
Cognitive and Composite Behavioural Welfare Assessments of Pet Cats between the Ages of 9–22 Months, Living in Single and Multi-Cat Households


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
- Investigate whether measures of relative judgment bias co-vary with CSSs. The CSS is believed to measure discrete behavioural responses to recent stress exposure, whereas judgment bias varies with underlying (and hence potentially longer term) affective states [32]. Hence, we would not expect to find a direct correlation between these measures.
- Assess the welfare of pet cats based on the presence of, and relationships with, other cats in the household, using the JBT and CSS.
- Cats in multi-cat households were hypothesized to show a longer latency to one or more ambiguous probes in a JBT (i.e., show a more ‘pessimistic’ judgment bias) and have higher CSSs compared to those living in single-cat households.
- Cats from multi-cat households where the focal cat was reported to show and/or be the recipient of agonistic behaviour were hypothesized to show a longer latency to one or more ambiguous probes in a JBT and higher CSS compared to those who did not exhibit/were not the recipients of agonistic behaviour.
2. Materials and Methods
2.1. Study Population
2.2. Experimental Protocol
2.2.1. Habituation
2.2.2. Verbal Questionnaire (Multi-Cat Households Only)
- Fluffing: ‘fluffing up’ at one another (when the hairs go on end making the cat look bigger).
- Freezing: freezing and staring at one another—for example, in corridors or doorways, for five seconds or more at a time.
- Vocalising: hissing/spitting/yowling/growling at one another.
- Blocking: blocking or inhibiting one another’s movements or being reluctant to pass one another in tight spaces (e.g., corridors and doorways).
- Aggression: fighting (aggression not occurring out of play), e.g., scratching/biting, chasing and attacking.
2.2.3. CSS
2.2.4. Feline Temperament Profiling
2.2.5. JBT
Training Phase
Test Phase
2.3. Statistical Analysis
2.3.1. CSS Preliminary Analysis
2.3.2. JBT Analysis
2.3.3. Comparing Welfare Measures Based on Social Conditions
3. Results
3.1. CSS
3.2. JBT
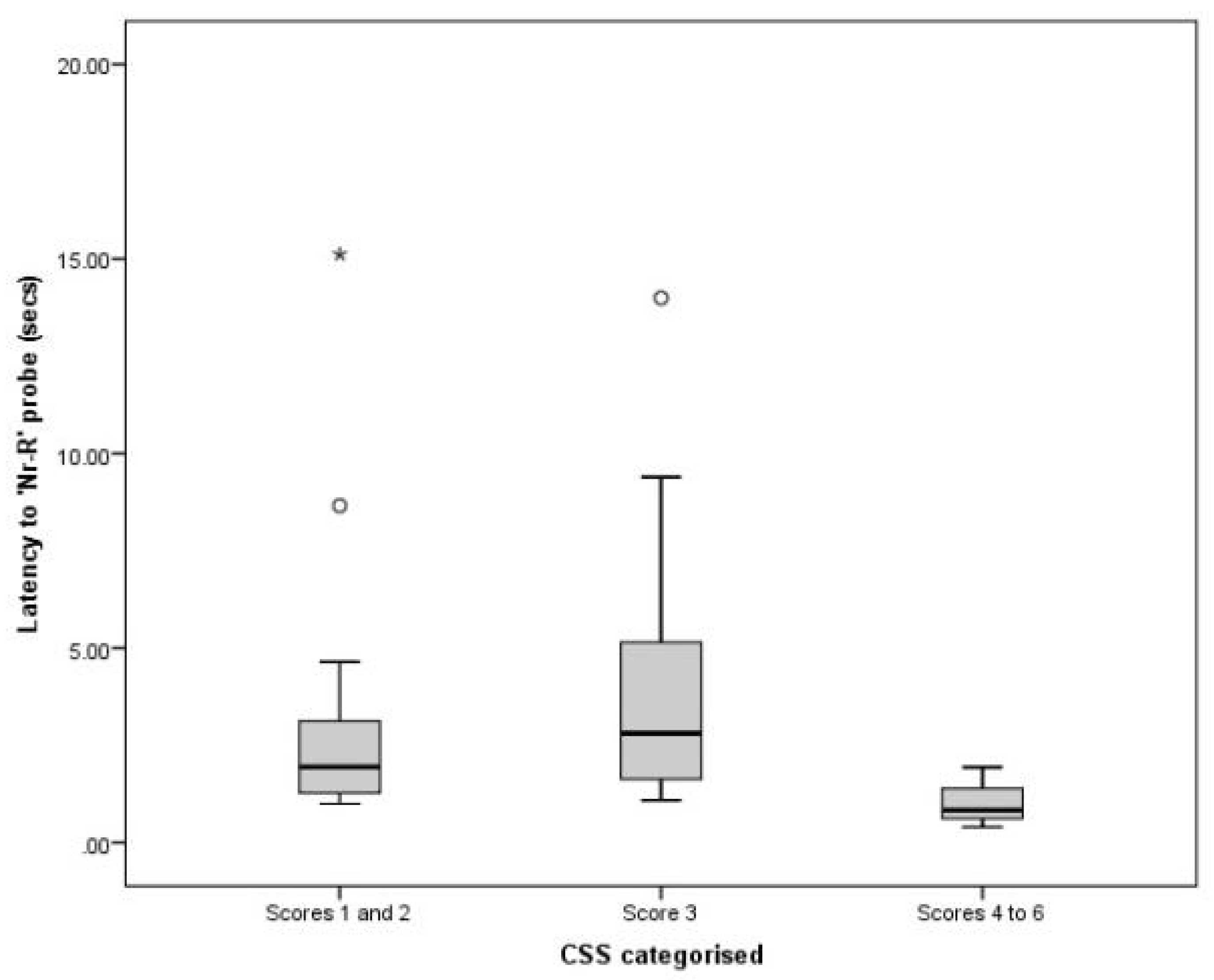
3.3. Comparing CSS and JBT Data
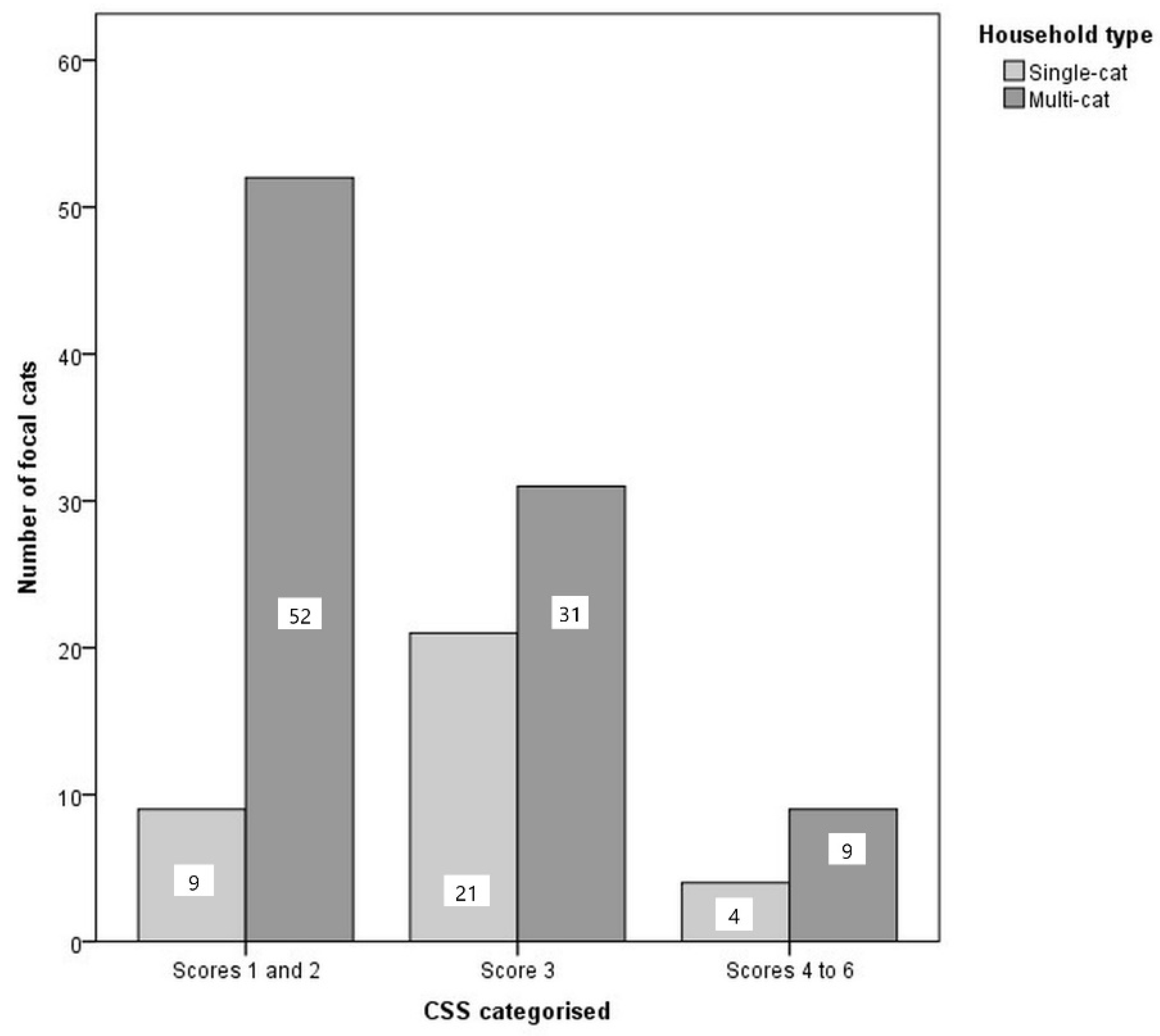
3.4. Comparing Welfare Measures between Multi-Cat and Single-Cat Households
3.5. Comparing Welfare Measures Based on the Occurrence of Agonistic Interactions in Multi-Cat Households
4. Discussion
4.1. Usability of CSS and JBT Welfare Measures in Home Environments
4.2. Comparing CSS and JBT Results
4.3. Comparing Groups of Cats Using JBT and CSS Welfare Measures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, J.K.; Browne, W.J.; Roberts, M.A.; Whitmarsh, A.; Gruffydd-Jones, T.J. Number and ownership profiles of cats and dogs in the UK. Vet. Rec. 2010, 166, 163–168. [Google Scholar] [CrossRef]
- Ramos, D. Common feline problem behaviors: Aggression in multi-cat households. J. Feline Med. Surg. 2019, 21, 221–233. [Google Scholar] [CrossRef]
- PDSA PET Animal Wellbeing (Paw) Report. Available online: https://www.pdsa.org.uk/media/7420/2019-paw-report_downloadable.pdf (accessed on 18 October 2020).
- Driscoll, C.A.; Menotti-Raymond, M.; Roca, A.L.; Hupe, K.; Johnson, W.E.; Geffen, E.; Harley, E.H.; Delibes, M.; Pontier, D.; Kitchener, A.C.; et al. The near eastern origin of cat domestication. Science 2007, 317, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Kleiman, D.G.; Eisenberg, J.F. Comparisons of canid and felid social systems from an evolutionary perspective. Anim. Behav. 1973, 21, 637–659. [Google Scholar] [CrossRef]
- Crowell-Davis, S.L.; Curtis, T.M.; Knowles, R.J. Social organization in the cat: A modern understanding. J. Feline Med. Surg. 2004, 6, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.W.S. Sociality in cats: A comparative review. J. Vet. Behav. 2016, 11, 113–124. [Google Scholar] [CrossRef]
- Bernstein, P.L.; Strack, M. A game of cat and house: Spatial patterns and behavior of 14 domestic cats (Felis catus) in the home. Anthrozoös 1996, 9, 25–39. [Google Scholar] [CrossRef]
- Pryor, P.A.; Hart, B.L.; Bain, M.J.; Cliff, K.D. Causes of urine marking in cats and effects of environmental management on frequency of marking. J. Am. Vet. Med. Assoc. 2001, 219, 1709–1713. [Google Scholar] [CrossRef] [PubMed]
- Casey, R.A.; Vandenbussche, S.; Bradshaw, J.W.S.; Roberts, M.A. Reasons for relinquishment and return of domestic cats (Felis silvestris catus) to rescue shelters in the UK. Anthrozoos 2009, 22, 347–358. [Google Scholar] [CrossRef]
- Vojtkovská, V.; Voslářová, E.; Večerek, V. Methods of assessment of the welfare of shelter cats: A review. Animals 2020, 10, 1527. [Google Scholar] [CrossRef]
- Lichtsteiner, M.; Turner, D.C. Influence of indoor-cat group size and dominance rank on urinary cortisol levels. Anim. Welf. 2008, 17, 215–237. [Google Scholar]
- Ramos, D.; Arena, M.N.; Reche-Junior, A.; Daniel, A.G.T.; Albino, M.V.C.; Vasconcellos, A.S.; Viau, P.; Oliveira, C.A. factors affecting faecal glucocorticoid levels in domestic cats (Felis catus): A pilot study with single and large multi-cat households. Anim. Welf. 2012, 21, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Ramos, D.; Reche-Junior, A.; Fragoso, P.L.; Palme, R.; Yanasse, N.K.; Gouvêa, V.R.; Beck, A.; Mills, D.S. Are cats (Felis catus) from multi-cat households more stressed? Evidence from assessment of fecal glucocorticoid metabolite analysis. Physiol. Behav. 2013, 122, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Broadley, H.M.; McCobb, E.C.; Slater, M.R. Effect of single-cat versus multi-cat home history on perceived behavioral stress in domestic cats (Felis silvestrus catus) in an animal shelter. J. Feline Med. Surg. 2014, 16, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Gourkow, N.; Fraser, D. The effect of housing and handling practices on the welfare, behavior, and selection of domestic cats (Felis sylvestris catus) by adopters in an animal shelter. Anim. Welf. 2006, 15, 371–377. [Google Scholar]
- Kessler, M.R.; Turner, D.C. Socialization and stress in cats (Felis silvestris catus) housed singly and in groups in animal shelters. Anim. Welf. 1999, 8, 15–26. [Google Scholar]
- Kessler, M.R.; Turner, D.C. Effects of density and cage size on stress in domestic cats (Felis silvestris catus) housed in animal shelters and boarding catteries. Anim. Welf. 1999, 8, 259–267. [Google Scholar]
- Ottway, D.S.; Hawkins, D.M. Cat housing in rescue shelters: A welfare comparison between communal and discrete-unit housing. Anim. Welf. 2003, 12, 173–189. [Google Scholar]
- Loberg, J.M.; Lundmark, F. The effect of space on behaviour in large groups of domestic cats kept indoors. Appl. Anim. Behav. Sci. 2016, 182, 23–29. [Google Scholar] [CrossRef]
- Roberts, C.; Gruffydd-Jones, T.; Williams, J.L.; Murray, J.K. Influence of living in a multicat household on health and behaviour in a cohort of cats from the united kingdom. Vet. Rec. 2020, 187, 27. [Google Scholar] [CrossRef] [Green Version]
- Finka, L.R.; Ellis, S.L.; Stavisky, J. A Critically Appraised Topic (CAT) to compare the effects of single and multi-cat housing on physiological and behavioural measures of stress in domestic cats in confined environments. BMC Vet. Res. 2014, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Bateson, M.; Matheson, S.M. Performance on a categorisation task suggests that removal of environmental enrichment induces “pessimism” in captive European starlings (Sturnus vulgaris). Anim. Welf. 2007, 16, 33–36. [Google Scholar]
- Mendl, M.; Burman, O.H.P.; Parker, R.M.A.; Paul, E.S. Cognitive bias as an indicator of animal emotion and welfare: Emerging evidence and underlying mechanisms. Appl. Anim. Behav. Sci. 2009, 118, 161–181. [Google Scholar] [CrossRef]
- Kasbaoui, N.; Cooper, J.; Mills, D.S.; Burman, O. Effects of long-term exposure to an electronic containment system on the behaviour and welfare of domestic cats. PLoS ONE 2016, 11, e0162073. [Google Scholar] [CrossRef] [Green Version]
- Papciak, J.; Popik, P.; Fuchs, E.; Rygula, R. Chronic psychosocial stress makes rats more “pessimistic” in the ambiguous-cue interpretation paradigm. Behav. Brain Res. 2013, 256, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.; Brooks, J.; Basse, C.; Burman, O.; Paul, E.; Blackwell, E.; Casey, R. Dogs showing separation-related behaviour exhibit a “pessimistic” cognitive bias. Curr. Biol. 2010, 20, R839–R840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tami, G.; Torre, C.; Compagnucci, M.; Manteca, X. Interpretation of ambiguous spatial stimuli in cats. Anim. Welf. 2011, 20, 185–189. [Google Scholar]
- Bartholomeus, M.; van der Borg, J.A.M.; Beerda, B.; Casey, R.A. Is the Judgment Bias Test Useful for Measuring Emotions in Privately-Owned Domestic Cats (Felis silvestris catus)? Minor Thesis, Wageningen University, Wageningen, The Netherlands, 2012. [Google Scholar]
- Kessler, M.R.; Turner, D.C. Stress and adaptation of cats (felis silvestris catus) housed singly, in pairs and in groups in boarding catteries. Anim. Welf. 1997, 6, 243–254. [Google Scholar]
- Casey, R.A.; Hawkins, K.; Bradshaw, J.W.S. Refinement and validation of the Kessler/Turner/McCune cat-stress-score. In Proceedings of the 40th International Congress of the ISAE, Bristol, UK, 8–12 August 2006. [Google Scholar]
- Mendl, M.; Burman, O.H.P.; Paul, E.S. An integrative and functional framework for the study of animal emotion and mood. Proc. Biol. Sci. 2010, 277, 2895–2904. [Google Scholar] [CrossRef] [Green Version]
- Research Randomiser. Available online: https://www.randomizer.org/ (accessed on 20 September 2014).
- Van den Bos, R.; de Vries, H. Clusters in social behaviour of female domestic cats (Felis Silvestris Catus) living in confinement. J. Ethol. 1996, 14, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Siegford, J.M.; Walshaw, S.O.; Brunner, P.; Zanella, A.J. Validation of a temperament test for domestic cats. Anthrozoos 2003, 16, 332–351. [Google Scholar] [CrossRef]
- StatsToDo, Cohen’s Kappa from Rating Scores. Available online: https://www.statstodo.com/Kappa_Pgm.php (accessed on 20 November 2014).
- McCobb, E.C.; Patronek, G.J.; Marder, A.; Dinnage, J.D.; Stone, M.S. Assessment of stress levels among cats in four animal shelters. J. Am. Vet. Med. Assoc. 2005, 226, 548–555. [Google Scholar] [CrossRef]
- Fureix, C.; Meagher, R.K. What can inactivity (in its various forms) reveal about affective states in non-human animals? A review. Appl. Anim. Behav. Sci. 2015, 171, 8–24. [Google Scholar] [CrossRef] [Green Version]
- Shreve, K.R.V.; Mehrkam, L.R.; Udell, M.A. Social interaction, food, scent or toys? A formal assessment of domestic pet and shelter cat (Felis silvestris catus) preferences. Behav. Process. 2017, 141, 322–328. [Google Scholar] [CrossRef]
- Doyle, R.E.; Fisher, A.D.; Hinch, G.N.; Boissy, A.; Lee, C. Release from restraint generates a positive judgment bias in sheep. Appl. Anim. Behav. Sci. 2010, 122, 28–34. [Google Scholar] [CrossRef]
- Vögeli, S.; Lutz, J.; Wolf, M.; Wechsler, B.; Gygax, L. Valence of physical stimuli, not housing conditions, affects behaviour and frontal cortical brain activity in sheep. Behav. Brain Res. 2014, 267, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Burman, O.H.P.; Parker, R.M.A.; Paul, E.S.; Mendl, M. A spatial judgment task to determine background emotional state in laboratory rats, Rattus norvegicus. Anim. Behav. 2008, 76I3, 801–809. [Google Scholar] [CrossRef] [Green Version]
- Finkler, H.; Terkel, J. Cortisol levels and aggression in neutered and intact free-roaming female cats living in urban social groups. Physiol. Behav. 2010, 99, 343–347. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Number of Cats in Household | Total Study Population | Study Population with Cat Stress Score Data | Study Population with Useable Judgment Bias Data | |||
|---|---|---|---|---|---|---|
| Number (%) of Households (Total = 105) | Number (%) of Focal Cats (Total = 128) | Number (%) of Households (Total = 103) | Number (%) of Focal Cats (Total = 126) | Number (%) of Households (Total = 37) | Number (%) of Focal Cats (Total = 42) | |
| 1 | 34 (32.4) | 34 (26.6) | 34 (33.0) | 34 (27.0) | 9 (24.3) | 9 (21.4) |
| 2 | 47 (44.8) | 61 (47.7) | 45 (43.7) | 59 (46.8) | 22 (59.5) | 26 (61.9) |
| 3 | 7 (6.7) | 10 (7.8) | 7 (6.8) | 10 (7.9) | 3 (8.1) | 4 (9.5) |
| 4 | 6 (5.7) | 9 (7.0) | 6 (5.8) | 9 (7.1) | 1 (2.7) | 1 (2.4) |
| 5 | 5 (4.8) | 6 (4.7) | 5 (4.9) | 6 (4.8) | 1 (2.7) | 1 (2.4) |
| 6 | 2 (1.9) | 2 (1.6) | 2 (1.9) | 2 (1.6) | 0 (0.0) | 0 (0.0) |
| 7 | 1 (1.0) | 1 (0.8) | 1 (1.0) | 1 (0.8) | 0 (0.0) | 0 (0.0) |
| 8 | 3 (2.9) | 5 (3.9) | 3 (2.9) | 5 (4.0) | 1 (2.7) | 1 (2.4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozgunay, S.; Murray, J.K.; Rowe, E.; Gee, N.R.; Bartholomeus, M.; Casey, R. Cognitive and Composite Behavioural Welfare Assessments of Pet Cats between the Ages of 9–22 Months, Living in Single and Multi-Cat Households. Animals 2021, 11, 1793. https://doi.org/10.3390/ani11061793
Ozgunay S, Murray JK, Rowe E, Gee NR, Bartholomeus M, Casey R. Cognitive and Composite Behavioural Welfare Assessments of Pet Cats between the Ages of 9–22 Months, Living in Single and Multi-Cat Households. Animals. 2021; 11(6):1793. https://doi.org/10.3390/ani11061793
Chicago/Turabian StyleOzgunay, Sezan, Jane K. Murray, Elizabeth Rowe, Nancy R. Gee, Marije Bartholomeus, and Rachel Casey. 2021. "Cognitive and Composite Behavioural Welfare Assessments of Pet Cats between the Ages of 9–22 Months, Living in Single and Multi-Cat Households" Animals 11, no. 6: 1793. https://doi.org/10.3390/ani11061793
APA StyleOzgunay, S., Murray, J. K., Rowe, E., Gee, N. R., Bartholomeus, M., & Casey, R. (2021). Cognitive and Composite Behavioural Welfare Assessments of Pet Cats between the Ages of 9–22 Months, Living in Single and Multi-Cat Households. Animals, 11(6), 1793. https://doi.org/10.3390/ani11061793