
LC-MS/MS Based Metabolomics Reveal Candidate Biomarkers and Metabolic Changes in Different Buffalo Species


 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Milk Composition Determination and Metabolite Extraction
2.3. LC-MS/MS Analysis
2.4. Data Analysis
2.5. Metabolic Pathway Analysis
3. Results
3.1. Routine Analysis of Milk Composition
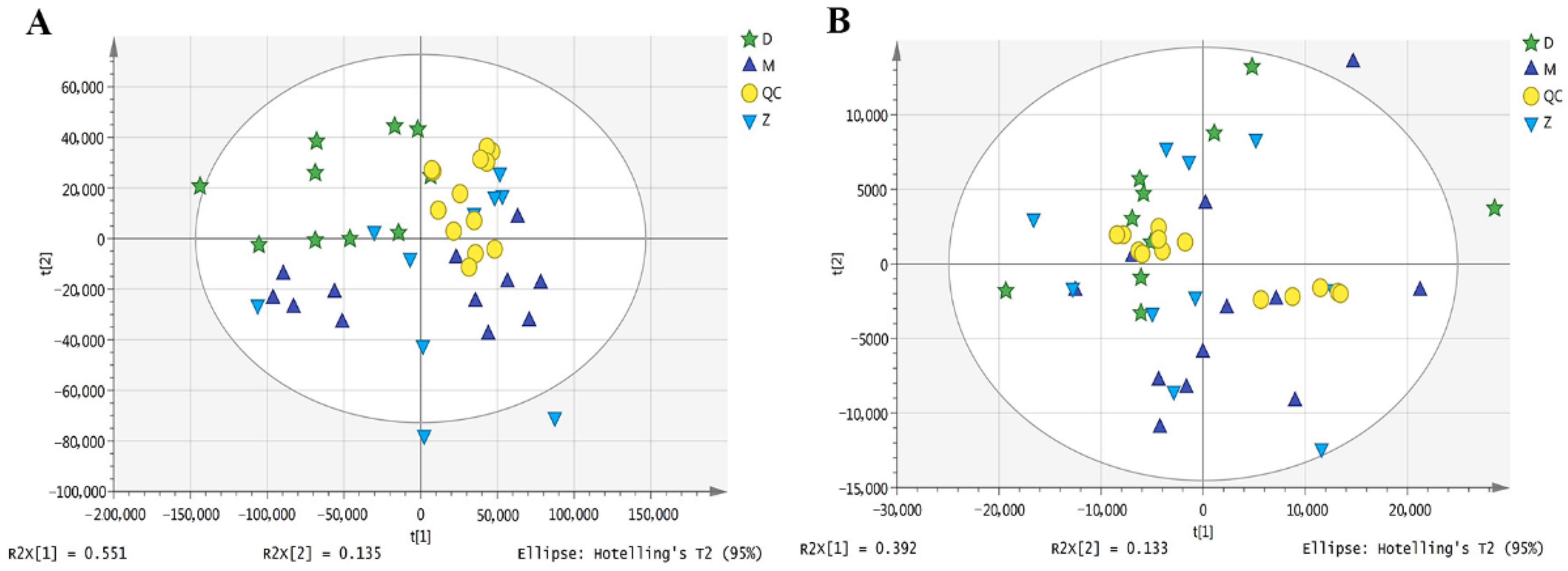
3.2. Milk Metabolome Profiles
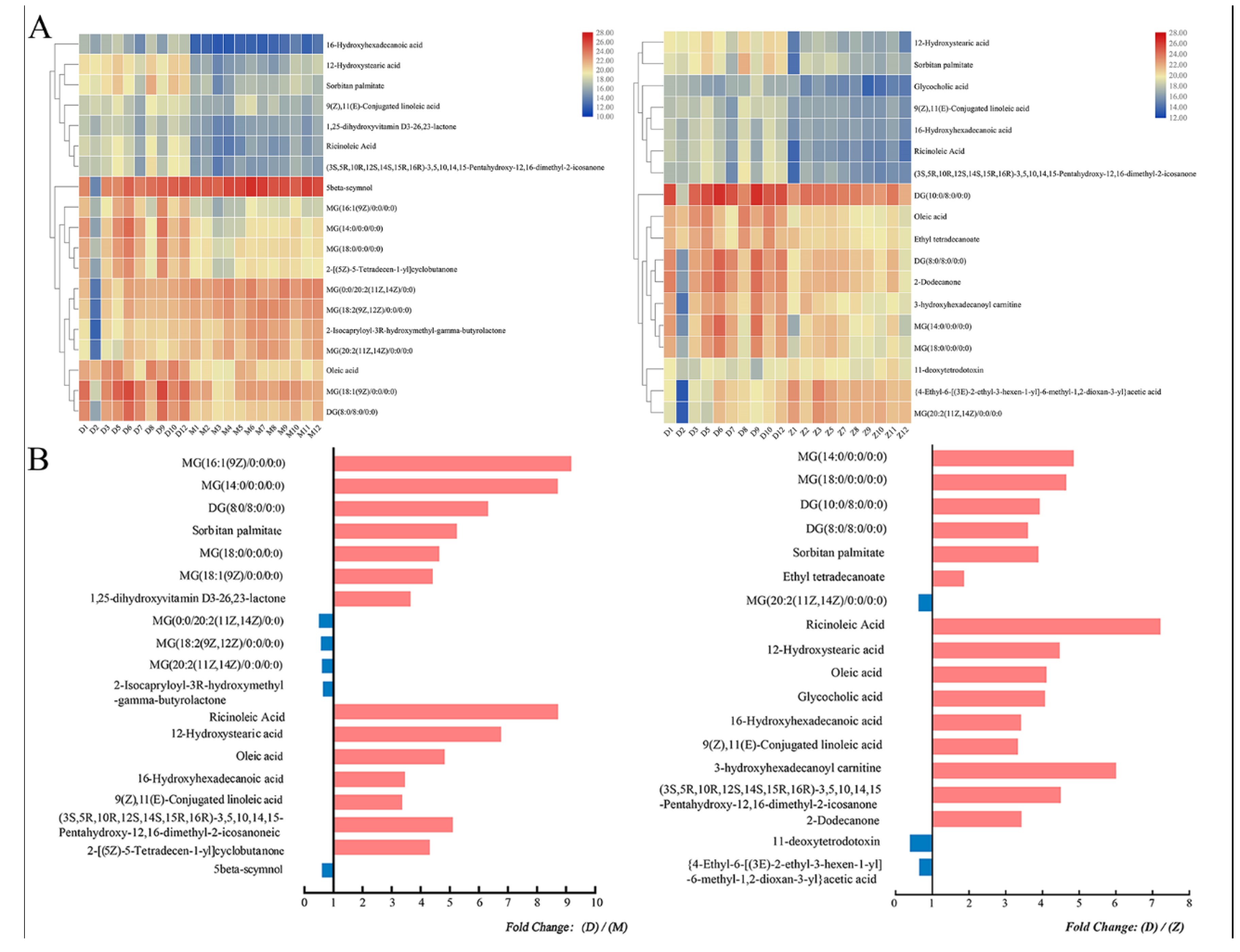
3.3. Identification of Differential Metabolites
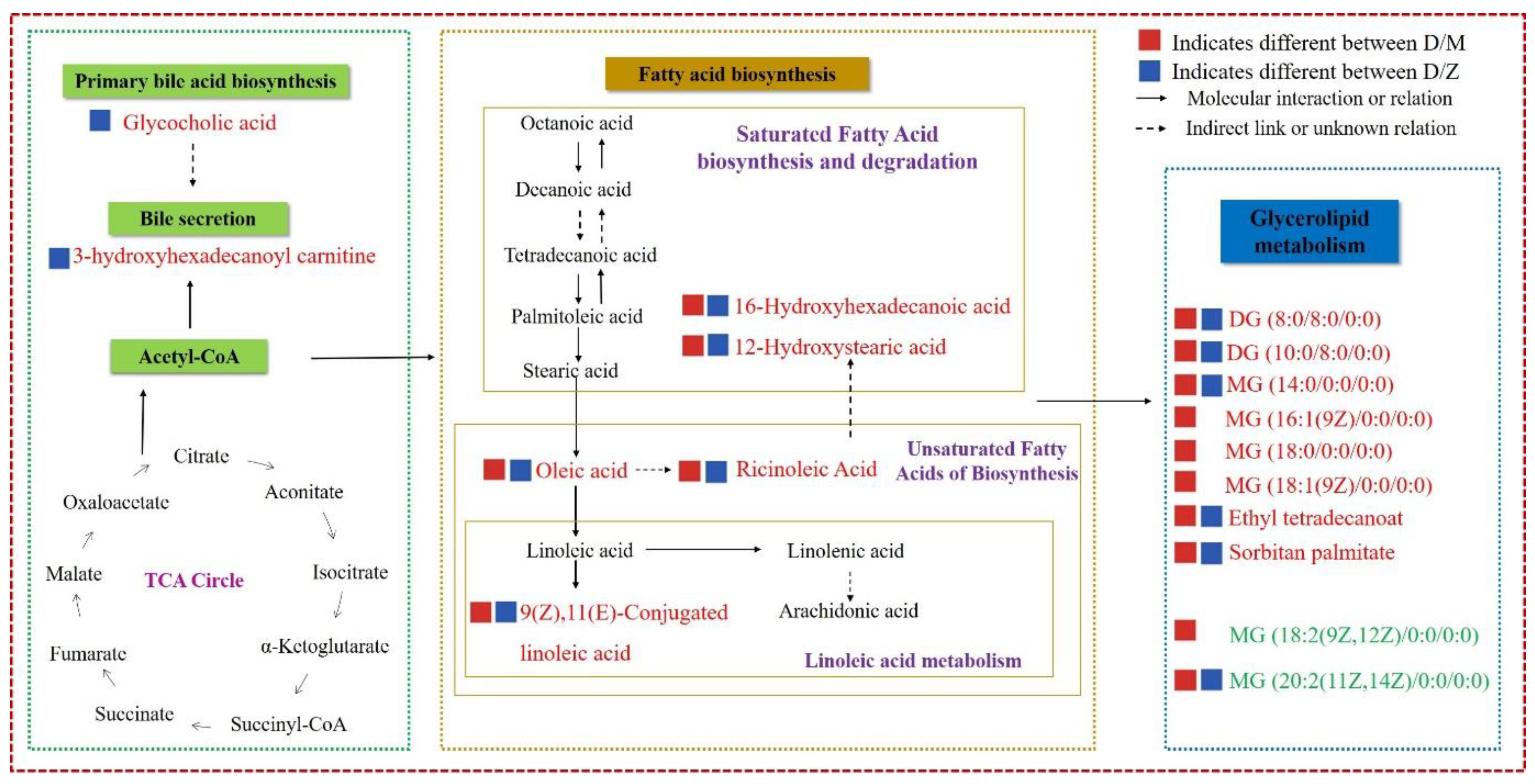
3.4. Integration of Key Different Metabolic Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Tomassini, A.; Curone, G.; Solè, M.; Capuani, G.; Sciubba, F.; Conta, G.; Miccheli, A.; Vigo, D. NMR-based metab-olomics to evaluate the milk composition from Friesian and autochthonous cows of Northern Italy at different lactation times. Nat. Prod. Res. 2019, 33, 1085–1091. [Google Scholar] [CrossRef]
- Wang, J.; He, Y.; Pang, K.; Zeng, Q.; Zhang, X.; Ren, F.; Guo, H. Changes in milk yield and composition of colos-trum and regular milk from four buffalo breeds in China during lactation. J. Sci. Food Agric. 2019, 99, 5799–5807. [Google Scholar] [CrossRef]
- Han, B.-Z.; Meng, Y.; Li, M.; Yang, Y.-X.; Ren, F.-Z.; Zeng, Q.-K.; Nout, M.R. A survey on the microbiological and chemical composition of buffalo milk in China. Food Control 2007, 18, 742–746. [Google Scholar] [CrossRef]
- Zhou, L.; Tang, Q.; Iqbal, M.W.; Xia, Z.; Huang, F.; Li, L.; Liang, M.; Lin, B.; Qin, G.; Zou, C. A comparison of milk protein, fat, lactose, total solids and amino acid profiles of three different buffalo breeds in Guangxi, China. Ital. J. Anim. Sci. 2018, 17, 873–878. [Google Scholar] [CrossRef] [Green Version]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jiang, M. Metabolomic profiles in yak mammary gland tissue during the lactation cycle. PLoS ONE 2019, 14, e0219220. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, N.; Zhao, X.; Zhang, Y.; Han, R.; Yang, J.; Zhao, S.; Li, S.; Guo, T.; Zang, C. Metabolomic bi-omarkers identify differences in milk produced by Holstein cows and other minor dairy animals. J. Proteom. 2016, 136, 174–182. [Google Scholar] [CrossRef]
- Li, S.; Wang, Q.; Lin, X.; Jin, X.; Liu, L.; Wang, C.; Chen, Q.; Liu, J.; Liu, H. The use of “Omics” in lactation research in dairy cows. Int. J. Mol. Sci. 2017, 18, 983. [Google Scholar] [CrossRef] [Green Version]
- Ceciliani, F.; Lecchi, C.; Urh, C.; Sauerwein, H. Proteomics and metabolomics characterizing the pathophysiology of adaptive reactions to the metabolic challenges during the transition from late pregnancy to early lactation in dairy cows. J. Proteom. 2018, 178, 92–106. [Google Scholar] [CrossRef]
- Rau, J.; Korte, N.; Dyk, M.; Wenninger, O.; Schreiter, P.; Hiller, E. Rapid animal species identification of feta and mozzarella cheese using MALDI-TOF mass-spectrometry. Food Control 2020, 117, 107349. [Google Scholar] [CrossRef]
- Tian, H.; Zheng, N.; Wang, W.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Integrated Metabolomics Study of the Milk of Heat-stressed Lactating Dairy Cows. Sci. Rep. 2016, 6, 24208. [Google Scholar] [CrossRef]
- Xu, W.; Vervoort, J.; Saccenti, E.; van Hoeij, R.; Kemp, B.; van Knegsel, A. Milk metabolomics data reveal the en-ergy balance of individual dairy cows in early lactation. Sci. Rep. 2018, 8, 15828. [Google Scholar] [CrossRef] [Green Version]
- Fontanesi, L. Metabolomics and livestock genomics: Insights into a phenotyping frontier and its applications in ani-mal breeding. Anim. Front. 2016, 6, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Boudonck, K.J.; Mitchell, M.W.; Wulff, J.; Ryals, J.A. Characterization of the biochemical variability of bovine milk using metabolomics. Metabolomics 2009, 5, 375–386. [Google Scholar] [CrossRef]
- Klein, M.S.; Buttchereit, N.; Miemczyk, S.P.; Immervoll, A.-K.; Louis, C.; Wiedemann, S.; Junge, W.; Thaller, G.; Oefner, P.J.; Gronwald, W. NMR Metabolomic Analysis of Dairy Cows Reveals Milk Glycerophosphocholine to Phosphocholine Ratio as Prognostic Biomarker for Risk of Ketosis. J. Proteome Res. 2011, 11, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Zou, C.; Lin, B.; Chen, Y.; Liang, X.; Liu, J. A Comparison of Milk Protein, Amino Acid and Fatty Acid Profiles of River Buffalo and Their F1 and F2 Hybrids with Swamp Buffalo in China. Pak. J. Zool. 2015, 47, 1459–1465. [Google Scholar]
- Villaseñor, A.; Garcia-Perez, I.; Garcia, A.; Posma, J.M.; Fernández-López, M.; Nicholas, A.J.; Modi, N.; Holmes, E.; Barbas, C. Breast Milk Metabolome Characterization in a Single-Phase Extraction, Multiplatform Analytical Approach. Anal. Chem. 2014, 86, 8245–8252. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tong, J.; Zhang, Y.; Xiong, B.; Jiang, L. Metabolomics reveals potential biomarkers in the rumen fluid of dairy cows with different levels of milk production. Asian-Australas. J. Anim. Sci. 2020, 33, 79–90. [Google Scholar] [CrossRef]
- Metaboanalyst 4.0 Software. Available online: http://www.metaboanalyst.ca (accessed on 21 July 2020).
- The Kyoto Encyclopedia of Genes and Genomes Database. Available online: https://www.kegg.jp (accessed on 21 July 2020).
- The Human Metabolome Database. Available online: http://www.hmdb.ca (accessed on 1 September 2020).
- The Chemical Entities of Biological Interest. Available online: https://www.ebi.ac.uk (accessed on 5 June 2020).
- The Metabolite and Tandem MS Database. Available online: https://metlin.scripps.edu (accessed on 2 October 2020).
- The PubChem database. Available online: https://pubchem.ncbi.nlm.nih.gov (accessed on 16 July 2020).
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haile, A.; Hilali, M.; Hassen, H.; Lobo, R.N.B.; Rischkowsky, B. Estimates of genetic parameters and genetic trends for growth, reproduction, milk production and milk composition traits of Awassi sheep. Animals 2019, 13, 1374. [Google Scholar] [CrossRef] [PubMed]
- Selvaggi, M.; D’Alessandro, A.G.; Dario, C. Environmental and genetic factors affecting milk yield and quality in three Italian sheep breeds. J. Dairy Res. 2016, 84, 27–31. [Google Scholar] [CrossRef]
- Laben, R. Factors Responsible for Variation in Milk Composition. J. Dairy Sci. 1963, 46, 1293–1301. [Google Scholar] [CrossRef]
- Pasquini, M.; Osimani, A.; Tavoletti, S.; Moreno, I.; Clementi, F.; Trombetta, M.F. Trends in the quality and hy-giene parameters of bulk Italian Mediterranean buffalo (Bubalus bubalis) milk: A three year study. Anim. Sci. J. 2018, 89, 176–185. [Google Scholar] [CrossRef]
- Yang, T.X.; Li, H.; Wang, F.; Liu, X.L.; Li, Q.Y. Effect of cattle breeds on milk composition and technological char-acteristics in china. Asian Austral. J. Anim. 2013, 26, 896. [Google Scholar]
- Yue, S.; Ding, S.; Zhou, J.; Yang, C.; Hu, X.; Zhao, X.; Wang, Z.; Wang, L.; Peng, Q.; Xue, B. Metabolomics Approach Explore Diagnostic Biomarkers and Metabolic Changes in Heat-Stressed Dairy Cows. Animals 2020, 10, 1741. [Google Scholar] [CrossRef]
- de Nicola, D.; Vinale, F.; Salzano, A.; D Errico, G.; Vassetti, A.; D Onofrio, N.; Balestrieri, M.L.; Neglia, G. Milk Metabolomics Reveals Potential Biomarkers for Early Prediction of Pregnancy in Buffaloes Having Undergone Artificial Insemination. Animals 2020, 10, 758. [Google Scholar] [CrossRef]
- Osorio, J.S.; Lohakare, J.; Bionaz, M. Biosynthesis of milk fat, protein, and lactose: Roles of transcriptional and post-transcriptional regulation. Physiol. Genom. 2016, 48, 231–256. [Google Scholar] [CrossRef] [Green Version]
- Guijas, C.; Montenegro-Burke, J.R.; Warth, B.; Spilker, M.E.; Siuzdak, G. Metabolomics activity screening for iden-tifying metabolites that modulate phenotype. Nat. Biotechnol. 2018, 36, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Reue, K. Biochemistry, physiology, and genetics of GPAT, AGPAT, and lipin enzymes in triglyceride synthesis. Am. J. Physiol. Metab. 2009, 296, E1195–E1209. [Google Scholar] [CrossRef] [Green Version]
- Bernard, L.; Leroux, C.; Chilliard, Y. Expression and nutritional regulation of lipogenic genes in the ruminant lactating mammary gland. In Bioactive Components of Milk; Springer: Berlin/Heidelberg, Germany, 2008; pp. 67–108. [Google Scholar]
- Yonezawa, T.; Yonekura, S.; Kobayashi, Y.; Hagino, A.; Katoh, K.; Obara, Y. Effects of Long-Chain Fatty Acids on Cytosolic Triacylglycerol Accumulation and Lipid Droplet Formation in Primary Cultured Bovine Mammary Epithelial Cells. J. Dairy Sci. 2004, 87, 2527–2534. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Lu, S.; Cui, K.; Shafique, L.; Rehman, S.U.; Luo, C.; Wang, Z.; Ruan, J.; Qian, Q.; Liu, Q. Fatty acid biosynthe-sis and transcriptional regulation of Stearoyl-CoA Desaturase 1 (SCD1) in buffalo milk. BMC Genet. 2020, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Luo, J.; Zhang, Z.; Wu, J.; Zhang, T.; Busato, S.; Huang, L.; Song, N.; Bionaz, M. CRISPR/Cas9-mediated Stearoyl-CoA Desaturase 1 (SCD1) Deficiency Affects Fatty Acid Metabolism in Goat Mammary Epithelial Cells. J. Agric. Food Chem. 2018, 66, 10041–10052. [Google Scholar] [CrossRef]
- Keating, A.F.; Kennelly, J.J.; Zhao, F. Characterization and regulation of the bovine stearoyl-CoA desaturase gene promoter. Biochem. Bioph. Res. Commun. 2006, 344, 233–240. [Google Scholar] [CrossRef]
- Nakajima, S.; Gotoh, M.; Fukasawa, K.; Murakami-Murofushi, K.; Kunugi, H. Oleic acid is a potent inducer for lipid droplet accumulation through its esterification to glycerol by diacylglycerol acyltransferase in primary cortical as-trocytes. Brain Res. 2019, 1725, 146484. [Google Scholar] [CrossRef] [PubMed]
- Malodobra-Mazur, M.; Cierzniak, A.; Dobosz, T. Oleic acid influences the adipogenesis of 3T3-L1 cells via DNA Meth-ylation and may predispose to obesity and obesity-related disorders. Lipids Health Dis. 2019, 18, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.T.; Nadeem, M.; Imran, M.; Asif, M.; Khan, M.K.; Din, A.; Ullah, R. Triglyceride, fatty acid profile and an-tioxidant characteristics of low melting point fractions of Buffalo Milk fat. Lipids Health Dis. 2019, 18, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, A.; Banu, J.; Rahman, M.; Causey, J.; Fernandes, G. Biological effects of conjugated linoleic acids in health and disease. J. Nutr. Biochem. 2006, 17, 789–810. [Google Scholar] [CrossRef] [PubMed]
- Dipasquale, D.; Basiricò, L.; Morera, P.; Primi, R.; Tröscher, A.; Bernabucci, U. Anti-inflammatory effects of conju-gated linoleic acid isomers and essential fatty acids in bovine mammary epithelial cells. Anim. Int. J. Anim. Biosci. 2018, 12, 2108–2114. [Google Scholar] [CrossRef]
- Zhang, H.; Dan, N.; Ao, C.; Wang, S.; Erdene, K.; Ashraf, M.U. Effects of exogenous C18 unsaturated fatty acids on milk lipid synthesis in bovine mammary epithelial cells. J. Dairy Res. 2020, 87, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Sheng, R.; Yan, S.M.; Qi, L.Z.; Zhao, Y.L. Effect of the ratios of unsaturated fatty acids on the expressions of genes related to fat and protein in the bovine mammary epithelial cells. Vitr. Cell. Dev. Biol. Anim. 2015, 51, 381–389. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breeds | Fat/% | Protein/% | Lactose/% | Total Solids/% |
|---|---|---|---|---|
| Mediterranean | 8.40 ± 0.27 a | 4.32 ± 0.13 a | 5.12 ± 0.07 a | 19.05 ± 0.38 a |
| Murrah | 7.37 ± 0.32 b | 4.69 ± 0.12 ab | 5.13 ± 0.07 a | 17.26 ± 0.51 b |
| Crossbred | 6.87 ± 0.31 b | 4.82 ± 0.15 b | 5.24 ± 0.04 a | 17.44 ± 0.39 b |
| Pathway Name | KEGG Map ID |
|---|---|
| Biosynthesis of unsaturated fatty acids | map01040 |
| Fatty acid biosynthesis | map00061 |
| Primary bile acid biosynthesis ∗ | map00120 |
| Linoleic acid metabolism | map00591 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.; Yuan, X.; Cui, K.; Li, H.; Fu, P.; Rehman, S.-U.; Shi, D.; Liu, Q.; Li, Z. LC-MS/MS Based Metabolomics Reveal Candidate Biomarkers and Metabolic Changes in Different Buffalo Species. Animals 2021, 11, 560. https://doi.org/10.3390/ani11020560
Shi W, Yuan X, Cui K, Li H, Fu P, Rehman S-U, Shi D, Liu Q, Li Z. LC-MS/MS Based Metabolomics Reveal Candidate Biomarkers and Metabolic Changes in Different Buffalo Species. Animals. 2021; 11(2):560. https://doi.org/10.3390/ani11020560
Chicago/Turabian StyleShi, Wen, Xiang Yuan, Kuiqing Cui, Hui Li, Penghui Fu, Saif-Ur Rehman, Deshun Shi, Qingyou Liu, and Zhipeng Li. 2021. "LC-MS/MS Based Metabolomics Reveal Candidate Biomarkers and Metabolic Changes in Different Buffalo Species" Animals 11, no. 2: 560. https://doi.org/10.3390/ani11020560
APA StyleShi, W., Yuan, X., Cui, K., Li, H., Fu, P., Rehman, S. -U., Shi, D., Liu, Q., & Li, Z. (2021). LC-MS/MS Based Metabolomics Reveal Candidate Biomarkers and Metabolic Changes in Different Buffalo Species. Animals, 11(2), 560. https://doi.org/10.3390/ani11020560