
Effects of Dietary Saccharomyces cerevisiae YFI-SC2 on the Growth Performance, Intestinal Morphology, Immune Parameters, Intestinal Microbiota, and Disease Resistance of Crayfish (Procambarus clarkia)

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Yeast Strains and Growth Conditions
2.3. Feeding Frequency
2.4. Growth for Performance and Sample Collection
2.4.1. Growth Performance
2.4.2. Sample Collection
2.5. Histopathology Analysis
2.6. Real-Time PCR Analysis of Immune-Related Genes
2.7. Genomic DNA Extraction and 16S rRNA Gene Sequencing
2.8. Challenge Test
2.9. Statistical and Correlation Analyses
3. Results
3.1. Growth Performance
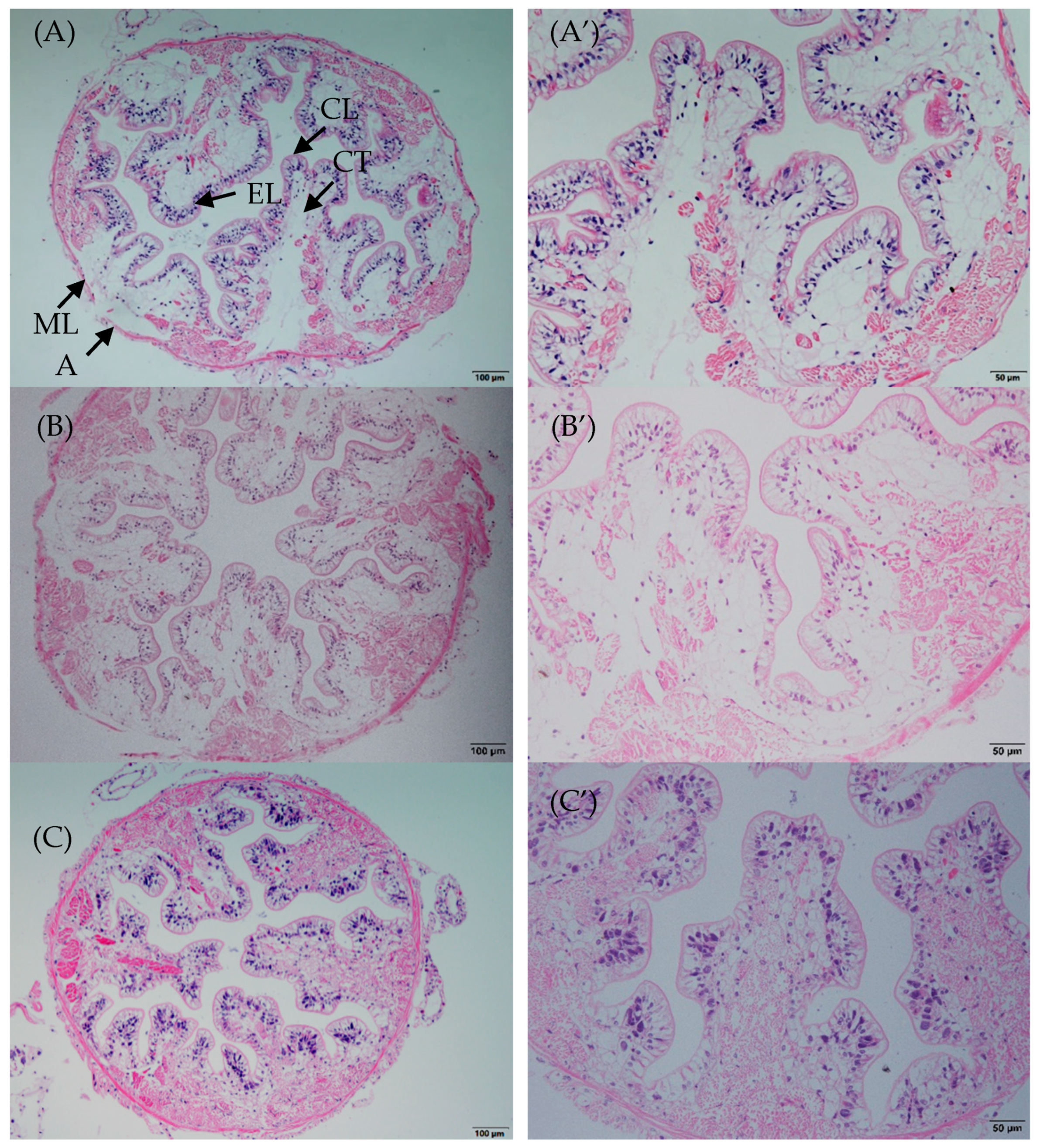
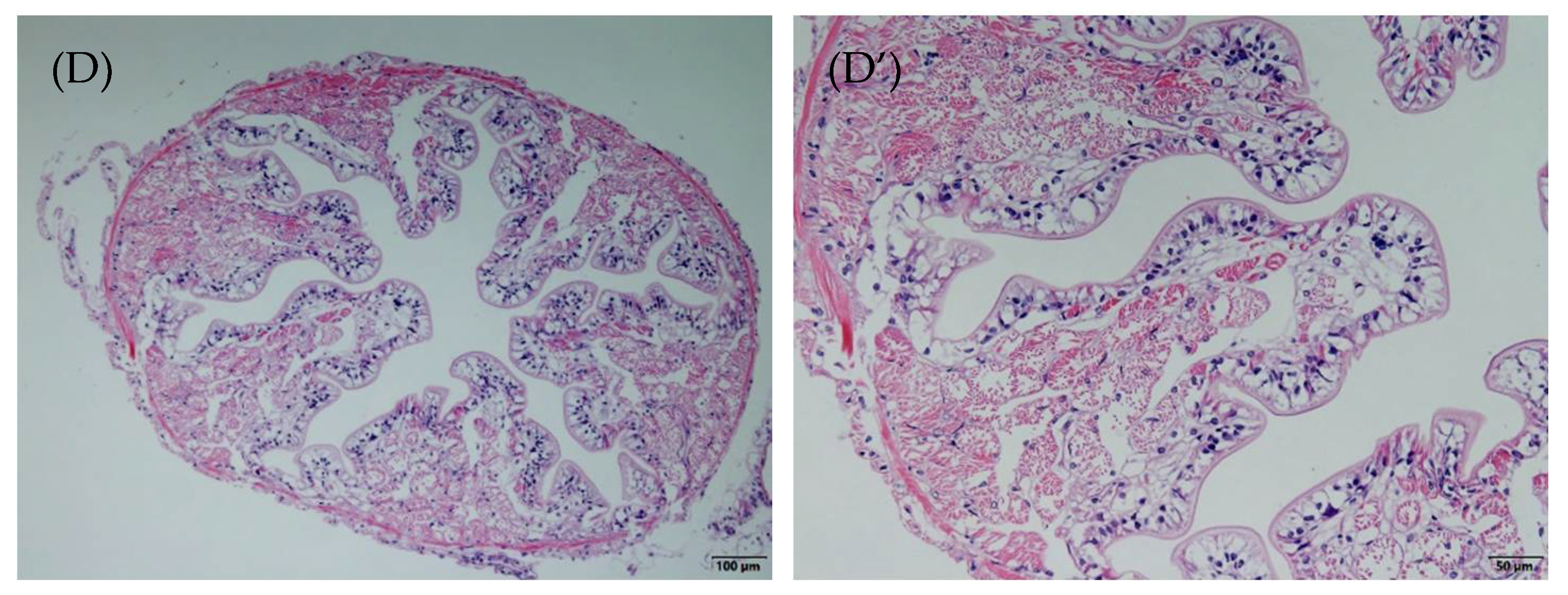
3.2. Intestinal Morphology Changes
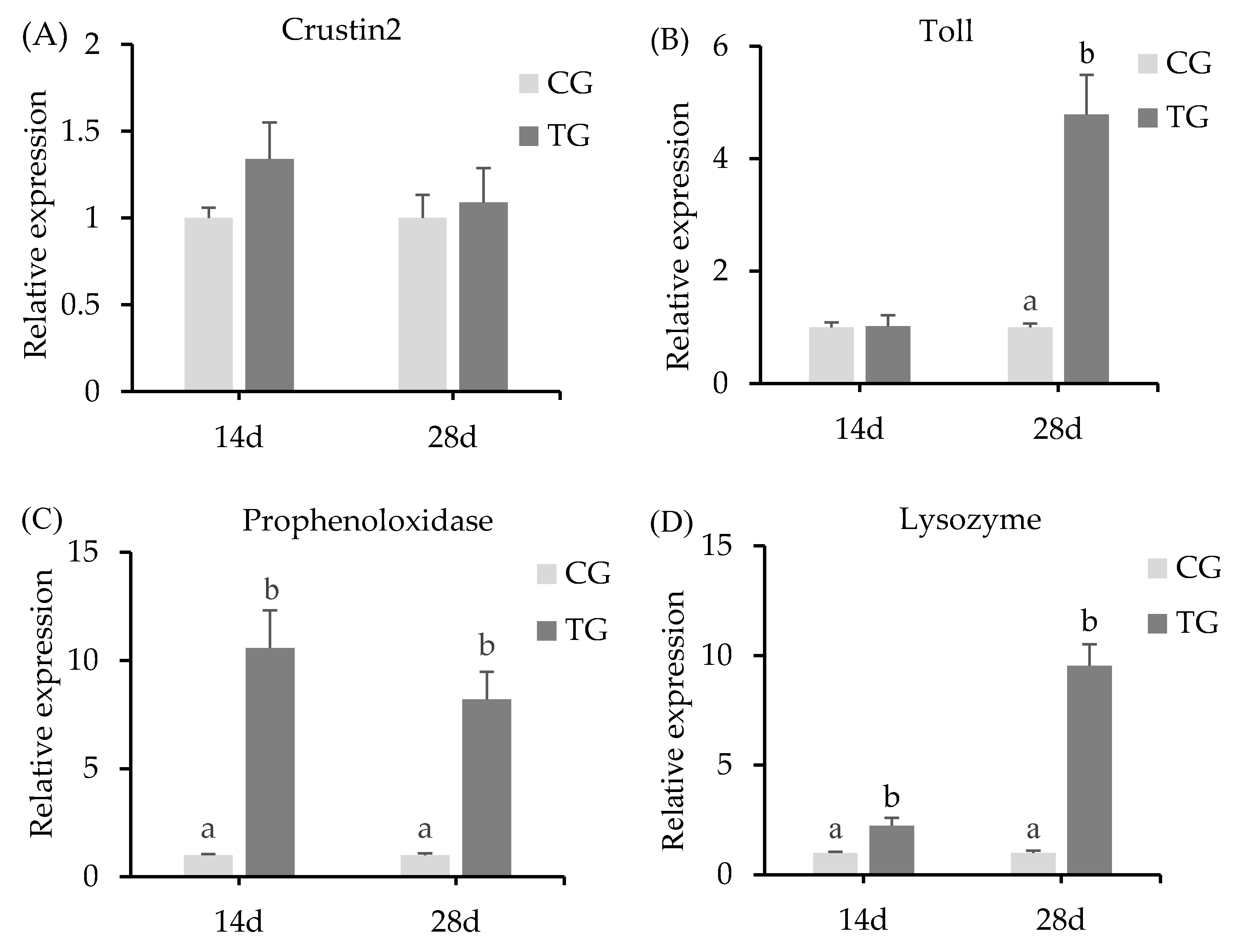
3.3. Expression Levels of Immunity-Related Genes in the Hepatopancreas
3.4. Richness and Diversity of Intestinal Microbiota
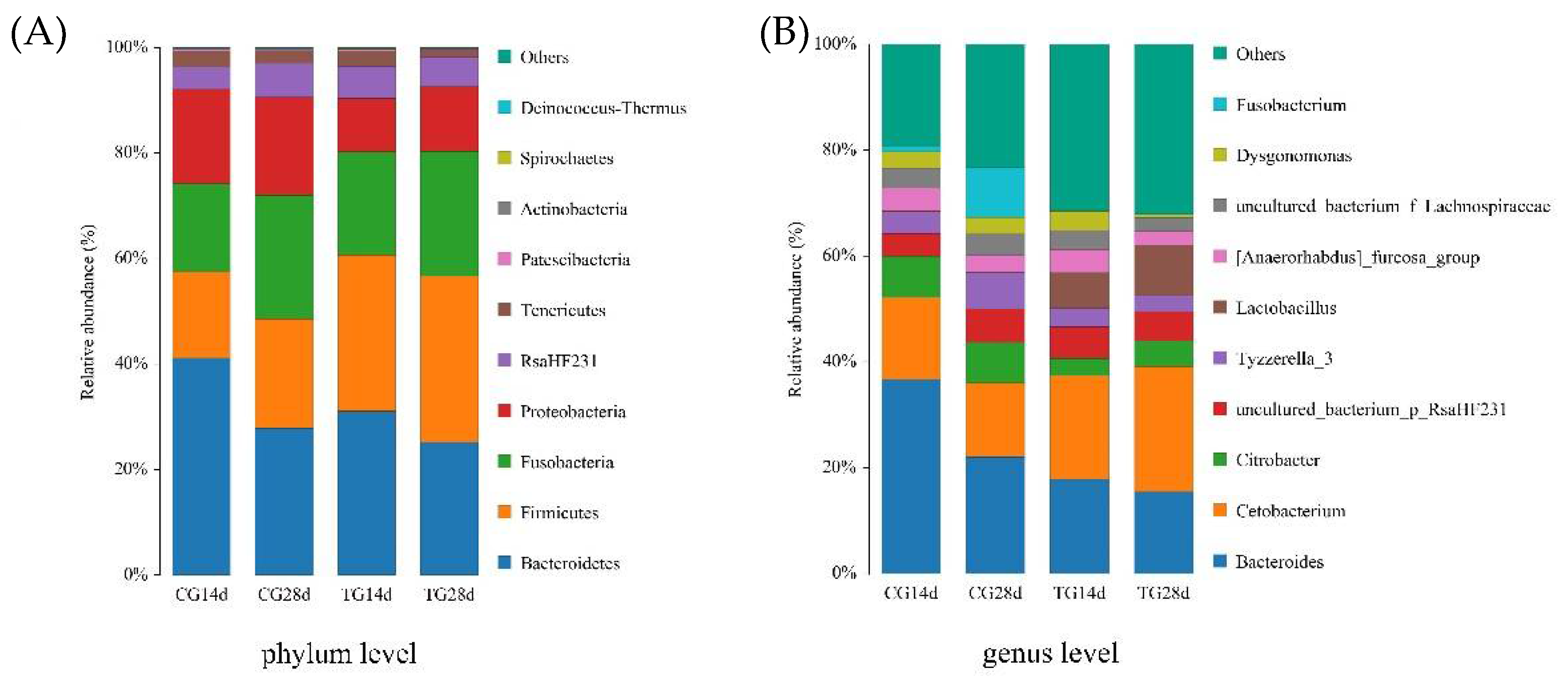
3.5. Phylum and Genus Performance
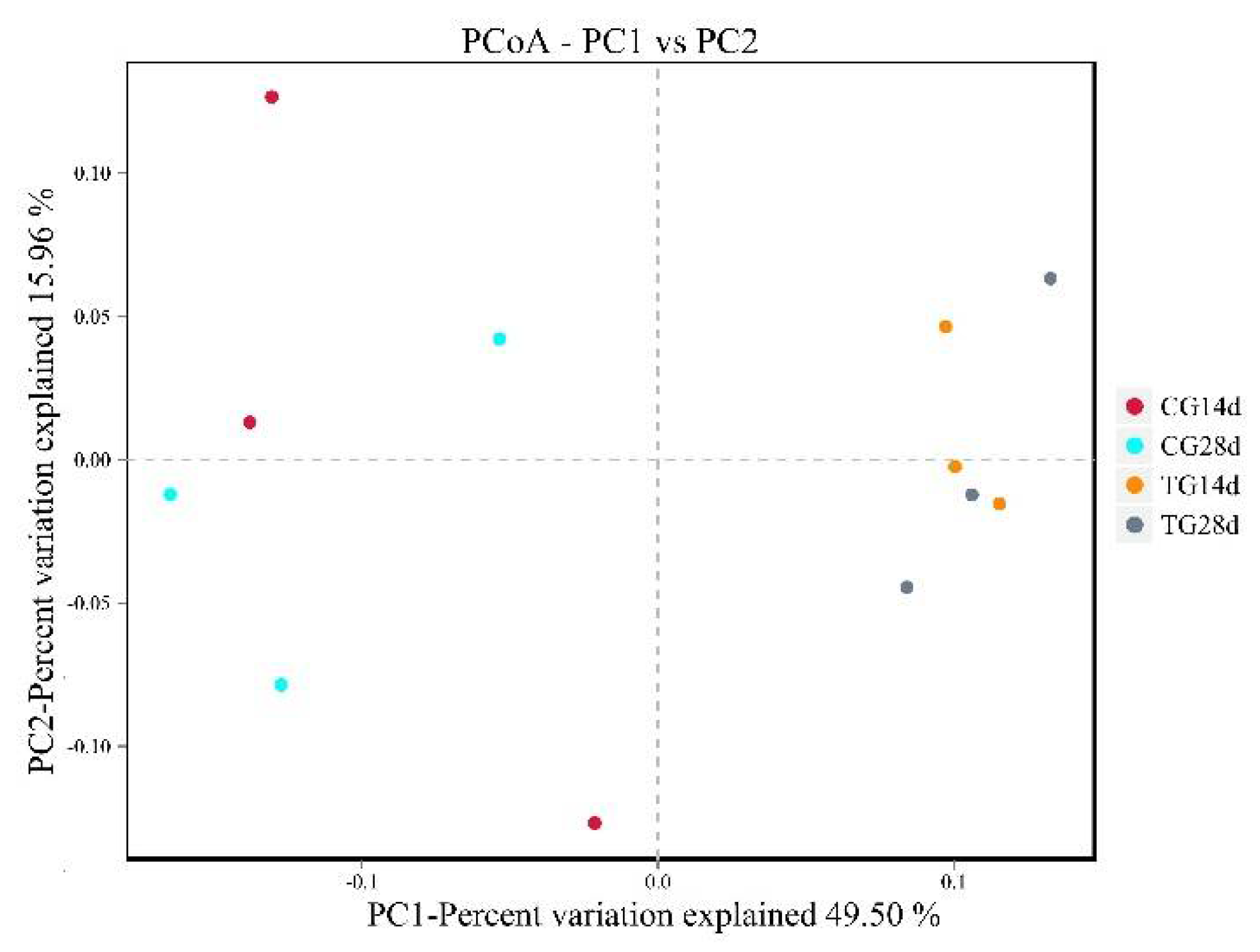
3.6. Principle Coordinate Analysis (PCoA)
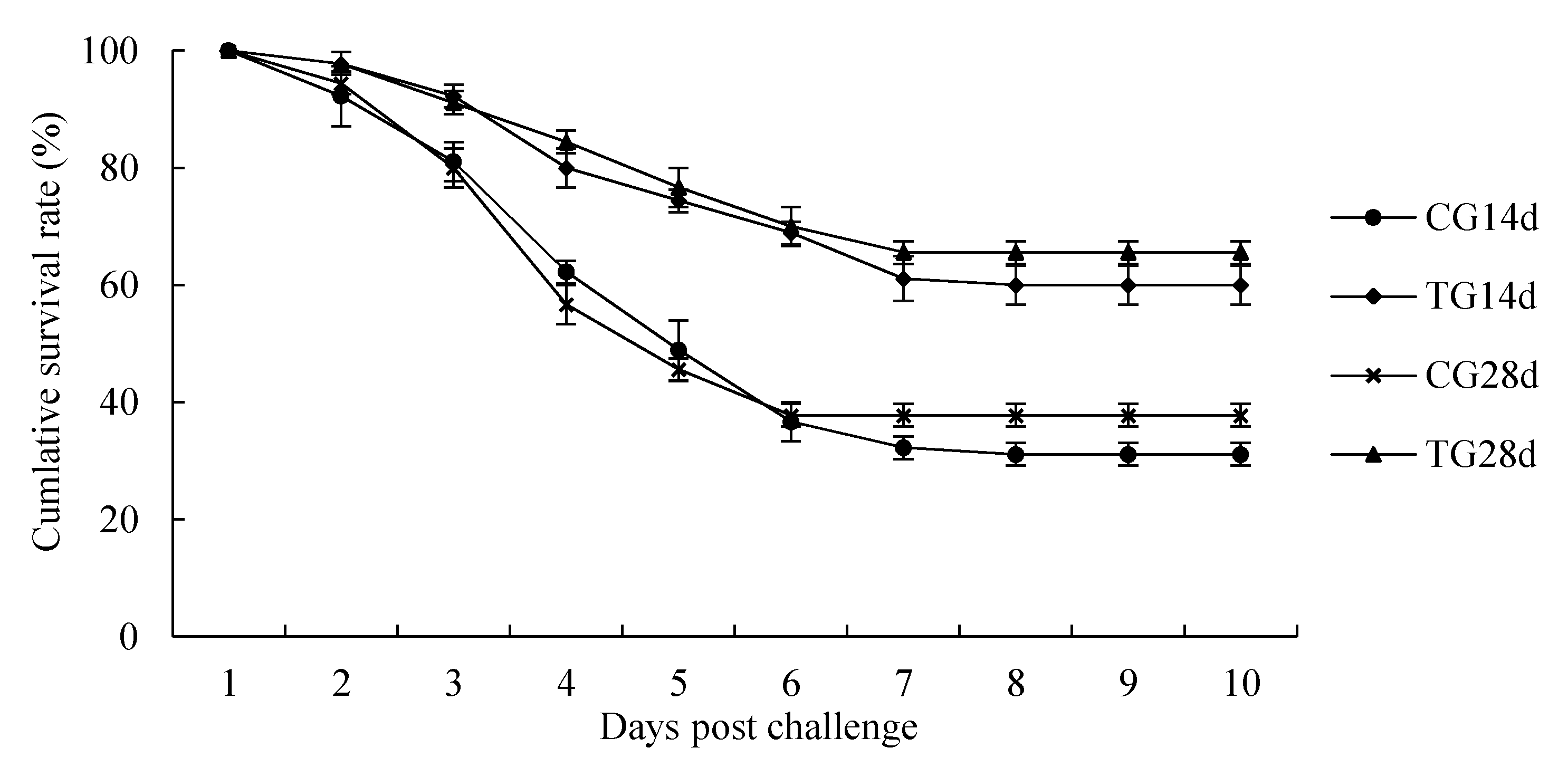
3.7. Challenge Test
4. Discussion
4.1. Growth Performance
4.2. Intestinal Histomorphometric
4.3. Immune Related Gene Expressions
4.4. Intestinal Microbiota Analysis
4.5. Disease Resistance Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harris, D.J.; Crandall, K.A. Intragenomic variation within ITS1 and ITS2 of freshwater crayfishes (Decapoda: Cambaridae): Implications for phylogenetic and microsatellite studies. Mol. Biol. Evol. 2000, 17, 284–291. [Google Scholar] [CrossRef]
- Xiong, B.; Xu, T.; Li, R.P.; Johnson, D.; Ren, D.; Liu, H.G.; Xi, Y.; Huang, Y.P. Heavy metal accumulation and health risk assessment of crayfish collected from cultivated and uncultivated ponds in the Middle Reach of Yangtze River. Sci. Total Environ. 2020, 739, 139963. [Google Scholar] [CrossRef]
- Liu, Q.; Long, Y.N.; Li, B.; Zhao, L.L.; Luo, X.; Xu, L.; Luo, W.; Du, Z.J.; Zhou, J.; Yang, S. Rice-shrimp culture: A better intestinal microbiota, immune enzymatic activities, and muscle relish of crayfish (Procambarus clarkii) in Sichuan Province. Appl. Microbiol. Biot. 2020, 104, 9413–9420. [Google Scholar] [CrossRef]
- The State of World Fisheries and Aquaculture; Sustainability in Action, Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; Volume 30.
- Fisheries and Fisheries Administration Bureau of Ministry of Agriculture and Industry. China Fisheries Yearbook; China Agriculture Press: Beijing, Chinese, 2020; Volume 26. [Google Scholar]
- Liu, X.D.; He, X.; An, Z.H.; Sun, W.; Chen, N.; Gao, X.J.; Li, X.X.; Zhang, X.J. Citrobacter freundii infection in red swamp crayfish (Procambarus clarkii) and host immune-related gene expression profiles. Aquaculture 2020, 515, 734499. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.Y.; Sergey, K.; Andrey, S.; Chen, C.; Feng, Y.J.; Ren, N.Q.; Sun, K. Effects of cadmium on intestinal histology and microbiota in freshwater crayfish (Procambarus clarkii). Chemosphere 2020, 242, 125105. [Google Scholar] [CrossRef] [PubMed]
- Joseph, E.B.; Karen, G.B.; Louis, E.B. Disease resistance of Pacific white shrimp, Litopenaeus vannamei, following the dietary administration of a yeast culture food supplement. Aquaculture 2003, 231, 1–8. [Google Scholar]
- Sruthy, K.S.; Nair, A.; Puthumana, J.; Antony, S.P.; Singh, I.B.; Philip, R. Molecular cloning, recombinant expression and functional characterization of an antimicrobial peptide, Crustin from the Indian white shrimp, Fenneropenaeus indicus. Fish Shellfish Immunol. 2017, 71, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Kenneth, S. Early events in crustacean innate immunity. Fish Shellfish Immunol. 2002, 12, 421–443. [Google Scholar]
- Li, F.H.; Xiang, J.H. Recent advances in researches on the innate immunity of shrimp in China. Dev. Comp. Immunol. 2013, 39, 11–26. [Google Scholar] [CrossRef]
- Liu, H.T.; Wang, J.; Mao, Y.; Liu, M.; Niu, S.F.; Qiao, Y.; Su, Y.Q.; Wang, C.Z.; Zheng, Z.P. Identification and expression analysis of a new invertebrate lysozyme in Kuruma shrimp (Marsupenaeus japonicus). Fish Shellfish Immunol. 2016, 49, 336–343. [Google Scholar] [CrossRef]
- Peera, H.; James, V. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenter. 2013, 6, 39–51. [Google Scholar]
- Seyed, H.H.; Einar, R.; Alireza, S.M.; Maria, Á.E. Probiotic, prebiotic and synbiotic supplements in sturgeon aquaculture: A review. Rev. Aquacult. 2016, 8, 89–102. [Google Scholar]
- Ortuño, J.; Cuesta, A.; Rodrıguez, A.; Esteban, M.; Meseguer, J. Oral administration of yeast, Saccharomyces cerevisiae enhances the cellular innate immune response of gilthead seabream (Sparus aurata L.). Vet. Immunol. Immunopathol. 2002, 85, 41–50. [Google Scholar] [CrossRef]
- Wang, S.J.; Guo, C.H.; Zhou, L.; Zhong, Z.D.; Zhu, W.Z.; Huang, Y.L.; Zhang, Z.F.; Gorgels, T.G.; Berendschot, T.T. Effects of dietary supplementation with epidermal growth factor expressing Saccharomyces cerevisiae on duodenal development in weaned piglets. Brit. J. Nutr. 2016, 115, 1509–1520. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.Y.; Yang, F.; Hu, J.P.; Han, D.; Liu, H.K.; Jin, J.Y.; Yang, Y.X.; Yi, J.H.; Zhu, X.M.; Xie, S.Q. Optimal form of yeast cell wall promotes growth, immunity and disease resistance in gibel carp (Carassius auratus gibelio). Aquac. Rep. 2020, 18, 100465. [Google Scholar] [CrossRef]
- Huang, L.; Ran, C.; He, S.X.; Ren, P.F.; Hu, J.; Zhao, Z.X.; Zhou, Z.J. Effects of dietary Saccharomyces cerevisiae culture or live cells with Bacillus amyloliquefaciens spores on growth performance, gut mucosal morphology, hsp70 gene expression, and disease resistance of juvenile common carp (Cyprinus carpio). Aquaculture 2015, 438, 33–38. [Google Scholar] [CrossRef]
- Dias, M.K.R.; Yoshioka, E.T.O.; Rodriguez, A.F.R.; Ribeiro, R.A.; Faria, F.S.E.D.V.; Ozório, R.O.A.; Tavares-Dias, M. Growth and hematological and immunological responses of Arapaima gigas fed diets supplemented with immunostimulant based on Saccharomyces cerevisiae and subjected to handling stress. Aquac. Rep. 2020, 17, 100335. [Google Scholar] [CrossRef]
- Mohammad, A.; Shapour, K.; Mehrdad, M. Immunity enhancement with administration of Gracilaria corticata and Saccharomyces cerevisiae compared to gamma irradiation in expose to WSSV in shrimp, in juvenile Litopenaeus vannamei: A comparative study. Fish Shellfish Immunol. 2016, 56, 21–33. [Google Scholar]
- Zhang, M.Z.; Pan, L.Q.; Fan, D.P.; He, J.J.; Su, C.; Gao, S.; Zhang, M.Y. Study of fermented feed by mixed strains and their effects on the survival, growth, digestive enzyme activity and intestinal flora of Penaeus vannamei. Aquaculture 2021, 530, 735703. [Google Scholar] [CrossRef]
- Fang, H.; Wang, B.J.; Jiang, K.Y.; Liu, M.; Wang, L. Effects of Lactobacillus pentosus HC-2 on the growth performance, intestinal morphology, immune-related genes and intestinal microbiota of Penaeus vannamei affected by aflatoxin B1. Aquaculture 2020, 525, 735289. [Google Scholar] [CrossRef]
- Shi, X.Z.; Zhang, R.R.; Jia, Y.P.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. Identification and molecular characterization of a Spätzle-like protein from Chinese shrimp (Fenneropenaeus chinensis). Fish Shellfish Immunol. 2009, 27, 610–617. [Google Scholar] [CrossRef]
- Li, Y.H.; Zheng, L.; Chen, H.Q.; Wang, H.Z.; Wang, L.Q.; Xu, D.P. Cloning and Sequence Analysis of Prophenoloxidase from Haemocytes of the Red Swamp Crayfish, Procambarus clarkii. Agric. Sci. China 2008, 8, 369–379. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Y.H.; Dai, Y.J.; Tan, J.M.; Huang, Y.; Lan, J.F.; Ren, Q. A novel vertebrates Toll-like receptor counterpart regulating the anti-microbial peptides expression in the freshwater crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2015, 43, 219–229. [Google Scholar] [CrossRef]
- Liu, B.; Zhou, Q.L.; Sun, C.X.; Song, C.Y.; Zhang, H.M.; Yang, Z.F.; Shan, F. Patterns of Bacterial Community Composition and Diversity Following the Embryonic Development Stages of Macrobrachium rosenbergii. Aquac. Rep. 2020, 17, 100372. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Grice, A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.; Nakhro, K.; Chowdhury, S.; Kamilya, D. Effects of potential probiotic Bacillus amyloliquefaciens FPTB16 on systemic and cutaneous mucosal immune responses and disease resistance of catla (Catla catla). Fish Shellfish Immunol. 2013, 35, 1547–1553. [Google Scholar] [CrossRef]
- Merrifield, D.; Bradley, G.; Baker, R.; Davies, S. Probiotic applications for rainbow trout (Oncorhynchus mykiss Walbaum) I. Effects on growth performance, feed utilization, intestinal microbiota and related health criteria. Aquac. Nutr. 2010, 16, 496–503. [Google Scholar] [CrossRef]
- Caridad, S.; Guevara, C.A. Probiotic Use of Yeast Saccharomyces Cerevisiae in Animal Feed. Res. J. Zool. 2018, 1. [Google Scholar] [CrossRef]
- Gupta, A.; Dhawan, A. Effect of dietary probiotic improval (Lactobacillus sporogenes and Saccharomyces cerevisiae) on growth and feed utilization of Macrobrachium rosenbergii post larvae. Anim. Nutr. Feed Technol. 2012, 12, 209–217. [Google Scholar]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; El Basuini, M.F.; Hossain, M.S.; Nhu, T.H.; Moss, A.S.; Dossou, S.; Wei, H. Dietary supplementation of β-glucan improves growth performance, the innate immune response and stress resistance of red sea bream, Pagrus major. Aquac. Nutr. 2017, 23, 148–159. [Google Scholar] [CrossRef]
- Lu, J.T.; Qi, C.G.; Samwel, M.L.; Han, F.L.; Yang, L.; Wang, X.D.; Jian, G.Q.; Chen, L.Q. Dietary mannan oligosaccharide (MOS) improves growth performance, antioxidant capacity, non-specific immunity and intestinal histology of juvenile Chinese mitten crabs (Eriocheir sinensis). Aquaculture 2019, 510, 337–346. [Google Scholar] [CrossRef]
- Waraporn, H.; Karun, T.; Naraid, S. Effects of dietary supplementation of oligosaccharides on growth performance, gut health and immune response of hybrid catfish (Pangasianodon gigas × Pangasianodon hypophthalmus). Aquaculture 2019, 507, 97–107. [Google Scholar]
- Lee, S.; Katya, K.; Park, Y.; Won, S.; Seong, M.; Hamidoghli, A.; Bai, S.C. Comparative evaluation of dietary probiotics Bacillus subtilis WB60 and Lactobacillus plantarum KCTC3928 on the growth performance, immunological parameters, gut morphology and disease resistance in Japanese eel, Anguilla japonica. Fish Shellfish Immunol. 2017, 61, 201–210. [Google Scholar] [CrossRef]
- Salah, M.A.; Yousef, A.A.; Ahlam, A.G.; Moahmed, F.M. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immunol. 2008, 25, 128–136. [Google Scholar]
- Magalhaes, J.G.; Tattoli, I.; Girardin, S.E. The intestinal epithelial barrier: How to distinguish between the microbial flora and pathogens. Semin. Immunol. 2007, 19, 106–115. [Google Scholar] [CrossRef]
- Burrells, C.; Williams, P.D.; Southgate, P.J. Dietary nucleotides: A novel supplement in fish feeds: 2. Effectson vaccination, salt water transfer, growth rates and physiology of Atlantic salmon (Salmo salar L). Aquaculture 2001, 199, 171–184. [Google Scholar] [CrossRef]
- Yao, W.X.; Li, X.Q.; Zhang, C.Y.; Wang, J.; Cai, Y.W.; Leng, X.J. Effects of dietary synbiotics supplementation methods on growth, intestinal health, non-specific immunity and disease resistance of Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2021, 112, 46–55. [Google Scholar] [CrossRef]
- Rebeca, C.; Milena, F.; Silvana, T.T.; José, M.; Miguel, Á.M.; Ángeles, E. Histological alterations and microbial ecology of the intestine in gilthead seabream (Sparus aurata L.) fed dietary probiotics and microalgae. Cell Tissue Res. 2012, 350, 477–489. [Google Scholar]
- Yang, X.Y.; He, Y.F.; Chi, S.Y.; Tan, B.P.; Lin, S.; Dong, X.H.; Yang, Q.H.; Liu, H.Y.; Zhang, S. Supplementation with Saccharomyces cerevisiae hydrolysate in a complex plant protein, low-fishmeal diet improves intestinal morphology, immune function and Vibrio harveyi disease resistance in Epinephelus coioides. Aquaculture 2020, 529, 735655. [Google Scholar] [CrossRef]
- Miao, S.Y.; Han, B.; Zhao, C.Z.; Hu, J.T.; Zhu, J.Y.; Zhang, X.; Sun, L.S. Effects of dietary Pediococcus acidilactici GY2 single or combined with Saccharomyces cerevisiae or/and β-glucan on the growth, innate immunity response and disease resistance of Macrobrachium rosenbergii. Fish Shellfish Immunol. 2020, 98, 68–76. [Google Scholar] [CrossRef]
- Alan, L.J.; Ángel, C.C.; Antonio, L.G.; Ilie, S.R.; Marlene, T.; Carlos, A. Dietary supplementation of marine yeast Yarrowia lipolytica modulates immune response in Litopenaeus vannamei. Fish Shellfish Immunol. 2020, 105, 469–476. [Google Scholar]
- Zhao, Y.C.; Ma, H.M.; Zhang, W.B.; Qing, H.; Ai, K.S.; Mai, W.; Xu, X.J.; Wang, Z.G.; Liu, F. Effects of dietary β-glucan on the growth, immune responses and resistance of sea cucumber, Apostichopus japonicus against Vibrio splendidus infection. Aquaculture 2011, 315, 269–274. [Google Scholar] [CrossRef]
- Biswas, G.; Korenaga, H.; Takayama, H.; Kono, T.; Shimokawa, H.; Sakai, M. Cytokine responses in the common carp, Cyprinus carpio L. treated with baker’s yeast extract. Aquaculture 2012, 356–357, 169–175. [Google Scholar] [CrossRef]
- Liang, Q.J.; Li, Z.H.; Ou, M.F.; Wu, X.J.; Qiao, X.L.; Wei, W.; Liu, Y.; Ye, J.M.; Wang, W.N. Hypoimmunity and intestinal bacterial imbalance are closely associated with blue body syndrome in cultured Penaeus vannamei. Aquaculture 2020, 522, 735118. [Google Scholar] [CrossRef]
- Zhang, M.L.; Sun, Y.H.; Liu, Y.H.; Qiao, F.; Chen, L.Q.; Liu, W.T.; Du, Z.Y.; Li, E. Response of gut microbiota to salinity change in two euryhaline aquatic animals with reverse salinity preference. Aquaculture 2016, 454, 72–80. [Google Scholar] [CrossRef]
- Emilie, C.; Yannick, G.; Kevin, M.; Bénédicte, L.; David, P.; Fabien, P.; Florian, N.; Denis, S. Bacterial community characterization of water and intestine of the shrimp Litopenaeus stylirostris in a biofloc system. BMC Microbiol. 2016, 16, 157–165. [Google Scholar]
- Oscar, C.; Thompson, C.; José, M.; Angel, G. Colonization and Impact of Disease and Other Factors on Intestinal Microbiota. Digest. Dis. Sci. 2007, 52, 2069–2077. [Google Scholar]
- Daniel, L.M.; Dean, B.; Graham, B.; Simon, J.D.; Remi, T.M. Microbial community diversity associated with the intestinal mucosa of farmed rainbow trout (Oncoryhnchus mykiss Walbaum). Aquac. Res. 2009, 40, 1064–1072. [Google Scholar]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Miao, S.Y.; Zhao, C.Z.; Zhu, J.Y.; Hu, J.T.; Dong, X.J.; Sun, L.S. Dietary soybean meal affects intestinal homoeostasis by altering the microbiota, morphology and inflammatory cytokine gene expression in northern snakehead. Sci. Rep. 2018, 8, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Li, X.D.; Zhang, R.Y.; Zhao, Y.Y.; Wu, G.F.; Liu, J.L.; Zhu, X.C.; Li, L. Comparative analysis of the intestinal bacterial community and expression of gut immunity genes in the Chinese Mitten Crab (Eriocheir sinensis). AMB Express 2018, 8, 192. [Google Scholar] [CrossRef]
- Wang, A.R.; Ran, C.; Einar, R.; Zhou, Z.G. Progress in fish gastrointestinal microbiota research. Rev. Aquacult. 2018, 10, 626–640. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, C.; Sakata, T.; Sugita, H. Novel ecological niche of Cetobacterium somerae, an anaerobic bacterium in the intestinal tracts of freshwater fish. Lett. Appl. Microbiol. 2008, 46, 43–48. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, P.J.; Shang, X.C.; Lu, Y.T.; Li, Y.H. Exposure of lead on intestinal structural integrity and the diversity of gut microbiota of common carp. Comp. Biochem. Physiol. Part C 2021, 239, 108877. [Google Scholar] [CrossRef]
- Stephen, A.; Shen, J.F.; Tan, B.P.; Dong, X.H.; Liu, H.Y. Effects of dietary yeast culture on shrimp growth, immune response, intestinal health and disease resistance against Vibrio harveyi. Fish Shellfish Immunol. 2020, 102, 286–295. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′-3′) | GenBank Number |
|---|---|---|
| For qPCR | ||
| 18S-F | TGGTGCATGGCCGTTCTTA | MT829236.1 |
| 18S-R | AATTGCTGGAGATCCGTCGAC | |
| Crustin2-F | GGGAAGAAAAGCACAATGGT | GQ301202.1 [23] |
| Crustin2-R | GGTATGGAGGTCGAGACAGG | |
| Prophenoloxidase-F | AGGTGGATCAGCCAGCAGT | EF595973.1 [24] |
| Prophenoloxidase-R | CGTAGTCAGCAGCGGAGGT | |
| Lysozyme-F | GATTGCTTAGGGTGCTTGTGCGA | GQ301200.1 [23] |
| Lysozyme-R | GGGTTTGCCAGCTTCATTCCAGT | |
| Toll-like receptor-F | GACTTGTCCAAAAACGATATACG | KP259728.1 [25] |
| Toll-like receptor-R | TGCGTTACAGTAGTGAGCGAAT | |
| For V3—V4 regions of 16S rRNA gene | ||
| 338F | ACTCCTACGGGAGGCAGCA | |
| 806R | GGACTACHVGGGTWTCTAAT | |
| Parameters | Dietary Treatments | |||
|---|---|---|---|---|
| CG14d | TG14d | CG28d | TG28d | |
| Initial weight (g) | 13.72 ± 0.18 | 13.69 ± 0.21 | 13.72 ± 0.18 | 13.69 ± 0.21 |
| Weight at sampling (g) | 15.13 ± 0.32 | 15.45 ± 0.39 | 16.67 ± 0.38 a | 17.43 ± 0.37 b |
| WGR (%) | 10.13 ± 0.43 | 12.86 ± 0.57 | 21.61 ± 1.03 a | 27.09 ± 1.87 b |
| SGR (%/d) | 0.69 ± 0.04 | 0.81 ± 0.13 | 0.72 ± 0.05 a | 0.86 ± 0.07 b |
| Survival rate (%) | 95.83 ± 1.44 | 96.25 ± 1.25 | 92.08 ± 0.72 | 93.75 ± 1.25 |
| Sample ID | OTUs | Chao 1 | Shannon |
|---|---|---|---|
| CG14d | 170 ± 28 a | 207.08 ± 15.89 a | 2.33 ± 0.27 a |
| TG14d | 241 ± 31 b | 258.97 ± 28.48 b | 2.97 ± 0.32 b |
| CG28d | 171 ± 17 a | 201.04 ± 9.75 a | 2.42 ± 0.15 a |
| TG28d | 228 ± 23 ab | 250.16 ± 16.48 b | 2.85 ± 0.39 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Li, Y.; Xue, M.; Yang, T.; Luo, X.; Fan, Y.; Meng, Y.; Liu, W.; Lin, G.; Li, B.; et al. Effects of Dietary Saccharomyces cerevisiae YFI-SC2 on the Growth Performance, Intestinal Morphology, Immune Parameters, Intestinal Microbiota, and Disease Resistance of Crayfish (Procambarus clarkia). Animals 2021, 11, 1963. https://doi.org/10.3390/ani11071963
Xu Y, Li Y, Xue M, Yang T, Luo X, Fan Y, Meng Y, Liu W, Lin G, Li B, et al. Effects of Dietary Saccharomyces cerevisiae YFI-SC2 on the Growth Performance, Intestinal Morphology, Immune Parameters, Intestinal Microbiota, and Disease Resistance of Crayfish (Procambarus clarkia). Animals. 2021; 11(7):1963. https://doi.org/10.3390/ani11071963
Chicago/Turabian StyleXu, Yan, Yiqun Li, Mingyang Xue, Tao Yang, Xiaowen Luo, Yuding Fan, Yan Meng, Wenzhi Liu, Ge Lin, Bo Li, and et al. 2021. "Effects of Dietary Saccharomyces cerevisiae YFI-SC2 on the Growth Performance, Intestinal Morphology, Immune Parameters, Intestinal Microbiota, and Disease Resistance of Crayfish (Procambarus clarkia)" Animals 11, no. 7: 1963. https://doi.org/10.3390/ani11071963
APA StyleXu, Y., Li, Y., Xue, M., Yang, T., Luo, X., Fan, Y., Meng, Y., Liu, W., Lin, G., Li, B., Zeng, L., & Zhou, Y. (2021). Effects of Dietary Saccharomyces cerevisiae YFI-SC2 on the Growth Performance, Intestinal Morphology, Immune Parameters, Intestinal Microbiota, and Disease Resistance of Crayfish (Procambarus clarkia). Animals, 11(7), 1963. https://doi.org/10.3390/ani11071963