
Enteric Viral Infections among Domesticated South American Camelids: First Detection of Mammalian Orthoreovirus in Camelids

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. MRV Isolation and Characterization
2.2. Fecal Samples
2.3. Viral RNA Extraction
2.4. Viral Detection and Identification
3. Results
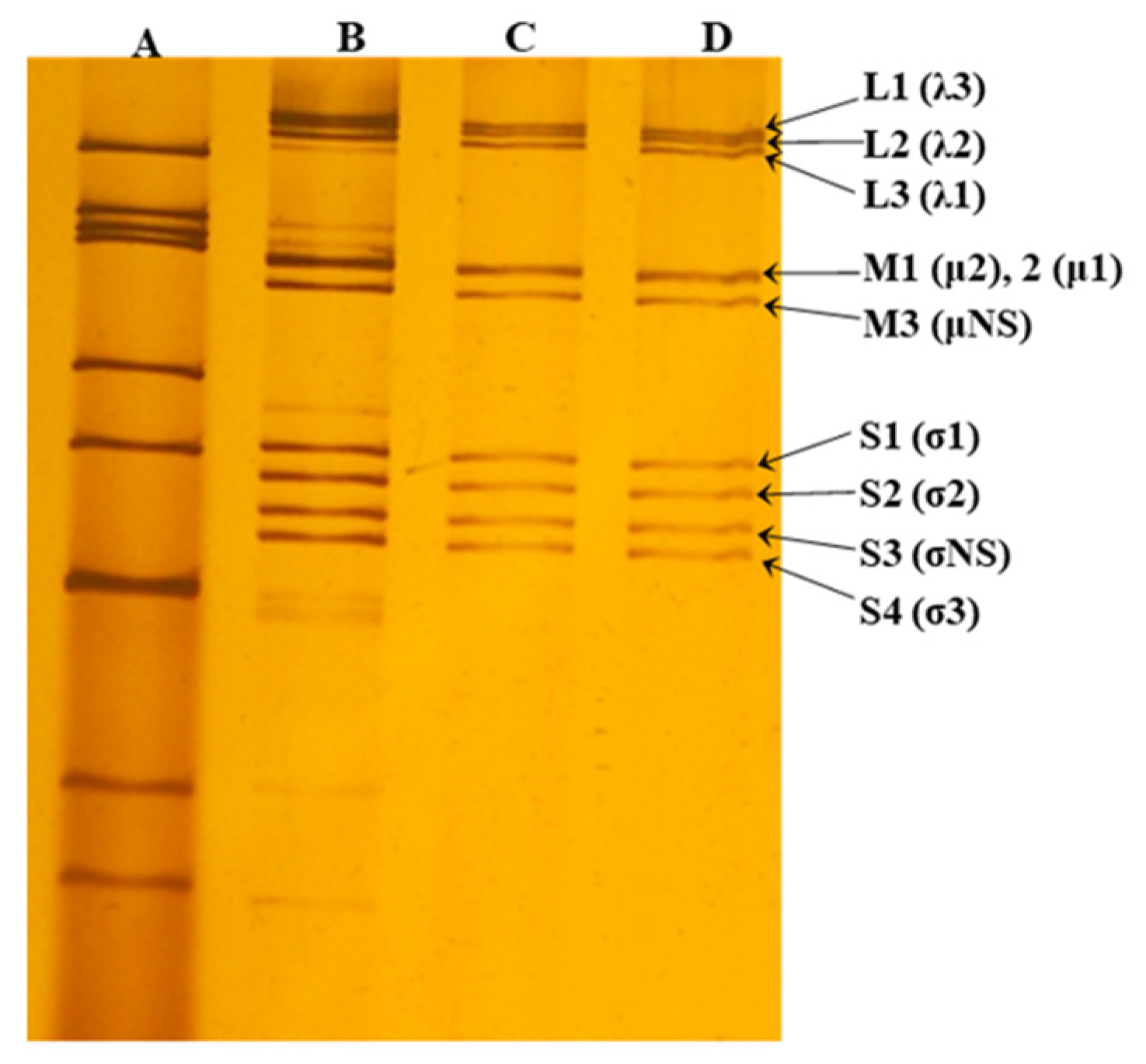
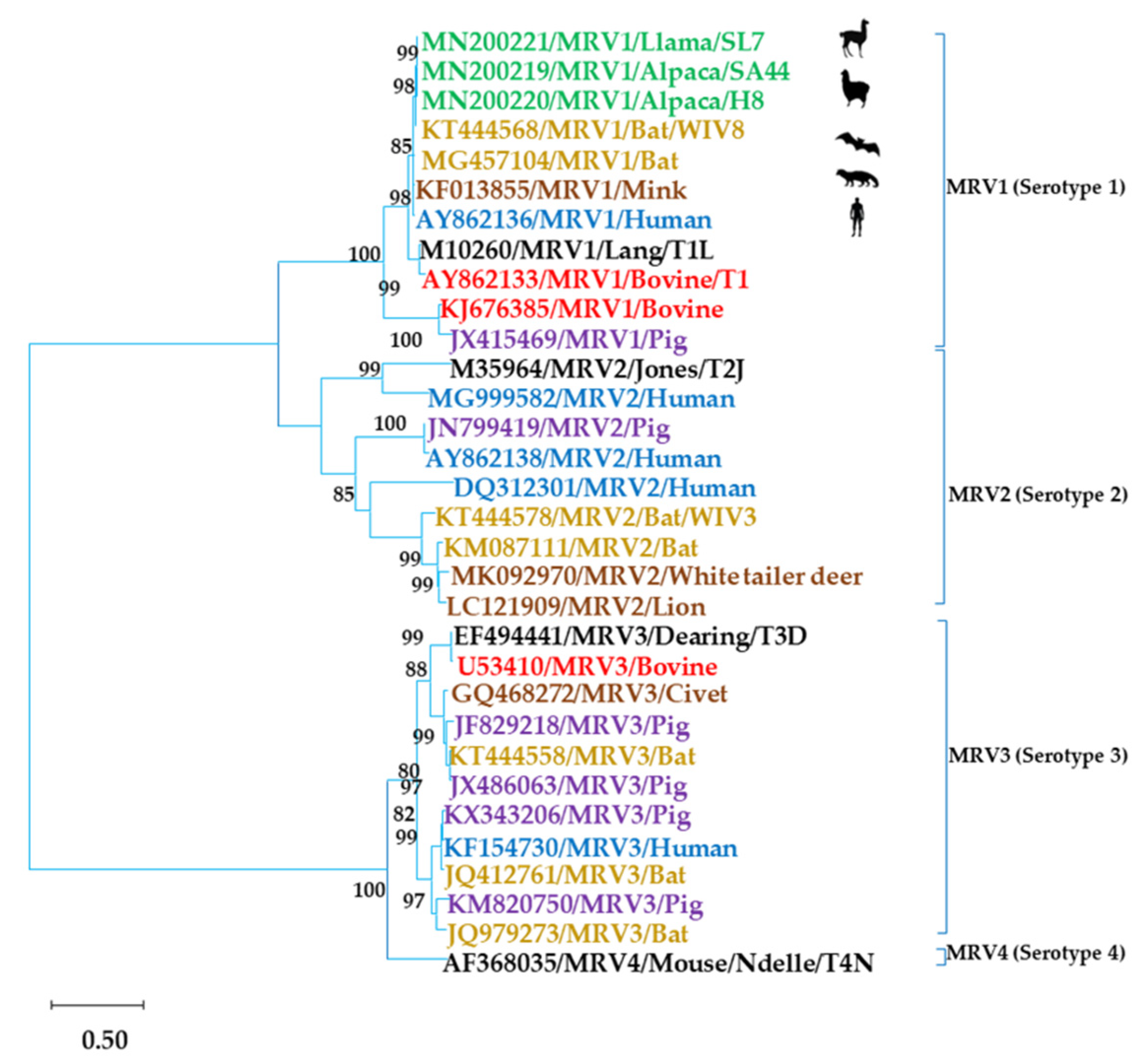
3.1. MRV Characterization by PAGE and Sequencing
3.2. Virus Detection
3.3. Identification of CoV Genus and Subgenus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ministério de Agricultura y Riego, Perú (MINAG). 2021. Available online: http://www.minag.gob.pe/portal/sector-agrario/pecuaria/situacion-de-las-actividades-de-crianza-y-produccion/cam%C3%A9lidos-sudamericanos (accessed on 31 March 2021).
- Consejo Nacional de Camélidos Sudamericanos (CONACS). Estrategia Nacional de Desarrollo: Los Camélideos Domésticos em El Perú. 2005. Available online: https://www.whyalpaca.com/assets/documentos/estrategia_nacional.pdf (accessed on 31 March 2021). (In Spanish).
- Food and Agriculture Organization of the United Nations (FAO). Situación Actual de los Camelidos Sudamericanos en el Perú. 2005. Available online: https://tarwi.lamolina.edu.pe/~emellisho/zootecnia_archivos/situacion%20alpcas%20peru.pdf (accessed on 31 March 2021). (In Spanish).
- Rosadio, A.R.; Maturrano, H.L.; Pérez, J.D.; Luna, E.L. El Complejo Entérico Neonatal en Alpacas Andinas. Rev. Investig. Vet. Perú 2012, 23, 261–271. [Google Scholar] [CrossRef]
- Martín Espada, M.; Pinto Jiménez, C.; Cid Vázquez, M. Camélidos Sudamericanos: Estado Sanitario de Sus Crías. Rev. Complut. Cienc. Vet. 2010, 4, 37–50. [Google Scholar]
- Rojas, M.; Manchego, A.; Rocha, C.B.; Fornells, L.A.; Silva, R.C.; Mendes, G.S.; Dias, H.G.; Sandoval, N.; Pezo, D.; Santos, N. Outbreak of Diarrhea among Preweaning Alpacas (Vicugna pacos) in the Southern Peruvian Highland. J. Infect. Dev. Ctries. 2016, 10, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Cebra, C.K.; Mattson, D.E.; Baker, R.J.; Sonn, R.J.; Dearing, P.L. Potential Pathogens in Feces from Unweaned Llamas and Alpacas with Diarrhea. J. Am. Vet. Med. Assoc. 2003, 223, 1806–1808. [Google Scholar] [CrossRef]
- Garmendia, A.E.; Lopez, W.; Ortega, N.; Chamorro, M.J. Molecular Characterization of Rotavirus Isolated from Alpaca (Vicugna pacos) Crias with Diarrhea in the Andean Region of Cusco, Peru. Vet. Microbiol. 2015, 180, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Wellington López, P.; Marycris Chamorro, L.; Antonio, E.; Garmendia, B. Detección Rápida de Rotavirus y Coronavirus en Crías de Alpaca (Vicugna pacos) con Diarrea en la Región del Cusco, Perú. Rev. Investig. Vet. Perú. 2011, 22, 407–411. [Google Scholar]
- Luna, L.; Brandão, P.E.; Maturrano, L.; Rosadio, R.; Silva, F.D.F.; Soares, R.M.; Gregori, F. Betacoronavirus 1 in Alpacas (Vicugna pacos) in the High Peruvian Andes. Small Rumin. Res. 2015, 133, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Puerta, L.A.; Gonzalez, A.E.; Vargas-Calla, A.; Lopez-Urbina, M.T.; Cama, V.; Xiao, L. Cryptosporidium parvum as a Risk Factor of Diarrhea Occurrence in Neonatal Alpacas in Peru. Parasitol. Res. 2020, 119, 243–248. [Google Scholar] [CrossRef]
- Rojas, M.A.; Gonçalves, J.L.S.; Dias, H.G.; Manchego, A.; Santos, N. Identification of Two Novel Rotavirus A Genotypes, G35 and P[50], from Peruvian Alpaca Faeces. Infect. Genet. Evol. 2017, 55, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Herring, A.J.; Inglis, N.F.; Ojeh, C.K.; Snodgrass, D.R.; Menzies, J.D. Rapid diagnosis of rotavirus infection by direct detection of viral nucleic acid in silver-stained polyacrylamide gels. J. Clin. Microbiol. 1982, 16, 473–477. [Google Scholar] [CrossRef] [Green Version]
- Moës, E.; Vijgen, L.; Keyaerts, E.; Zlateva, K.; Li, S.; Maes, P.; Pyrc, K.; Berkhout, B.; van der Hoek, L.; van Ranst, M. A Novel Pancoronavirus RT-PCR Assay: Frequent Detection of Human Coronavirus NL63 in Children Hospitalized with Respiratory Tract Infections in Belgium. BMC Infect. Dis. 2005, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Brandão, P.E. Coronavírus Bovino (BCoV): Ocorrência, Diversidade Molecular e Padronização de PCR para Diagnóstico a partir de Amostras Fecais de Bezerros com e sem Diarreia Criados em Municípios dos Estados de São Paulo e Minas Gerais, Brasil. Ph.D. Thesis, Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, Brazil, 4 April 2004. [Google Scholar]
- Matthijnssens, J.; Ciarlet, M.; Rahman, M.; Attoui, H.; Bányai, K.; Estes, M.K.; Gentsch, J.R.; Iturriza-Gómara, M.; Kirkwood, C.D.; Martella, V.; et al. Recommendations for the Classification of Group A Rotaviruses Using All 11 Genomic RNA Segments. Arch. Virol. 2008, 153, 1621–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, C.B.; Fornells, L.A.A.M.G.; Rojas, M.; Libetal, M.; Manchego, A.; Pezo, D.; Santos, N. Molecular Epidemiology of Coronavirus in Faeces of Brazilian Calves and Peruvian Camelid Herds. J. Infect. Dev. Ctries. 2018, 12, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Siever Morales, C.; Daniel Paredes, L.; Danilo Pezo, C. Asociación de Rotavirus y Escherichia coli Fimbriada como Agentes Causales de Infecciones Entéricas en Alpacas Neonatas. Rev. Investig. Vet. Perú. 2007, 18, 150–153. [Google Scholar]
- Quispe, E.; Rodríguez, T.; Iñiguez, L.; Mueller, J. Producción de Fibra de Alpaca, Llama, Vicuña y Guanaco en Sudamérica. Anim. Genet. Resour. Inf. 2009, 45, 1–14. [Google Scholar] [CrossRef]
- Hermoza, L.G. Situacion y Perspectivas del Desarrollo del Sub Sector Camelidos Peruanos. 2011. Available online: https://1library.co/document/q5ovmr3z-situacion-perspectivas-desarrollo-sub-sector-camelidos-peruanos.html (accessed on 31 March 2021). (In Spanish).
- Whitehead, C.E. Neonatal Diseases in Llamas and Alpacas. Vet. Clin. N. Am. Food Anim. Pract. 2009, 25, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Lucas, L.J.R.; Morales, C.S.; Barrios, A.M.; Rodríguez, G.J.; Vásquez, C.M.; Lira, M.B.; Torres, L.B.; Casas, A.E.; Espinoza, B.J. Patógenos Involucrados en Casos Fatales de Diarrea en Crías de Alpaca de la Sierra Central del Perú. Rev. Investig. Vet. Perú. 2016, 27, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Steyer, A.; Gutiérrez-Aguire, I.; Kolenc, M.; Koren, S.; Kutnjak, D.; Pokorn, M.; Poljšak-Prijatelj, M.; Rački, N.; Ravnikar, M.; Sagadin, M.; et al. High Similarity of Novel Orthoreovirus Detected in a Child Hospitalized with Acute Gastroenteritis to Mammalian Orthoreoviruses Found in Bats in Europe. J. Clin. Microbiol. 2013, 51, 3818–3825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anbalagan, S.; Spaans, T.; Hause, B.M. Genome Sequence of the Novel Reassortant Mammalian Orthoreovirus Strain MRV00304/13, Isolated from a Calf with Diarrhea from the United States. Genome Announc. 2014, 2, e00451-14. [Google Scholar] [CrossRef] [Green Version]
- Qin, P.; Li, H.; Wang, J.W.; Wang, B.; Xie, R.H.; Xu, H.; Zhao, L.-Y.; Li, L.; Pan, Y.; Song, Y.; et al. Genetic and Pathogenic Characterization of a Novel Reassortant Mammalian Orthoreovirus 3 (MRV3) from a Diarrheic Piglet and Seroepidemiological Survey of MRV3 in Diarrheic Pigs from East China. Vet. Microbiol. 2017, 208, 126–136. [Google Scholar] [CrossRef]
- Li, Z.; Shao, Y.; Liu, C.; Liu, D.; Guo, D.; Qiu, Z.; Tian, J.; Zhang, X.; Liu, S.; Qu, L. Isolation and pathogenicity of the mammalian orthoreovirus MPC/04 from masked civet cats. Infect. Genet. Evol. 2015, 36, 55–61. [Google Scholar] [CrossRef]
- Narayanappa, A.T.; Sooryanarain, H.; Deventhiran, J.; Cao, D.; Venkatachalam, B.A.; Kambiranda, D.; Kambiranda, D.; LeRoith, T.; Heffron, C.L.; Lindstrom, N.; et al. A Novel Pathogenic Mammalian Orthoreovirus from Diarrheic Pigs and Swine Blood Meal in the United States. MBio 2015, 6, e00593-15. [Google Scholar] [CrossRef] [Green Version]
- Chua, K.B.; Voon, K.; Crameri, G.; Tan, H.S.; Rosli, J.; McEachern, J.A.; Suluraju, S.; Yu, M.; Wang, L.-F. Identification and Characterization of a New Orthoreovirus from Patients with Acute Respiratory Infections. Schwartz O, editor. PLoS ONE 2008, 3, e3803. [Google Scholar] [CrossRef]
- Ahasan, M.S.; Subramaniam, K.; Sayler, K.A.; Loeb, J.C.; Popov, V.L.; Lednicky, J.A.; Wisely, S.M.; Krauer, J.M.C.; Waltzek, T.B. Molecular Characterization of a Novel Reassortment Mammalian Orthoreovirus Type 2 Isolated from a Florida White-Tailed Deer Fawn. Virus Res. 2019, 270, 197642. [Google Scholar] [CrossRef] [PubMed]
- Rosa, U.A.; Ribeiro, G.O.; Villanova, F.; Luchs, A.; Milagres, F.A.P.; Komninaki, S.V.; Tahmasebi, R.; Lobato, M.C.A.B.S.; Brustulin, R.; Chagas, R.T.; et al. First identification of Mammalian Orthoreovirus Type 3 by Gut Virome Analysis in Diarrheic Child in Brazil. Sci. Rep. 2019, 9, 18599. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Cebra, C.K.; Baker, R.J.; Mattson, D.E.; Cohen, S.A.; Alvarado, D.E.; Rohrmann, G.F. Analysis of the Genome Sequence of an Alpaca Coronavirus. Virology 2007, 365, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, M.; Dias, H.G.; Gonçalves, J.L.S.; Manchego, A.; Rosadio, R.; Pezo, D.; Santos, N. Genetic Diversity and Zoonotic Potential of Rotavirus A Strains in the Southern Andean Highlands, Peru. Transbound. Emerg. Dis. 2019, 66, 1718–1726. [Google Scholar] [CrossRef]
- Fung, T.S.; Liu, D.X. Human Coronavirus: Host-Pathogen Interaction. Annu. Rev. Microbiol. 2019, 73, 529–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dóró, R.; Farkas, S.L.; Martella, V.; Banyai, K. Zoonotic Transmission of Rotavirus: Surveillance and Control. Expert Rev. Anti Infect. Ther. 2015, 13, 1337–1350. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Virus | Gene | Assay | Primer * | Primer Sequence 5′→3′ | Position | Product Size (bp) | Reference |
|---|---|---|---|---|---|---|---|
| All CoV | RpRd + | RT-PCR | Cor-FW | ACWCARHTVAAYYTNAARTAYGC | 14,922–14,944 | 251 | [14] |
| Cor-RV | TCRCAYTTDGGRTARTCCCA | 15,153–15,172 | |||||
| BetaCoV | RpRd | Nested PCR | Beta.CoV.F | ATTAGTGCWAAGAATAGAGCYCGCAC | 14,946–14,971 | 227 | This study |
| Beta.CoV.R | TCACAYTTWGGRTARTCCCADCCCA | 15,148–15,172 | |||||
| Embecovirus | RpRd | Nested PCR | CV2U.F | TACTATGACTGGCAGAATGTTTCA | 14,996–15,019 | 136 | [15] |
| CV2L.R | AACATCTTTAATAAGGCGRCGTAA | 15,108–15,131 | |||||
| All MRV | RpRd | RT-PCR | MRV-FM | CCNATATCNGGAATGCAGAA | 1943–1962 | 181 | This study |
| MRV-RM | TCCATCATCGTRCTATTRTTNGC | 2102–2124 | |||||
| All RVA | NSP5 | RT-PCR | Gen_NSP5F | GGCTTTTAAAGCGCTACAG | 1–19 | 667 | [16] |
| Gen_NSP5R | GGTCACAAAACGGGAGT | 651–667 | |||||
| All RVA | NSP5 | Nested PCR | Max_1FM | CGTCAACTCTTTCTGGAAAATCTA | 95–121 | 562 | [12] |
| Max_4RM | GTGGGGAGCTCCCTAGT | 637–656 |
| Community/District | Geographic Coordinates | Host Species | No Tested Samples | No of Positive Samples (%) | ||||
|---|---|---|---|---|---|---|---|---|
| CoV | MRV | RVA | Coinfections † | Total | ||||
| Silly/Marangani | 14°21′12″ S, 71°10′17″ W, 3800 masl ± | Alpaca | 33 | 7 | 2 | 0 | 23 (16 CoV + MRV; 3 CoV + RVA; 1 MRV + RVA; 3 CoV + MRV + RVA) | 32 (97) |
| Quisini/Marangani | 14°39′72″ S, 71°10′89″ W, 4300 masl | Alpaca | 23 | 10 | 0 | 0 | 11 (4 CoV + MRV; 3 CoV + RVA; 4 CoV + MRV + RVA) | 21 (91.3) |
| Pataccalasaya/Sicuani | 14°16′9.5″ S, 71°9′39.1″ W, 4700 masl | Alpaca | 12 | 6 | 1 | 0 | 4 (2 CoV + MRV; 1 CoV + RVA; 1 MRV + RVA) | 11 (91.7) |
| Total | 68 | 23 | 3 | 0 | 38 | 64 (94.1) | ||
| Silly/Marangani | 14°21′12″ S, 71°10′17″ W, 3800 masl ± | Llama | 12 | 6 | 1 | 0 | 5 (CoV + MRV) | 12 (100) |
| Total | 80 | 29 | 4 | 0 | 43 | 76 (95) | ||
| Host Species | No of Samples | Genus/Subgenus | ||
|---|---|---|---|---|
| BetaCoV/EmbeCoV | BetaCoV/ Not Identified | Genus/Subgenus Not Identified | ||
| Alpacas | 59 | 15 | 40 | 4 |
| Llamas | 11 | 1 | 10 | 0 |
| Total | 70 | 16 (22.9%) | 50 (71.4%) | 4 (5.7%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castilla, D.; Escobar, V.; Ynga, S.; Llanco, L.; Manchego, A.; Lázaro, C.; Navarro, D.; Santos, N.; Rojas, M. Enteric Viral Infections among Domesticated South American Camelids: First Detection of Mammalian Orthoreovirus in Camelids. Animals 2021, 11, 1455. https://doi.org/10.3390/ani11051455
Castilla D, Escobar V, Ynga S, Llanco L, Manchego A, Lázaro C, Navarro D, Santos N, Rojas M. Enteric Viral Infections among Domesticated South American Camelids: First Detection of Mammalian Orthoreovirus in Camelids. Animals. 2021; 11(5):1455. https://doi.org/10.3390/ani11051455
Chicago/Turabian StyleCastilla, Dayana, Victor Escobar, Sergio Ynga, Luis Llanco, Alberto Manchego, César Lázaro, Dennis Navarro, Norma Santos, and Miguel Rojas. 2021. "Enteric Viral Infections among Domesticated South American Camelids: First Detection of Mammalian Orthoreovirus in Camelids" Animals 11, no. 5: 1455. https://doi.org/10.3390/ani11051455
APA StyleCastilla, D., Escobar, V., Ynga, S., Llanco, L., Manchego, A., Lázaro, C., Navarro, D., Santos, N., & Rojas, M. (2021). Enteric Viral Infections among Domesticated South American Camelids: First Detection of Mammalian Orthoreovirus in Camelids. Animals, 11(5), 1455. https://doi.org/10.3390/ani11051455