
Transport and Recovery of Turbot (Scophthalmus maximus) Sedated with MS-222 and Eugenol: Effects on Intermediary Metabolism and Osmoregulation




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Turbot
2.2. Measurements and Analytical Procedures
2.2.1. Sample Collection
2.2.2. Chemical Composition of Flesh
2.2.3. Flesh pH
2.2.4. Biochemical Analysis of Flesh Samples
2.2.5. Determination of Nucleotides
2.2.6. Determination of Serum Biochemical Indicators
2.2.7. Statistical Analysis
3. Results
3.1. Chemical Composition of Flesh
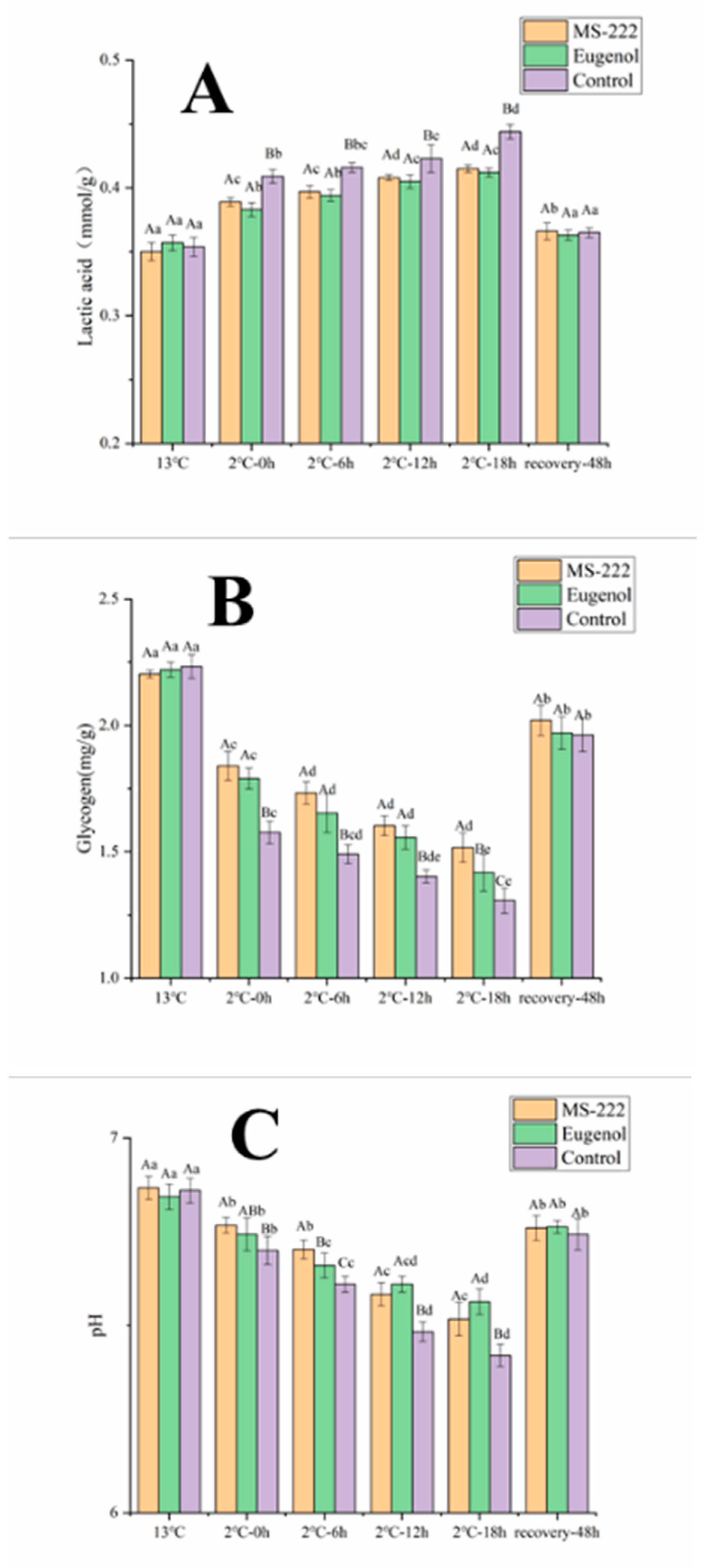
3.2. Flesh Biochemical Indicators
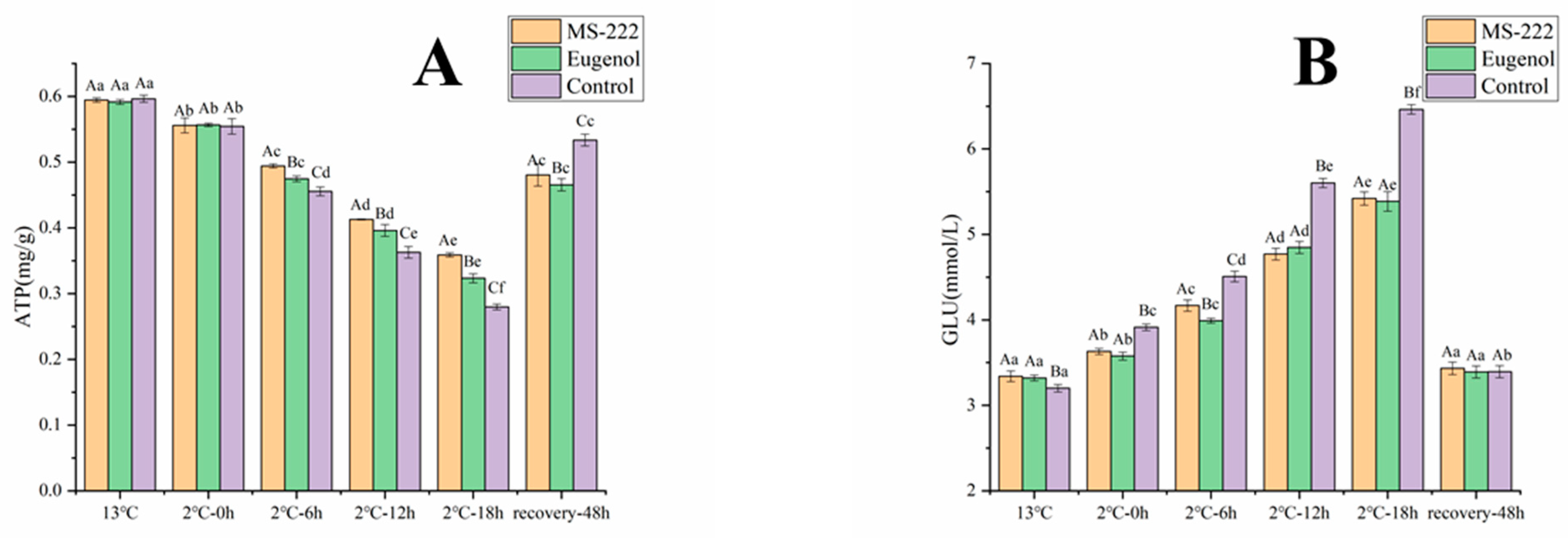
3.3. Changes in Blood Glucose and Flesh ATP in Turbot during Simulated Transport
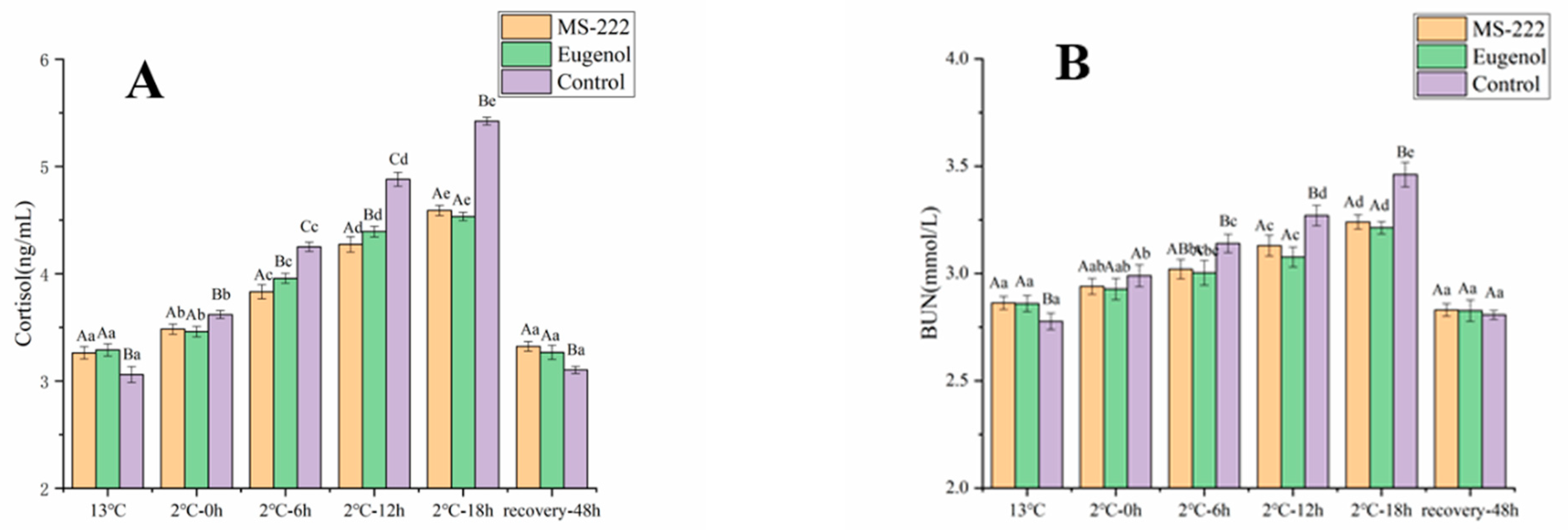
3.4. Changes of Cortisol and BUN in Turbot during Waterless Transport
3.5. Changes of Nucleotides in Turbot
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- He, R.P.; Su, Y.P.; Wang, A.L.; Lei, B.; Cui, K.P. Survival and serum biochemical responses of spotted sea bass Lateolabrax maculatus during simulated waterless live transportation. Aquacult. Res. 2020, 51, 3495–3505. [Google Scholar] [CrossRef]
- Refaey, M.M.; Tian, X.; Tang, R.; Li, D.P. Changes in physiological responses, muscular composition and flesh quality of channel catfish Ictalurus punctatus suffering from transport stress. Aquaculture 2017, 478, 9–15. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhang, X.S.; Nga, M.T.T.; Yu, H.R. Development and evaluation of key ambient factors online monitoring system in live Urechis unicinctus transportation strategies. Comput. Electron. Agric. 2018, 145, 43–52. [Google Scholar] [CrossRef]
- Nie, X.B.; Lei, J.L.; Chen, S.X.; Zhang, Y.T.; Zhang, C.F.; Hong, W.S. Physiological, proteomic, and gene expression analysis of turbot (Scophthalmus maximus) in response to cold acclimation. Aquaculture 2018, 495, 281–287. [Google Scholar] [CrossRef]
- De Oliveira, C.P.B.; Lemos, C.H.D.; Silva, A.F.E.; de Souza, S.A.; Albinati, A.C.L.; Lima, A.O.; Copatti, C.E. Use of eugenol for the anaesthesia and transportation of freshwater angelfish (Pterophyllum scalare). Aquaculture 2019, 513, 6. [Google Scholar] [CrossRef]
- Favero, G.C.; Silva, W.D.E.; Boaventura, T.P.; Leme, F.D.P.; Luz, R.K. Eugenol or salt to mitigate stress during the transport of juvenile Lophiosilurus alexandri, a Neotropical carnivorous freshwater catfish. Aquaculture 2019, 512, 6. [Google Scholar] [CrossRef]
- Ribeiro, P.A.P.; Miranda, K.C.; De Melo, D.C.; Luz, R.K. Efficiency of eugenol as anesthetic for the early life stages of Nile tilapia (Oreochromis niloticus). An. Acad. Bras. Cienc. 2015, 87, 529–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovic, N.T.; Strunjak-Perovic, I.; Coz-Rakovac, R.; Barisic, J.; Jadan, M.; Berakovic, A.P.; Klobucar, R.S. Tricaine methane-sulfonate (MS-222) application in fish anaesthesia. J. Appl. Ichthyol. 2012, 28, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Park, I.-S.; Gil, H.W.; Lee, T.H.; Nam, Y.K.; Lim, S.G.; Kim, D.S. Effects of Clove Oil and Lidocaine-HCl Anesthesia on Water Parameter during Simulated Transportation in the Marine Medaka, Oryzias dancena. Dev. Reprod. 2017, 21, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Prystay, T.S.; Elvidge, C.K.; Twardek, W.M.; Logan, J.M.; Reid, C.H.; Clarke, S.H.; Foster, J.G.; Cooke, E.L.L.; Cooke, S.J. Comparison of the Behavioral Consequences and Recovery Patterns of Largemouth Bass Exposed to MS-222 or Electrosedation. Trans. Am. Fish. Soc. 2017, 146, 556–566. [Google Scholar] [CrossRef]
- Rozynski, M.; Ziomek, E.; Demska-Zakes, K.; Zakes, Z. Impact of inducing general anaesthesia with MS-222 on haematological and biochemical parameters of pikeperch (Sander lucioperca). Aquacult. Res. 2019, 50, 2125–2132. [Google Scholar] [CrossRef]
- Jacobsen, J.V.; Steen, K.; Nilssen, K.J. Anaesthetic efficacy of Aqui-S, Benzoak, and MS-222 on lumpfish (Cyclopterus lumpus) fries. Impact from temperature, salinity, and fasting. PLoS ONE 2019, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Park, I.-S. The Anesthetic Effects of Clove Oil and MS-222 on Far Eastern Catfish, Silurus asotus. Dev. Reprod. 2019, 23, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Feng, G.; Zhuang, P.; Zhang, L.; Kynard, B.; Shi, X.; Duan, M.; Liu, J.; Huang, X. Effect of anaesthetics MS-222 and clove oil on blood biochemical parameters of juvenile Siberian sturgeon (Acipenser baerii). J. Appl. Ichthyol. 2011, 27, 595–599. [Google Scholar] [CrossRef]
- Palić, D.; Herolt, D.M.; Andreasen, C.B.; Menzel, B.W.; Roth, J.A. Anesthetic efficacy of tricaine methanesulfonate, metomidate and eugenol: Effects on plasma cortisol concentration and neutrophil function in fathead minnows (Pimephales promelas Rafinesque, 1820). Aquaculture 2006, 254, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Teles, M.; Oliveira, M.; Jerez-Cepa, I.; Franco-Martinez, L.; Tvarijonaviciute, A.; Tort, L.; Mancera, J.M. Transport and Recovery of Gilthead Sea Bream (Sparus aurata L.) Sedated With Clove Oil and MS222: Effects on Oxidative Stress Status. Front Physiol. 2019, 10, 523. [Google Scholar] [CrossRef]
- Vidal, L.V.O.; Albinati, R.C.B.; Albinati, A.C.L.; de Lira, A.D.; de Almeida, T.R.; Santos, G.B. Eugenol as an anesthetic for Nile tilapia. Pesqui. Agropecu. Bras. 2008, 43, 1069–1074. [Google Scholar] [CrossRef] [Green Version]
- Shukry, M.; Abd El-Kader, M.F.; Hendam, B.M.; Dawood, M.A.O.; Farrag, F.A.; Aboelenin, S.M.; Soliman, M.M.; Abdel-Latif, H.M.R. Dietary Aspergillus oryzae Modulates Serum Biochemical Indices, Immune Responses, Oxidative Stress, and Transcription of HSP70 and Cytokine Genes in Nile Tilapia Exposed to Salinity Stress. Animals 2021, 11, 1621. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Bai, J.; Ao, Z.; Wei, Z.; Hu, Y.; Liu, S. Effects of Dietary Paper Mulberry (Broussonetia papyrifera) on Growth Performance and Muscle Quality of Grass Carp (Ctenopharyngodon idella). Animals 2021, 11, 1655. [Google Scholar] [CrossRef]
- Liu, W.; Shen, Y.; Li, N.; Mei, J.; Xie, J. Application of Gelatin Incorporated with Red Pitaya Peel Methanol Extract as Edible Coating for Quality Enhancement of Crayfish Procambarus clarkii during Refrigerated Storage. J. Food Qual. 2019, 2019, 1–8. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Opinion of the Scientific Panel on Animal Health and Welfare (AHAW) on a request from the Commission related with the risks of poor welfare in intensive calf farming systems. EFSA J. Eur. Food Saf. Auth. 2006, 4, 366. [Google Scholar]
- Jrpeland, G.; Imsland, A.; Stien, L.H.; Bleie, H.; Roth, B.J.A.R. Effects of filleting method, stress, storage and season on the quality of farmed Atlantic cod (Gadus morhua L.). Aquac. Res. 2015, 46, 1597–1607. [Google Scholar] [CrossRef]
- Kristoffersen, S.; Tobiassen, T.; Steinsund, V.; Olsen, R.L. Slaughter stress, postmortem muscle pH and rigor development in farmed Atlantic cod (Gadus morhua L.). Int. J. Food Sci. Technol. 2006, 41, 861–864. [Google Scholar] [CrossRef]
- Bonga, S.E.W. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef] [PubMed]
- Moraes, G.; Avilez, I.M.; Altran, A.E.; Barbosa, C.C. Biochemical and hematological responses of the banded knife fish Gymnotus carapo (Linnaeus, 1758) exposed to environmental hypoxia. Braz. J. Biol. 2002, 62, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Varga, D.; Szabo, A.; Hancz, C.; Jeney, Z.; Ardo, L.; Molnar, M.; Molnar, T. Impact of Handling and Pre-Mortal Stress on the Flesh Quality of Common Carp (Cyprinus carpio L.). Isr. J. Aquacult. Bamidgeh. 2014, 66, 20746. [Google Scholar]
- Speers-Roesch, B.; Sandblom, E.; Lau, G.Y.; Farrell, A.P.; Richards, J.G. Effects of environmental hypoxia on cardiac energy metabolism and performance in tilapia. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2010, 298, R104–R119. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, J.; Reisine, T.D. Stress hormones: Their interaction and regulation. Science 1984, 224, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.L.R.; Rezende, F.P.; Moron, S.E. Stress-related physiological and histological responses of tambaqui (Colossoma macropomum) to transportation in water with tea tree and clove essential oil anesthetics. Aquaculture 2020, 523, 10. [Google Scholar] [CrossRef]
- Bortoletti, M.; Maccatrozzo, L.; Radaelli, G.; Caberlotto, S.; Bertotto, D. Muscle Cortisol Levels, Expression of Glucocorticoid Receptor and Oxidative Stress Markers in the Teleost Fish Argyrosomus regius Exposed to Transport Stress. Animals 2021, 11, 1160. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.; Jones, J.; Jacobson, S.; Reimschuessel, R. Elevated blood urea nitrogen (BUN) levels in goldfish as an indicator of gill dysfunction. J. Aquat. Anim. Health 1999, 11, 52–60. [Google Scholar] [CrossRef]
- McCormick, S.D. Endocrine control of osmoregulation in teleost fish. Am. Zool. 2001, 41, 781–794. [Google Scholar] [CrossRef]
- Barton, B.A.; Haukenes, A.H.; Parsons, B.G.; Reed, J.R. Plasma cortisol and chloride stress responses in juvenile walleyes during capture, transport, and stocking procedures. N. Am. J. Aqualcult. 2003, 65, 210–219. [Google Scholar] [CrossRef]
- Gomes, L.D.; Araujo-Lima, C.; Roubach, R.; Urbinati, E.C. Assessment on the effect of salt and density on tambaqui fish transportation. Pesqui. Agropecu. Bras. 2003, 38, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Wang, Q.; Cao, J.; Mei, J.; Xie, J. Effects of Ascorbic Acid and beta-1,3-Glucan on Survival, Physiological Response and Flesh Quality of Cultured Tiger Grouper (Epinephelus fuscoguttatus) during Simulated Transport in Water. Biology 2020, 9, 18. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Indicators/Transport | Samples | 13 °C | 2 °C—0 h | 2 °C—6 h | 2 °C—12 h | 12 °C—18 h | Recovery—48 h |
|---|---|---|---|---|---|---|---|
| Water content (%) | Control | 78.89 ± 0.26 Aa | 78.11 ± 0.10 Ab | 77.87 ± 0.11 Ac | 77.63 ± 0.06 Ad | 77.51 ± 0.08 Ad | 78.80 ± 0.12 Aa |
| MS-222 | 78.77 ± 0.19 Aa | 78.32 ± 0.08 Bb | 78.11 ± 0.12 ABc | 77.93 ± 0.13 Bd | 77.75 ± 0.05 Bd | 78.67 ± 0.09 Aa | |
| Eugenol | 78.74 ± 0.33 Aa | 78.27 ± 0.11 ABb | 78.10 ± 0.05 Bc | 77.95 ± 0.07 Bd | 77.80 ± 0.10 Bd | 78.66 ± 0.15 Aa | |
| Crude fat (%) | Control | 1.44 ± 0.06 Aa | 1.24 ± 0.04 Ab | 1.17 ± 0.03 Abc | 1.10 ± 0.04 Acd | 1.04 ± 0.05 Ad | 1.39 ± 0.08 Aa |
| MS-222 | 1.47 ± 0.05 Aa | 1.32 ± 0.07 Bb | 1.24 ± 0.05 Abc | 1.16 ± 0.03 Acd | 1.10 ± 0.06 Ad | 1.41 ± 0.07 Aa | |
| Eugenol | 1.40 ± 0.03 Aa | 1.28 ± 0.05 ABb | 1.22 ± 0.04 Ab | 1.18 ± 0.05 Abc | 1.12 ± 0.05 Ac | 1.37 ± 0.06 Aa | |
| Crude protein (%) | Control | 18.65 ± 0.11 Aa | 18.08 ± 0.10 Ab | 17.95 ± 0.05 Ab | 17.83 ± 0.16 Abc | 17.62 ± 0.10 Ac | 18.51 ± 0.09 Aa |
| MS-222 | 18.61 ± 0.07 Aa | 18.19 ± 0.09 ABb | 18.06 ± 0.14 ABb | 17.98 ± 0.11 Ab | 17.89 ± 0.14 ABb | 18.60 ± 0.11 Aa | |
| Eugenol | 18.66 ± 0.09 Aa | 18.26 ± 0.0.05 Bb | 18.13 ± 0.08 Bb | 18.00 ± 0.10 Abc | 17.92 ± 0.07 Bc | 18.69 ± 0.13 Aa |
| Transport (h) | Sample | IMP (mg/100 g) | TAV | AMP (mg/100 g) | TAV |
|---|---|---|---|---|---|
| 13 °C | Control | 319.44 ± 11.55 Ab | 12.78 | 24.73 ± 0.71 Aa | 0.49 |
| MS-222 | 312.45 ± 6.11 Ab | 12.50 | 20.96 ± 0.76 Ba | 0.42 | |
| Eugenol | 310.37 ± 5.20 Ab | 12.41 | 21.62 ± 1.09 Ba | 0.43 | |
| 2 °C—0 h | Control | 287.81 ± 7.29 Aa | 11.51 | 19.41 ± 1.14 Ab | 0.39 |
| MS-222 | 296.17 ± 5.16 Aa | 11.85 | 18.23 ± 0.97 Ab | 0.36 | |
| Eugenol | 292.50 ± 5.65 Aa | 11.70 | 18.96 ± 1.63 Abc | 0.38 | |
| 2 °C—6 h | Control | 343.39 ± 7.86 Ac | 13.74 | 16.65 ± 0.80 Ac | 0.33 |
| MS-222 | 325.91 ± 5.25 Bc | 13.04 | 15.99 ± 0.63 Ac | 0.32 | |
| Eugenol | 325.63 ± 3.31 Bc | 13.03 | 16.74 ± 1.41 Acd | 0.33 | |
| 2 °C—12 h | Control | 356.18 ± 6.04 Acd | 14.25 | 13.81 ± 0.47 Bd | 0.28 |
| MS-222 | 340.62 ± 2.36 Bd | 13.62 | 14.55 ± 0.48 ABc | 0.29 | |
| Eugenol | 341.49 ± 5.69 Bd | 13.66 | 15.41 ± 1.21 Ad | 0.31 | |
| 2 °C—18 h | Control | 369.76 ± 7.22 Ad | 14.79 | 16.92 ± 1.39 Bc | 0.34 |
| MS-222 | 349.67 ± 1.44 Be | 13.99 | 18.11 ± 1.19 Ab | 0.36 | |
| Eugenol | 348.97 ± 6.69 Bd | 13.96 | 18.2 ± 0.23 Abc | 0.36 | |
| Recovery—48 h | Control | 320.00 ± 13.62 Ab | 12.80 | 23.24 ± 0.56 Ba | 0.46 |
| MS-222 | 322.78 ± 3.80 Ac | 12.91 | 20.64 ± 0.83 Aa | 0.41 | |
| Eugenol | 314.93 ± 4.14 Ab | 12.60 | 20.91 ± 1.87 Aab | 0.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, J.; Wang, Q.; Qiu, W.; Mei, J.; Xie, J. Transport and Recovery of Turbot (Scophthalmus maximus) Sedated with MS-222 and Eugenol: Effects on Intermediary Metabolism and Osmoregulation. Animals 2021, 11, 2228. https://doi.org/10.3390/ani11082228
Cao J, Wang Q, Qiu W, Mei J, Xie J. Transport and Recovery of Turbot (Scophthalmus maximus) Sedated with MS-222 and Eugenol: Effects on Intermediary Metabolism and Osmoregulation. Animals. 2021; 11(8):2228. https://doi.org/10.3390/ani11082228
Chicago/Turabian StyleCao, Jie, Qi Wang, Weiqiang Qiu, Jun Mei, and Jing Xie. 2021. "Transport and Recovery of Turbot (Scophthalmus maximus) Sedated with MS-222 and Eugenol: Effects on Intermediary Metabolism and Osmoregulation" Animals 11, no. 8: 2228. https://doi.org/10.3390/ani11082228
APA StyleCao, J., Wang, Q., Qiu, W., Mei, J., & Xie, J. (2021). Transport and Recovery of Turbot (Scophthalmus maximus) Sedated with MS-222 and Eugenol: Effects on Intermediary Metabolism and Osmoregulation. Animals, 11(8), 2228. https://doi.org/10.3390/ani11082228