
Distress Regulates Different Pathways in the Brain of Common Carp: A Preliminary Study



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing Conditions and Sampling
2.2. PCR Conditions
2.3. Calculations and Statistics
3. Results
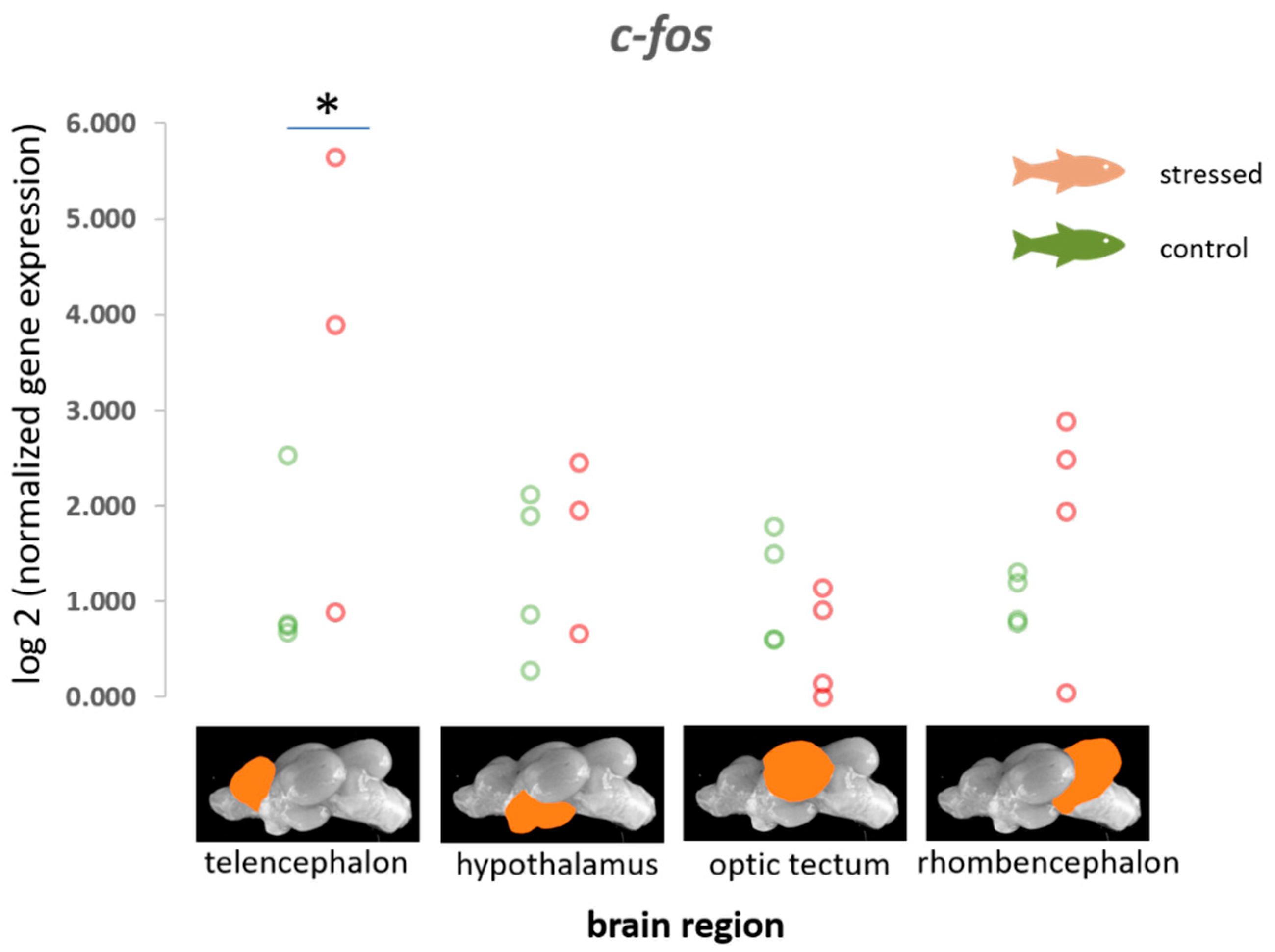
3.1. Immediate Early Genes (c-fos, egr-1, erk-1 and erk-2, palld) and gapdh
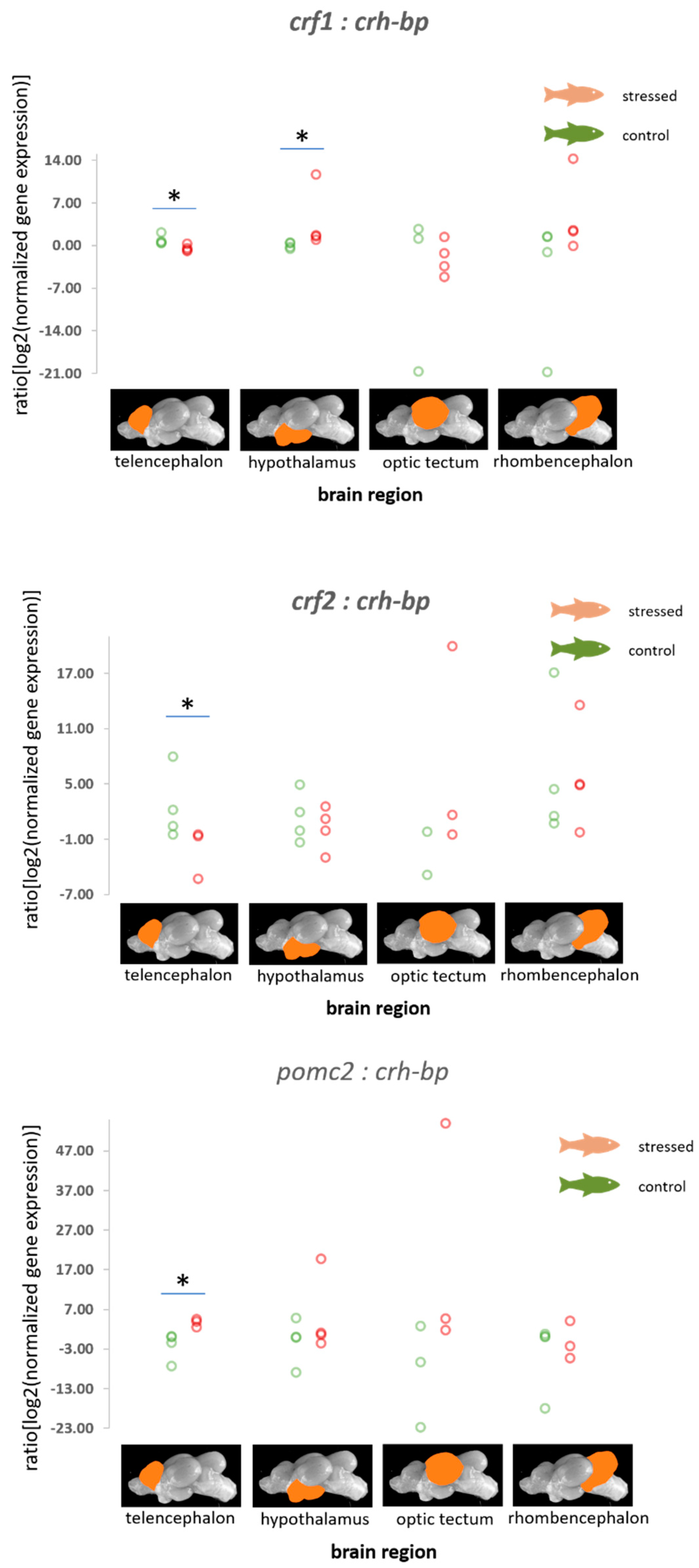
3.2. HPI Axis-Related Genes
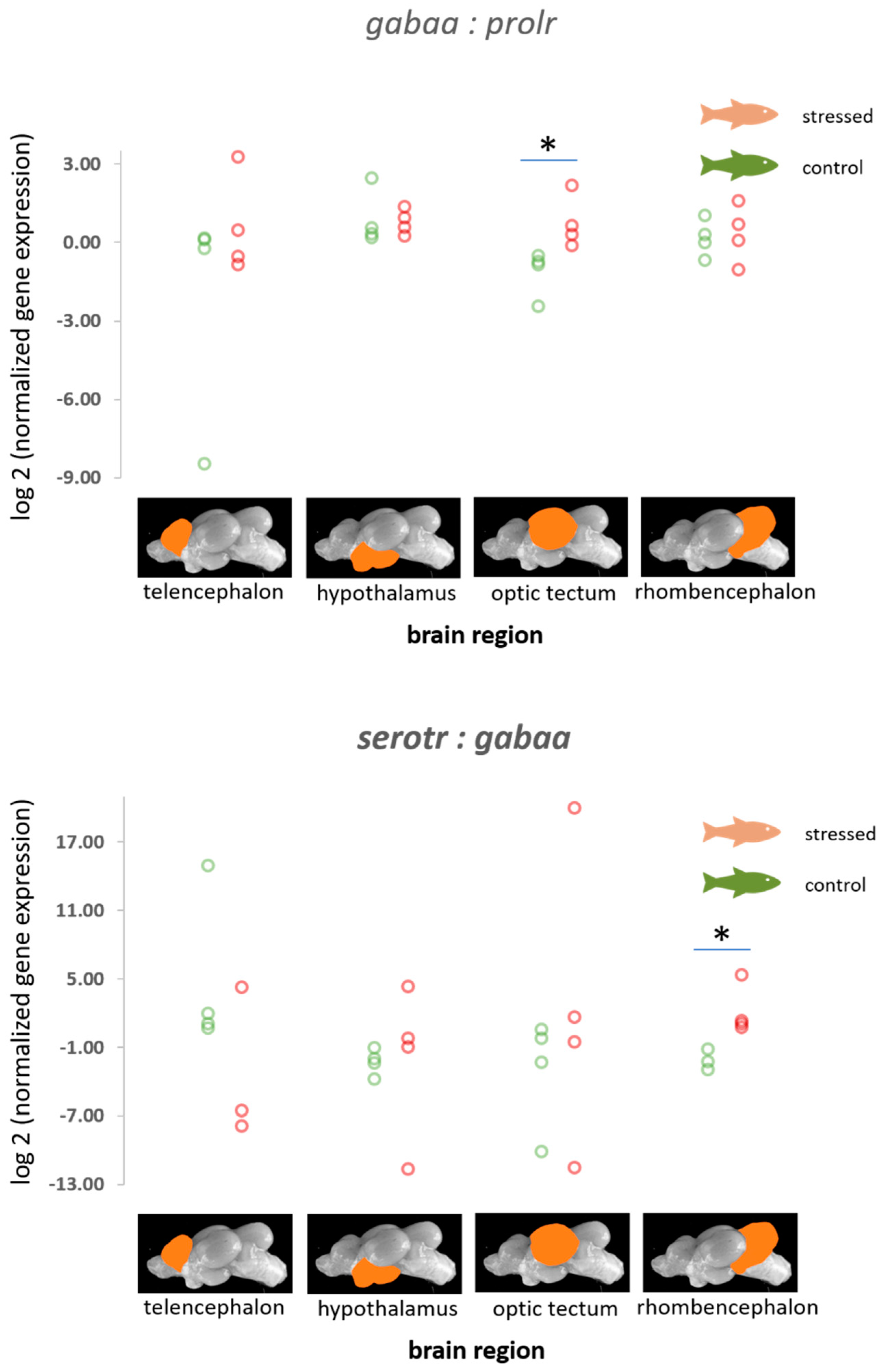
3.3. The Gene Expression Patterns of the Serotonergic Genes, gabaa, Isotocin Precursor, and the Prolactin Receptor
4. Discussion
4.1. Immediate Early Genes (IEGs)
4.2. HPI Axis-Related Genes
4.3. Genes of the Serotonergic and the GABAergic Pathway
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Moberg, G.P. Problems in defining stress and distress in animals. J. Am. Veter. Med. Assoc. 1987, 191, 1207–1211. [Google Scholar]
- Carstens, E.; Moberg, G.P. Recognizing pain and distress in laboratory animals. ILAR J. 2000, 41, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Gornati, R.; Papis, E.; Rimoldi, S.; Terova, G.; Saroglia, M.; Bernardini, G. Rearing density influences the expression of stress-related genes in sea bass (Dicentrarchus labrax, L.). Gene 2004, 341, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Gao, G.; Palti, Y.; Cleveland, B.M.; Weber, G.M.; Rexroad, C.E. RNA-seq analysis of early hepatic response to handling and confinement stress in rainbow trout. PLoS ONE 2014, 9, e88492. [Google Scholar] [CrossRef]
- Sun, P.; Yin, F.; Tang, B. Effects of acute handling stress on expression of growth-related genes in Pampus argenteus. J. World Aquac. Soc. 2016, 48, 166–179. [Google Scholar] [CrossRef]
- Samaras, A.; Santo, C.E.; Papandroulakis, N.; Mitrizakis, N.; Pavlidis, M.; Höglund, E.; Pelgrim, T.N.M.; Zethof, J.; Spanings, F.A.T.; Vindas, M.A.; et al. Allostatic load and stress physiology in European seabass (Dicentrarchus labrax L.) and gilthead seabream (Sparus aurata L.). Front. Endocrinol. 2018, 9, 451. [Google Scholar] [CrossRef] [Green Version]
- Vindas, M.A.; Gorissen, M.; Höglund, E.; Flik, G.; Tronci, V.; Damsgård, B.; Thörnqvist, P.O.; Nilsen, T.O.; Winberg, S.; Øverli, Ø.; et al. How do individuals cope with stress? Behavioural, physiological and neuronal differences between proactive and reactive coping styles in fish. J. Exp. Biol. 2017, 220, 1524–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronson, L.R. Forebrain function in teleost fishes. Trans. N. Y. Acad. Sci. 1967, 29, 390–396. [Google Scholar] [CrossRef]
- Von Trotha, J.W.; Vernier, P.; Bally-Cuif, L. Emotions and motivated behavior converge on an amygdala-like structure in the zebrafish. Eur. J. Neurosci. 2014, 40, 3302–3315. [Google Scholar] [CrossRef] [Green Version]
- Lal, P.; Tanabe, H.; Suster, M.L.; Ailani, D.; Kotani, Y.; Muto, A.; Itoh, M.; Iwasaki, M.; Wada, H.; Yaksi, E.; et al. Identification of a neuronal population in the telencephalon essential for fear conditioning in zebrafish. BMC Biol. 2018, 16, 1–18. [Google Scholar] [CrossRef]
- Lau, B.Y.B.; Mathur, P.; Gould, G.G.; Guo, S. Identification of a brain center whose activity discriminates a choice behavior in zebrafish. Proc. Natl. Acad. Sci. USA 2011, 108, 2581–2586. [Google Scholar] [CrossRef] [Green Version]
- Paton, J.J.; Belova, M.A.; Morrison, S.E.; Salzman, C.D. The primate amygdala represents the positive and negative value of visual stimuli during learning. Nat. Cell Biol. 2006, 439, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Murray, E.A. The amygdala, reward and emotion. Trends Cogn. Sci. 2007, 11, 489–497. [Google Scholar] [CrossRef]
- Morrison, S.E.; Salzman, C.D. Re-valuing the amygdala. Curr. Opin. Neurobiol. 2010, 20, 221–230. [Google Scholar] [CrossRef]
- Johansen, J.P.; Cain, C.K.; Ostroff, L.E.; LeDoux, J.E. Molecular mechanisms of fear learning and memory. Cell 2011, 147, 509–524. [Google Scholar] [CrossRef] [Green Version]
- Ingle, D.J.; Irwin, L.N. Optic Tectum. In Comparative Neuroscience and Neurobiology. Readings from the Encyclopedia of Neuroscience; Birkhäuser: Boston, MA, USA, 1988; pp. 100–101. [Google Scholar]
- Springer, A.D.; Easter, S.S.; Agranoff, B.W. The role of the optic tectum in various visually mediated behaviors of goldfish. Brain Res. 1977, 128, 393–404. [Google Scholar] [CrossRef]
- Roeser, T.; Baier, H. Visuomotor behaviors in larval zebrafish after GFP-guided laser ablation of the optic tectum. J. Neurosci. 2003, 23, 3726–3734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.W.; Elmquist, J.K. From neuroanatomy to behavior: Central integration of peripheral signals regulating feeding behavior. Nat. Neurosci. 2012, 15, 1350–1355. [Google Scholar] [CrossRef] [Green Version]
- Moens, C.B.; Prince, V.E. Constructing the hindbrain: Insights from the zebrafish. Dev. Dyn. 2002, 224, 1–17. [Google Scholar] [CrossRef]
- Duran, E.; Ocaña, F.M.; Martín-Monzón, I.; Rodríguez, F.; Salas, C. Cerebellum and spatial cognition in goldfish. Behav. Brain Res. 2014, 259, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Adams, D.H.; Simen, B.B. Transcription factors as modulators of stress responsivity. In Handbook of Molecular-Genetic Techniques for Brain and Behavior Research; Elsevier Science: Amsterdam, The Netherlands, 2005; Volume 15, pp. 679–698. [Google Scholar]
- Ashlin, T.G.; Blunsom, N.J.; Ghosh, M.; Cockcroft, S.; Rihel, J. Pitpnc1a regulates zebrafish sleep and wake behavior through modulation of insulin-like growth factor signaling. Cell Rep. 2018, 24, 1389–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, D.; Tran, S.; Shams, S.; Gerlai, R. A simple method for immunohistochemical staining of zebrafish brain sections for c-fos protein expression. Zebrafish 2015, 12, 414–420. [Google Scholar] [CrossRef]
- Salierno, J.; Snyder, N.; Murphy, A.; Poli, M.; Hall, S.; Baden, D.; Kane, A. Harmful algal bloom toxins alter c-Fos protein expression in the brain of killifish, Fundulus heteroclitus. Aquat. Toxicol. 2006, 78, 350–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, M.; Bell, A.M. Changes in behavior and brain immediate early gene expression in male threespined sticklebacks as they become fathers. Horm. Behav. 2018, 97, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Randlett, O.; Wee, C.L.; Naumann, E.A.; Nnaemeka, O.; Schoppik, D.; Fitzgerald, J.E.; Portugues, R.; Lacoste, A.M.; Riegler, C.; Engert, F.; et al. Whole-brain activity mapping onto a zebrafish brain atlas. Nat. Methods 2015, 12, 1039–1046. [Google Scholar] [CrossRef] [Green Version]
- Boukhelifa, M.; Parast, M.M.; Valtschanoff, J.G.; LaMantia, A.S.; Meeker, R.B.; Otey, C.A. A role for the cytoskeleton-associated protein palladin in neurite outgrowth. Mol. Biol. Cell. 2001, 12, 2721–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Chen, L.; Ge, Y.; Li, S.; Peng, W.; Li, C.; Zhang, Y.; Hu, X.; Zhou, Z.; Shi, L.; et al. Genetic mapping of Koi herpesvirus resistance (KHVR) in mirror carp (Cyprinus carpio) revealed genes and molecular mechanisms of disease resistance. Aquaculture 2020, 519, 734850. [Google Scholar] [CrossRef]
- Artelt, N.; Ludwig, T.A.; Rogge, H.; Kavvadas, P.; Siegerist, F.; Blumenthal, A.; Brandt, J.V.D.; Otey, C.A.; Bang, M.-L.; Amann, K.; et al. The role of palladin in podocytes. J. Am. Soc. Nephrol. 2018, 29, 1662–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flik, G.; Klaren, P.H.; Burg, E.H.V.D.; Metz, J.R.; Huising, M.O. CRF and stress in fish. Gen. Comp. Endocrinol. 2006, 146, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Conde-Sieira, M.; Chivite, M.; Míguez, J.M.; Soengas, J.L. Stress effects on the mechanisms regulating appetite in teleost fish. Front. Endocrinol. 2018, 9, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyon, C.; Trudeau, V.; Moon, T.W. Stress elevates corticotropin-releasing factor (CRF) and CRF-binding protein mRNA levels in rainbow trout (Oncorhynchus mykiss). J. Endocrinol. 2005, 186, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Huising, M.; Van Schooten, C.; Taverne-Thiele, A.J.; Hermsen, T.; Kemenade, B.M.L.V.-V.; Flik, G.; Metz, J.R. Structural characterisation of a cyprinid (Cyprinus carpio L.) CRH, CRH-BP and CRH-R1, and the role of these proteins in the acute stress response. J. Mol. Endocrinol. 2004, 32, 627–648. [Google Scholar] [CrossRef] [Green Version]
- Seasholtz, A.F.; Valverde, R.A.; Denver, R.J. Corticotropin-releasing hormone-binding protein: Biochemistry and function from fishes to mammals. J. Endocrinol. 2002, 175, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Sturm, A.; Bury, N.; Dengreville, L.; Fagart, J.; Flouriot, G.; Rafestin-Oblin, M.E.; Prunet, P. 11-deoxycorticosterone is a potent agonist of the rainbow trout (Oncorhynchus mykiss) mineralocorticoid receptor. Endocrinology 2005, 146, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Stolte, E.H.; De Mazon, A.F.; Leon-Koosterziel, K.M.; Jesiak, M.; Bury, N.R.; Sturm, A.; Savelkoul, H.F.J.; Van Kemenade, B.M.L.V.; Flik, G. Corticosteroid receptors involved in stress regulation in common carp, Cyprinus carpio. J. Endocrinol. 2008, 198, 403–417. [Google Scholar] [CrossRef] [Green Version]
- Gesto, M.; López-Patiño, M.A.; Hernández, J.; Soengas, J.L.; Míguez, J.M. Gradation of the stress response in rainbow trout exposed to stressors of different severity: The role of brain serotonergic and dopaminergic systems. J. Neuroendocr. 2014, 27, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Pantoja, C.; Hoagland, A.; Carroll, E.; Schoppik, D.; Isacoff, E.Y. Measuring behavioral individuality in the acoustic startle behavior in zebrafish. Bio-Protocol 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlsen, R.I.; Pilowsky, L.S. The place of partial agonism in psychiatry: Recent developments. J. Psychopharmacol. 2005, 19, 408–413. [Google Scholar] [CrossRef]
- Kumar, S.; Porcu, P.; Werner, D.F.; Matthews, D.B.; Diaz-Granados, J.L.; Helfand, R.S.; Morrow, A.L. The role of GABAA receptors in the acute and chronic effects of ethanol: A decade of progress. Psychopharmacology 2009, 205, 529–564. [Google Scholar] [CrossRef] [Green Version]
- Abreu, M.S.; Giacomini, A.C.V.; Gusso, D.; Rosa, J.G.; Koakoski, G.; Kalichak, F.; Idalêncio, R.; Oliveira, T.A.; Barcellos, H.H.; Bonan, C.D.; et al. Acute exposure to waterborne psychoactive drugs attract zebrafish. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2016, 179, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Sokołowska, E.; Gozdowska, M.; Kulczykowska, E. Nonapeptides arginine, vasotocin and isotocin in fishes: Advantage of bioactive molecules measurement. Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Manzon, L.A. The role of prolactin in fish osmoregulation: A review. Gen. Comp. Endocrinol. 2002, 125, 291–310. [Google Scholar] [CrossRef] [Green Version]
- Power, D. Developmental ontogeny of prolactin and its receptor in fish. Gen. Comp. Endocrinol. 2005, 142, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Patil, M.J.; Henry, M.A.; Akopian, A.N. Prolactin receptor in regulation of neuronal excitability and channels. Channels 2014, 8, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grattan, D.R. 60 YEARS OF NEUROENDOCRINOLOGY: The hypothalamo-prolactin axis. J. Endocrinol. 2015, 226, T101–T122. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, Y.; Kozono, K.; Esaka, M.; Iijima, N.; Nagamatsu, Y.; Yoshida, M.; Uematsu, K. Molecular cloning and effect of c-fos mRNA on pharmacological stimuli in the goldfish brain. Comp. Biochem. Physiol. Part. D Genom. Proteom. 2006, 1, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, D.K. Evidence for prolactin feedback actions on hypothalamic oxytocin, vasoactive intestinal peptide and dopamine secretion. Neuroendocrinology 1989, 49, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Prunet, P.; Gonnard, J.-F.; Paboeuf, G. GABA-ergic control of prolactin release in rainbow trout (Oncorhynchus mykiss) pituitaries in vitro. Fish. Physiol. Biochem. 1993, 11, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, S.C.; Nadeau, K.; Abbasi, M.; Lachance, C.; Nguyen, M.; Fenrich, J. The ultimate qPCR experiment: Producing publication quality, reproducible data the first time. Trends Biotechnol. 2019, 37, 761–774. [Google Scholar] [CrossRef] [Green Version]
- Zhu, A.; Srivastava, A.; Ibrahim, J.G.; Patro, R.; Love, M.I. Nonparametric expression analysis using inferential replicate counts. Nucleic Acids Res. 2019, 47, e105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bürkner, P.-C. Advanced Bayesian multilevel modeling with the R package brms. R J. 2018, 10, 395–411. [Google Scholar] [CrossRef]
- Remans, T.; Keunen, E.; Bex, G.J.; Smeets, K.; Vangronsveld, J.; Cuypers, A. Reliable gene expression analysis by reverse transcription-quantitative PCR: Reporting and minimizing the uncertainty in data accuracy. Plant. Cell 2014, 26, 3829–3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas-Hernandez, N.; Véliz, D.; Vega-Retter, C. Selection of suitable reference genes for gene expression analysis in gills and liver of fish under field pollution conditions. Sci. Rep. 2019, 9, 3459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Øvergård, A.-C.; Nerland, A.H.; Patel, S. Evaluation of potential reference genes for real time RT-PCR studies in Atlantic halibut (Hippoglossus hippoglossus, L.); during development, in tissues of healthy and NNV-injected fish, and in anterior kidney leucocytes. BMC Mol. Biol. 2010, 11, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasnov, A.; Koskinen, H.; Pehkonen, P.; Rexroad, C.E., III; Afanasyev, S.; Mölsä, H. Gene expression in the brain and kidney of rainbow trout in response to handling stress. BMC Genom. 2005, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Kocmarek, A.L.; Ferguson, M.M.; Danzmann, R.G. Differential gene expression in small and large rainbow trout derived from two seasonal spawning groups. BMC Genom. 2014, 15, 57. [Google Scholar] [CrossRef] [Green Version]
- Barber, R.D.; Harmer, D.W.; Coleman, R.A.; Clark, B.J. GAPDH as a housekeeping gene: Analysis of GAPDH mRNA expression in a panel of 72 human tissues. Physiol. Genom. 2005, 21, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Pavlidis, M.; Theodoridi, A.; Tsalafouta, A. Neuroendocrine regulation of the stress response in adult zebrafish, Danio rerio. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2015, 60, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Bernier, N.J. The corticotropin-releasing factor system as a mediator of the appetite-suppressing effects of stress in fish. Gen. Comp. Endocrinol. 2006, 146, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Karakatsouli, N.; Katsakoulis, P.; Leondaritis, G.; Kalogiannis, D.; Papoutsoglou, S.E.; Chadio, S.; Sakellaridis, N. Acute stress response of European sea bass Dicentrarchus labrax under blue and white light. Aquaculture 2012, 48–52. [Google Scholar] [CrossRef]
- Alderman, S.L.; Bernier, N.J. Localization of corticotropin-releasing factor, urotensin I, and CRF-binding protein gene expression in the brain of the zebrafish, Danio rerio. J. Comp. Neurol. 2007, 502, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Bernier, N.J.; Flik, G.; Klaren, P.H.M. Chapter 6: Regulation and contribution of the corticotropic, melanotropic and thyrotropic axes to the stress response in fishes. In Fish Physiology: Fish Neuroendocrinology; Academic Press: Cambridge, MA, USA, 2009; pp. 235–311. ISSN 1546-5098. [Google Scholar]
- Williams, T.A.; Bergstrome, J.C.; Scott, J.; Bernier, N.J. CRF and urocortin 3 protect the heart from hypoxia/reoxygenation-induced apoptosis in zebrafish. Am. J. Physiol. Integr. Comp. Physiol. 2017, 313, R91–R100. [Google Scholar] [CrossRef]
- Manuel, R.; Metz, J.R.; Flik, G.; Vale, W.W.; Huising, M.O. Corticotropin-releasing factor-binding protein (CRF-BP) inhibits CRF- and urotensin-I-mediated activation of CRF receptor-1 and -2 in common carp. Gen. Comp. Endocrinol. 2014, 202, 69–75. [Google Scholar] [CrossRef]
- Fu, Y.; Neugebauer, V. Differential mechanisms of CRF1 and CRF2 receptor functions in the amygdala in pain-related synaptic facilitation and behavior. J. Neurosci. 2008, 28, 3861–3876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, M.J.; Cerdá-Reverter, J.M.; Soengas, J.L. Hypothalamic integration of metabolic, endocrine, and circadian signals in fish: Involvement in the control of food intake. Front. Neurosci. 2017, 11, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tellez, R.; Gómez-Viquez, L.; Liy-Salmeron, G.; Meneses, A. GABA, glutamate, dopamine and serotonin transporters expression on forgetting. Neurobiol. Learn. Mem. 2012, 98, 66–77. [Google Scholar] [CrossRef]
- Sanacora, G.; Mason, G.F.; Rothman, D.L.; Krystal, J.H. Increased occipital cortex GABA concentrations in depressed patients after therapy with selective serotonin reuptake inhibitors. Am. J. Psychiatry 2002, 159, 663–665. [Google Scholar] [CrossRef] [PubMed]
- Moltesen, M.; Laursen, D.C.; Thörnqvist, P.-O.; Andersson, M.; Winberg, S.; Höglund, E. Effects of acute and chronic stress on telencephalic neurochemistry and gene expression in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 2016, 219, 3907–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meaney, M.J.; Szyf, M. Maternal care as a model for experience-dependent chromatin plasticity? Trends Neurosci. 2005, 28, 456–463. [Google Scholar] [CrossRef]
- Meaney, M.J.; Diorio, J.; Francis, D.; Weaver, S.; Yau, J.; Chapman, K.; Seckl, J.R. Postnatal handling increases the expression of cAMP-inducible transcription factors in the rat hippocampus: The effects of thyroid hormones and serotonin. J. Neurosci. 2000, 20, 3926–3935. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Tel * | Hyp ** | Opt *** | Rho **** |
|---|---|---|---|---|
| 18S RNA | 0.16 (0.59) −1–1.39, p = 0.602 | 0.29 (0.46) −0.5–1.31, p = 0.740 | 0.75 (0.83) −0.76–2.47, p = 0.824 | 0.35 (0.46) −0.52–1.28, p = 0.780 |
| 5-ht-r | 0.22 (0.60) −0.9–1.51, p = 0.646 | 0 (0.41) −0.87–0.86, p = 0.492 | 0.09 (0.82) 1.57–1.77, p = 0.540 | 0.03 (0.45) −0.87–0.91, p = 0.526 |
| eIF4E | −0.01 (0.60) −1.22–1.22, p = 0.496 | −0.48 (0.51) −1.66–0.31, p = 0.162 | −0.66 (0.85) −2.41–0.98, p = 0.210 | 0.03 (0.46) −0.90–0.96, p = 0.520 |
| b2m | 0.01 (0.58) −1.15–1.18, p = 0.515 | 0.09 (0.42) −0.75–1.01, p = 0.580 | −0.85 (0.82) −2.58–0.64, p = 0.144 | 0.15 (0.44) −0.72–1.07, p = 0.630 |
| bactin | 0.13 (0.60) −1.06–1.38, p = 0.584 | 0.14 (0.43) −0.67–1.12, p = 0.621 | 0.33 (0.82) −1.23–2.03, p = 0.650 | −0.08 (0.45) −1.00–0.80, p = 0.432 |
| c-fos | 0.91 (0.70) −0.26–2.40, p = 0.912 | 0.37 (0.48) −0.40–1.49, p = 0.783 | −1.5 (0.93) −3.41–0.21, p = 0.045 | 0.33 (0.46) −0.53–1.28, p = 0.758 |
| crf-1 | −0.05 (0.57) −1.26–1.07, p = 0.467 | 0.12 (0.43) −0.70–1.08, p = 0.611 | −0.31 (0.82) −1.97–1.28, p = 0.350 | 0.25 (0.47) −0.64–1.22, p = 0.698 |
| crf-2 | −0.87 (0.69) −2.31–0.27, p = 0.088 | 0.05 (0.44) −0.88–1.00, p = 0.546 | 1.16 (0.87) −0.45–2.93, p = 0.918 | 0.30 (0.47) −0.57–1.29, p = 0.734 |
| crf-r1 | 0.04 (0.57) −1.12–1.21, p = 0.520 | 0.21 (0.44) −0.58–1.20, p = 0.670 | 0.39 (0.80) −1.17–2.06, p = 0.686 | −0.37 (0.45) −1.32–0.47, p = 0.204 |
| crf-r2 | −0.11 (0.58) −1.31–1.02, p = 0.420 | 0.28 (0.46) −0.53–1.34, p = 0.727 | 0.10 (0.81) −1.50–1.68, p = 0.546 | −0.31 (0.47) −1.29–0.57, p = 0.260 |
| crh-bp | 0.39 (0.61) −0.72–1.72, p = 0.737 | 0 (0.41) −0.89–0.84, p = 0.494 | 0.08 (0.80) −1.49–1.67, p = 0.545 | −0.15 (0.46) −1.10–0.72, p = 0.372 |
| ef | −0.13 (0.59) −1.33–1.06, p = 0.414 | 0.12 (0.43) −0.68–1.09, p = 0.606 | 0.38 (0.81) −1.24–2.07, p = 0.688 | −0.1 (0.46) −1.03–0.81, p = 0.411 |
| egr-1 | −0.08 (0.59) −1.31–1.07, p = 0.444 | −0.04 (0.43) −0.95–0.84, p = 0.468 | −0.81 (0.85) −2.55–0.76, p = 0.160 | 0.16 (0.46) −0.77–1.12, p = 0.632 |
| erk-1 | −0.15 (0.57) −1.34–0.97, p = 0.395 | −0.23 (0.44) −1.25–0.52, p = 0.312 | 0.14 (0.79) −1.38–1.75, p = 0.567 | −0.22 (0.45) −1.15–0.63, p = 0.318 |
| erk−2 | 0.10 (0.57) −1.02–1.24, p = 0.570 | 0.27 (0.47) −0.55–1.36, p = 0.717 | 0.65 (0.82) −0.88–2.34, p = 0.788 | 0.02 (0.45) −0.90–0.92, p = 0.513 |
| gabaa | −0.07 (0.58) −1.22–1.09, p = 0.454 | −0.08 (0.42) −1.01–0.75, p = 0.426 | −1.62 (0.93) −3.55–0.07, p = 0.035 | −0.36 (0.46) −1.30–0.49, p = 0.214 |
| gapdh | 0.09 (0.58) −1.05–1.30, p = 0.564 | 0.41 (0.50) −0.38–1.56, p = 0.807 | 0.53 (0.82) −1.00–2.16, p = 0.745 | 0.04 (0.47) −0.93–0.99, p = 0.539 |
| gr1 | −0.07 (0.59) −1.33–1.04, p = 0.456 | −0.33 (0.46) −1.37–0.44, p = 0.230 | −0.90 (0.85) −2.59–0.68, p = 0.140 | −0.15 (0.45) −1.06–0.75, p = 373 |
| gr2 | 0 (0.59) −1.21–1.24, p = 0.490 | 0.16 (0.44) −0.66–1.12, p = 0.631 | 0.30 (0.81) −1.29–1.96, p = 0.649 | −0.24 (0.45) −1.17–0.65, p = 0.299 |
| isopre | 1.21 (0.79) −0.07–2.87, p = 0.953 | 0.12 (0.44) −0.74–1.08, p = 0.599 | −0.79 (0.85) 2.47–0.80, p = 0.170 | −0.02 (0.46) −0.07–2.87, p = 0.953 |
| mr | −0.31 (0.61) −1.64–0.89, p = 0.306 | −0.31 (0.47) −1.39–0.45, p = 0.256 | 0.20 (0.82) −1.42–1.83, p = 0.603 | −0.25 (0.47) −1.22–0.65, p = 0.298 |
| palld | 0.11 (0.55) −0.96–1.26, p = 0.577 | 0.20 (0.45) −0.61–1.22, 0.656 | −0.40 (0.83) −2.08–1.18, p = 0.313 | −0.16 (0.46) −1.10–0.76, p = 0.363 |
| pomc1 | 0.09 (0.60) −1.08–1.38, p = 0.546 | −0.39 (0.50) −1.58–0.39, p = 0.221 | 1.47 (0.93) −0.22–3.40, p = 952 | 1.68 (0.73) 0.28–3.13, p = 0.997 |
| pomc2 | 0.75 (0.71) −0.41–2.31, p = 0.867 | −0.22 (0.44) −1.23–0.56, p = 0.316 | 2.05 (1.12) 0.03–4.45, p = 0.977 | 1.09 (0.63) −0.01–2.40, p = 0.974 |
| prolr | −0.45 (0.61) −1.78–0.66, p = 0.233 | 0.14 (0.43) −0.67–1.07, p = 0.618 | −0.99 (0.89) −2.84–0.63, p = 0.118 | 0.11 (0.46) −0.77–1.07, p = 0.586 |
| serotr | −1.47 (0.87) −3.27–0.02, p = 0.033 | −0.61 (0.58) −1.96–0.21, p = 0.117 | −0.74 (0.85) −2.44–0.87, p = 0.194 | −0.32 (0.47) −1.30–0.54, p = 0.244 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burren, A.; Pietsch, C. Distress Regulates Different Pathways in the Brain of Common Carp: A Preliminary Study. Animals 2021, 11, 585. https://doi.org/10.3390/ani11020585
Burren A, Pietsch C. Distress Regulates Different Pathways in the Brain of Common Carp: A Preliminary Study. Animals. 2021; 11(2):585. https://doi.org/10.3390/ani11020585
Chicago/Turabian StyleBurren, Alexander, and Constanze Pietsch. 2021. "Distress Regulates Different Pathways in the Brain of Common Carp: A Preliminary Study" Animals 11, no. 2: 585. https://doi.org/10.3390/ani11020585
APA StyleBurren, A., & Pietsch, C. (2021). Distress Regulates Different Pathways in the Brain of Common Carp: A Preliminary Study. Animals, 11(2), 585. https://doi.org/10.3390/ani11020585