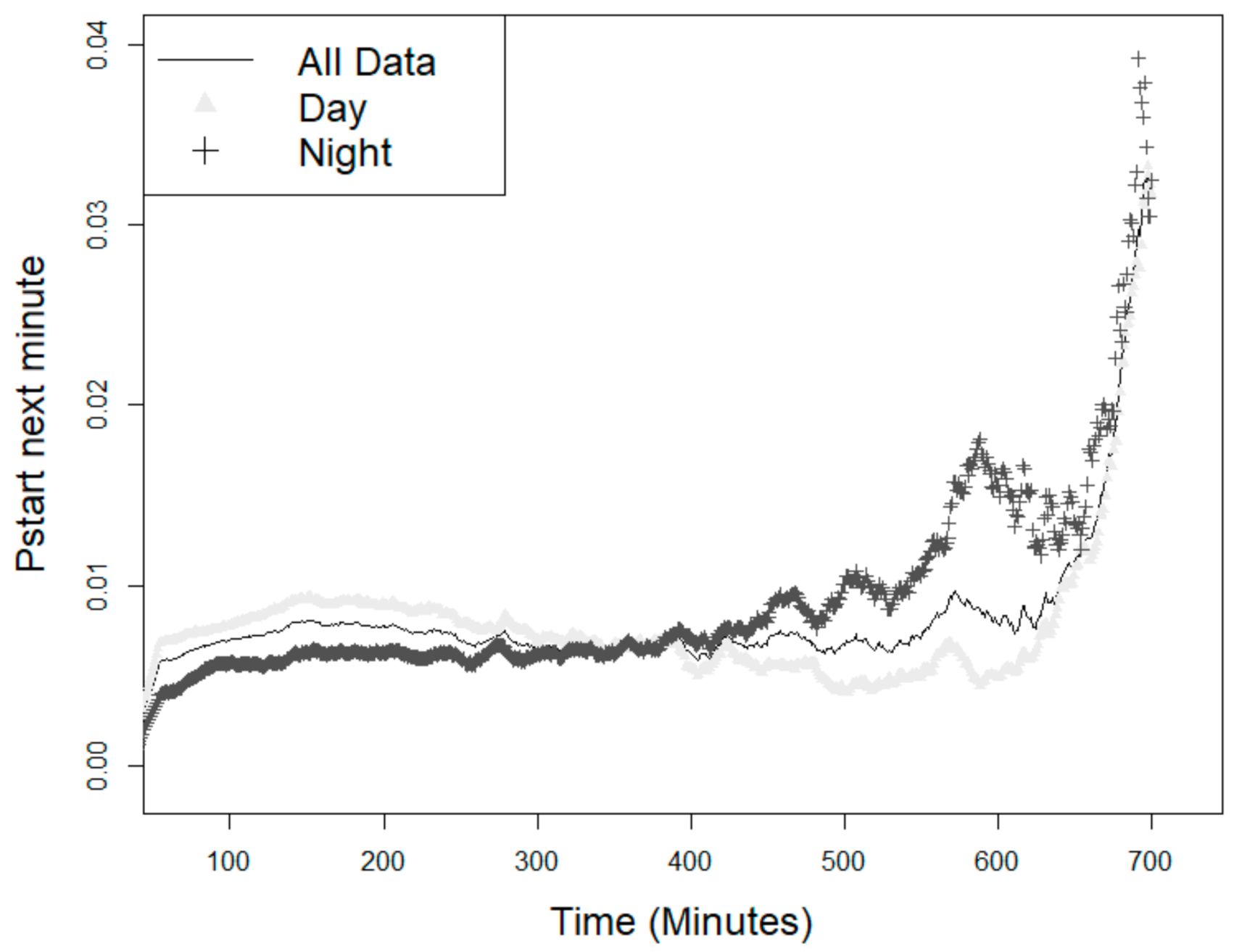
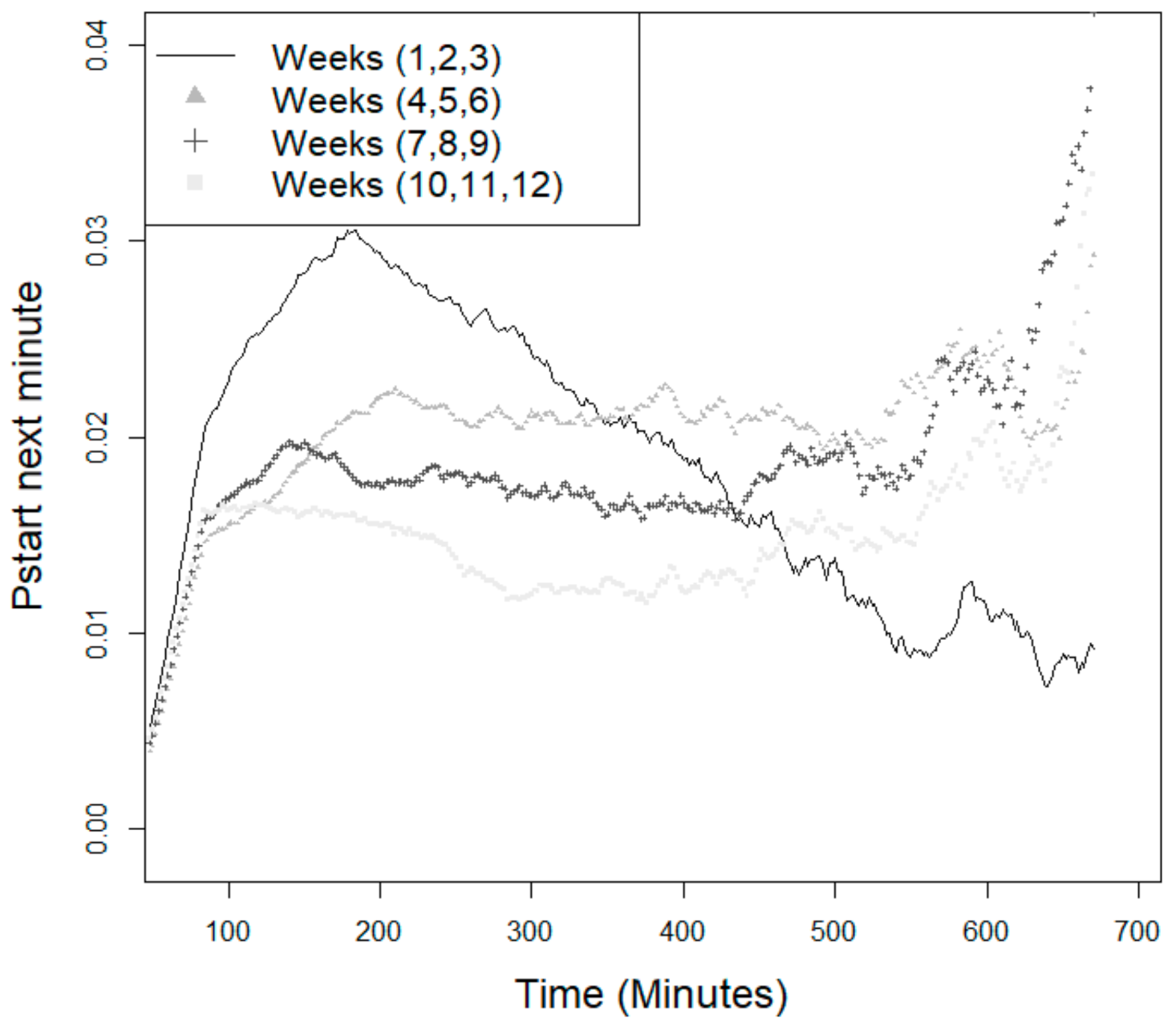
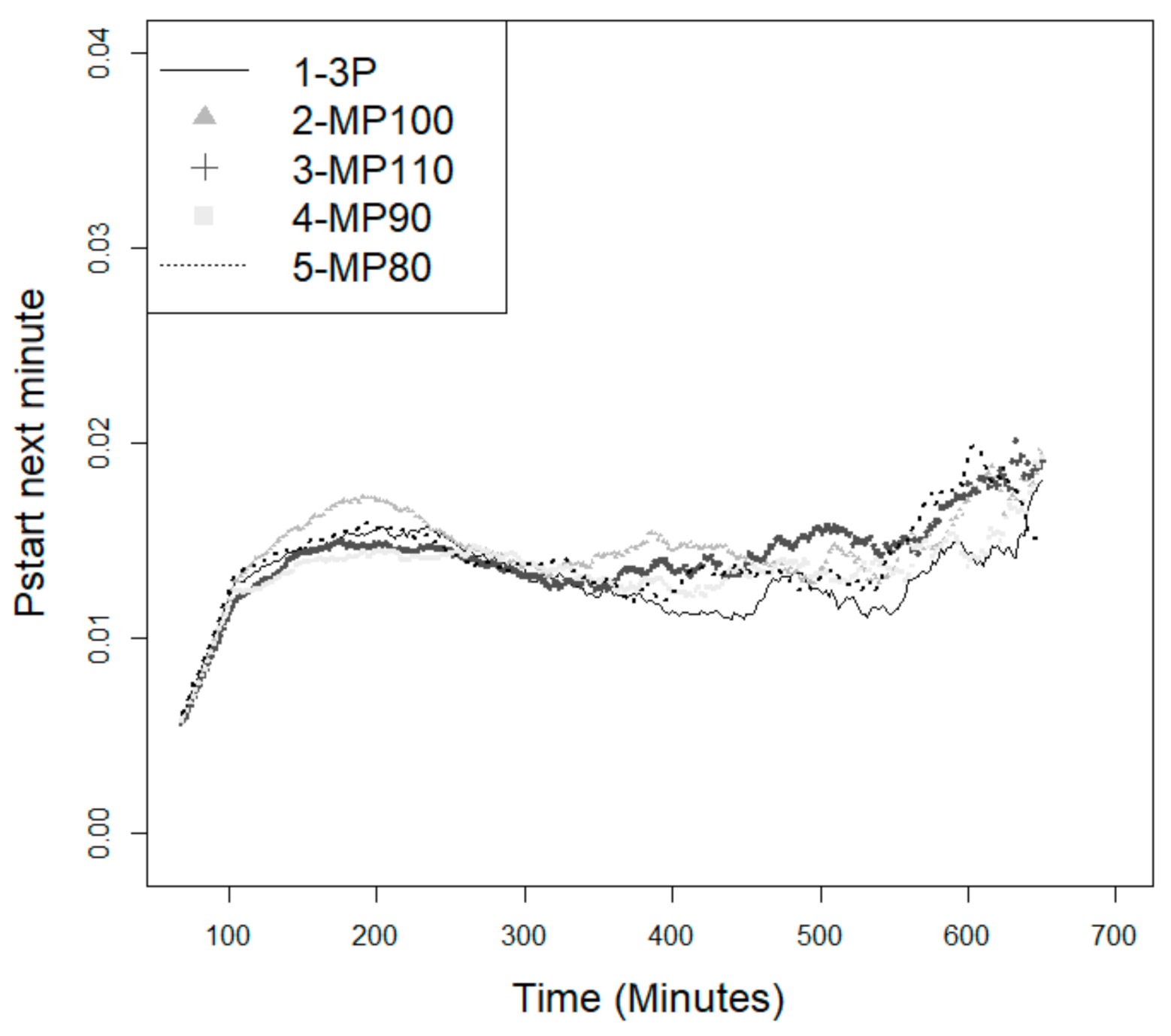
3.2. Probability of the Animal Starting a New Feeding Event
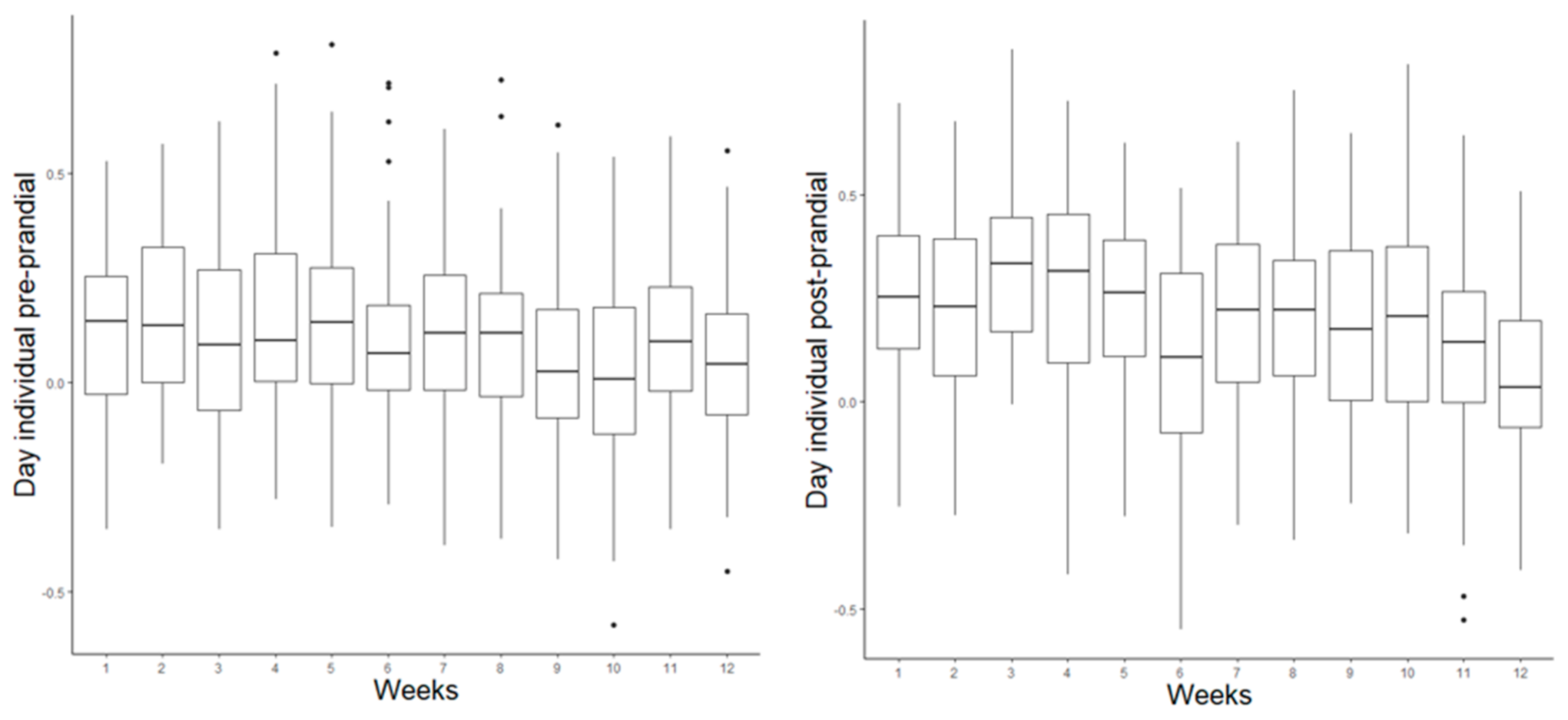
3.3. Prandial Correlations
4. Discussion
4.1. Meal Criterion
4.2. Description of Feeding Behavior
4.3. Probability of Starting a New Feeding Event—The Circadian Rhythm
4.4. Probability of Starting a New Feeding Event—Effect of Age
4.5. Probability of Starting a New Feeding Event—Effect of the Feeding Programs
4.6. Prandial Correlations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rayer, H.; Shirali, M.; Siriluck, P.; Murani, E.; Valey, P.F.; Jensen, J.; Wimmers, K. Exploring the genetics of feed efficiency and feeding behaviour traits in a pig line highly selected for performance characteristics. Mol. Genet. Genom. 2017, 292, 1001–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, C.A.; Emmans, G.C.; Tolkamp, B.J.; Kyriazakis, I. Analysis of the feeding behavior of pigs using different models. Physiol. Behav. 2000, 68, 395–403. [Google Scholar] [CrossRef]
- Andretta, I.; Pomar, C.; Rivest, J.; Pomar, J.; Radunz, J. Precision feeding can significantly reduce lysine intake and nitrogen excretion without compromising the performance of growing pigs. Animal 2016, 10, 1137–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maselyne, J.; Saeysa, W.; Van Nuffel, A. Review: Quantifying animal feeding behaviour with a focus on pigs. Physiol. Behav. 2015, 138, 37–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolkamp, B.; Howie, A.J.; Bleyb, A.G.T.; Kyriazakisc, L. Prandial correlations and the structure of feeding behavior. Appl. Anim. Behav. Sci. 2012, 137, 53–65. [Google Scholar] [CrossRef]
- Savory, C.J. Correlations between meals and inter-meal intervals in Japanese quail and their significance in the control of feeding. Behav. Process. 1981, 6, 23–36. [Google Scholar] [CrossRef]
- Collier, G.; Johnson, D.F.; Mitchell, C. The relation between meal size and the time between meals: Effects of cage complexity and food cost. Physiol. Behav. 1999, 67, 339–346. [Google Scholar] [CrossRef]
- Tolkamp, B.J.; Schweitzer, D.P.N.; Kyriazakis, I. The biologically relevant unit for the analysis of short-term feeding behavior of dairy cows. J. Dairy Sci. 2000, 83, 2057–2068. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine: Eleventh Revised Edition; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Jondreville, C.; Dourmad, J.Y. Le phosphore dans la nutrition des porcs. Inra Prod. Anim. 2005, 18, 183–192. [Google Scholar] [CrossRef]
- Hauschild, L.; Lovatto, P.A.; Pomar, J.; Pomar, C. Development of sustainable precision farming systems for swine: Estimating real-time individual amino acid requirements in growing-finishing pigs. J. Anim. Sci. 2012, 90, 2255–2263. [Google Scholar] [CrossRef]
- Hauschild, L.; Pomar, C.; Lovatto, P.A. Systematic comparison of the empirical and factorial methods used to estimate the nutrient requirements of growing pigs. Animal 2010, 4, 714–723. [Google Scholar] [CrossRef]
- Bigelow, J.A.; Houpt, T.R. Feeding and drinking patterns in young pigs. Physiol. Behav. 1988, 43, 99–109. [Google Scholar] [CrossRef]
- Howie, J.A.; Tolkamp, B.J.; Avendaño, S.; Kyriazakis, I. A novel flexible method to split feeding behaviour into bouts. Appl. Anim. Behav. Sci. 2009, 116, 101–109. [Google Scholar] [CrossRef]
- Tolkamp, B.J.; Allcroft, D.J.; Barrio, J.P.; Bley, T.A.G.; Howie, J.A.; Jacobsen, T.B.; Morgan, C.A.; Schweitzer, D.P.N.; Wilkinson, S.; Yeates, M.P.; et al. The temporal structure of feeding behavior. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, 378–393. [Google Scholar] [CrossRef] [Green Version]
- Geary, N. A new way of looking at eating. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, 1444–1446. [Google Scholar] [CrossRef]
- Zorrilla, E.P.; Inoue, K.; Fekete, E.M.; Tabarin, A.; Valdez, G.R.; Koob, G.F. Measuring meals: Structure of prandial food and water intake of rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, 1450–1467. [Google Scholar] [CrossRef] [Green Version]
- Drouilhet, L.; Monteville, R.; Molette, C.; Lague, M.; Cornuez, A.; Canario, L.; Ricard, E.; Gilbert, H. Impact of selection for residual feed intake on production traits and behavior of mule ducks. Poult. Sci. 2016, 95, 1999–2010. [Google Scholar] [CrossRef]
- Slater, P.J.B.; Lester, N.P. Minimising Errors in Splitting Behaviour into Bouts. Behaviour 1982, 79, 153–161. [Google Scholar] [CrossRef]
- Forbes, J.M. Food preferences in farm animals: Why don’t they always choose wisely? Proc. Nutr. Soc. 1997, 54, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Sparkes, G.M.; Cole, D.J.A.; Lewis, D. 1981. The effect of dietary-protein level on voluntary feed-intake in the pig. Anim. Prod. 1981, 32, 356–357. [Google Scholar]
- Bokkers, E.A.M.; Koene, P. Behaviour of fast- and slow growing broilers to 12 weeks of age and the physical consequences. Appl. Anim. Behav. Sci. 2003, 81, 59–72. [Google Scholar] [CrossRef]
- Young, R.J.; Lawrence, A.B. Feeding behaviour of pigs in groups monitored by a computerized feeding system. Anim. Sci. 1994, 58, 145–152. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response | First Quartile | Median | Mean | Third Quartile |
|---|---|---|---|---|
| Interval between visits, minutes | 0.05 | 0.25 | 43.6 | 25.5 |
| Time duration per visits, minutes | 0.61 | 1.11 | 2.03 | 2.31 |
| Intake per visits, g | 21.0 | 44.0 | 89.5 | 104.8 |
| Interval between meals, minutes | 83.2 | 141.5 | 198.3 | 242.4 |
| Meal size, minutes | 5.05 | 9.33 | 12.95 | 15.65 |
| Intake per meal, g | 153.8 | 305.2 | 358.9 | 504.8 |
| Weeks | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| Pre-prandial correlations | ||||||||||||
| 24 h | 0.25 * | 0.21 * | 0.18 * | 0.18 * | 0.13 * | 0.11 * | 0.10 * | 0.13 * | 0.07 * | 0.02 † | 0.02 * | –0.02 † |
| Day | 0.18 * | 0.19 * | 0.14 * | 0.13 * | 0.09 * | 0.07 * | 0.09 * | 0.09 * | 0.06 * | 0.06 † | 0.02 * | 0.03 * |
| Night | 0.26 * | 0.25 * | 0.33 * | 0.28 * | 0.24 * | 0.21 * | 0.14 * | 0.22 * | 0.16 * | 0.13 † | 0.14 * | 0.15 * |
| Post-prandial correlation | ||||||||||||
| 24 h | 0.26 * | 0.23 * | 0.27 * | 0.24 * | 0.21 * | 0.12 * | 0.20 * | 0.25 * | 0.23 * | 0.23 * | 0.16 * | 0.14 * |
| Day | 0.32 * | 0.28 * | 0.35 * | 0.30 * | 0.24 * | 0.13 * | 0.22 * | 0.27 * | 0.22 * | 0.24 * | 0.17 * | 0.14 * |
| Night | 0.19 * | 0.24 * | 0.24 * | 0.22 * | 0.23 * | 0.11 * | 0.19 * | 0.19 * | 0.24 * | 0.18 * | 0.14 † | 0.13 † |
| Feeding Program 1,2 | |||||
|---|---|---|---|---|---|
| 3P | MP110 | MP100 | MP90 | MP80 | |
| 24 h pre-prandial | 0.14 | 0.11 | 0.13 | 0.14 | 0.13 |
| 24 h post-prandial | 0.26 | 0.23 | 0.23 | 0.20 | 0.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, B.C.K.; Andretta, I.; Valk, M.; Pomar, C.; Hauschild, L.; Fraga, A.Z.; Kipper, M.; Trevizan, L.; Remus, A. Prandial Correlations and Structure of the Ingestive Behavior of Pigs in Precision Feeding Programs. Animals 2021, 11, 2998. https://doi.org/10.3390/ani11102998
Gomes BCK, Andretta I, Valk M, Pomar C, Hauschild L, Fraga AZ, Kipper M, Trevizan L, Remus A. Prandial Correlations and Structure of the Ingestive Behavior of Pigs in Precision Feeding Programs. Animals. 2021; 11(10):2998. https://doi.org/10.3390/ani11102998
Chicago/Turabian StyleGomes, Bruna C. K., Ines Andretta, Marcio Valk, Candido Pomar, Luciano Hauschild, Alícia Z. Fraga, Marcos Kipper, Luciano Trevizan, and Aline Remus. 2021. "Prandial Correlations and Structure of the Ingestive Behavior of Pigs in Precision Feeding Programs" Animals 11, no. 10: 2998. https://doi.org/10.3390/ani11102998
APA StyleGomes, B. C. K., Andretta, I., Valk, M., Pomar, C., Hauschild, L., Fraga, A. Z., Kipper, M., Trevizan, L., & Remus, A. (2021). Prandial Correlations and Structure of the Ingestive Behavior of Pigs in Precision Feeding Programs. Animals, 11(10), 2998. https://doi.org/10.3390/ani11102998