
Effects of Tourism on the Habitat Use by a Threatened Large Rodent at a World Heritage Site

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
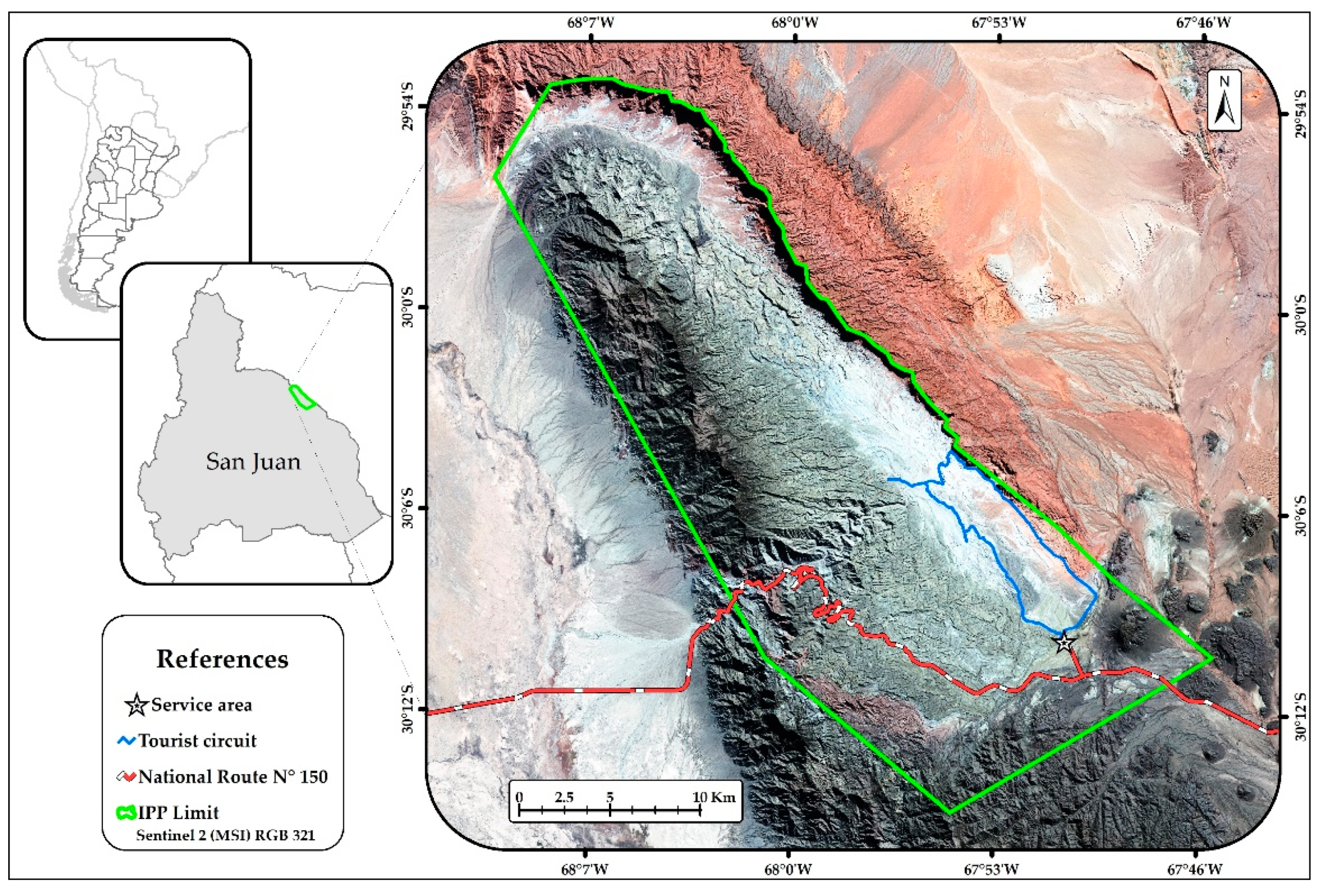
2. Materials and Methods
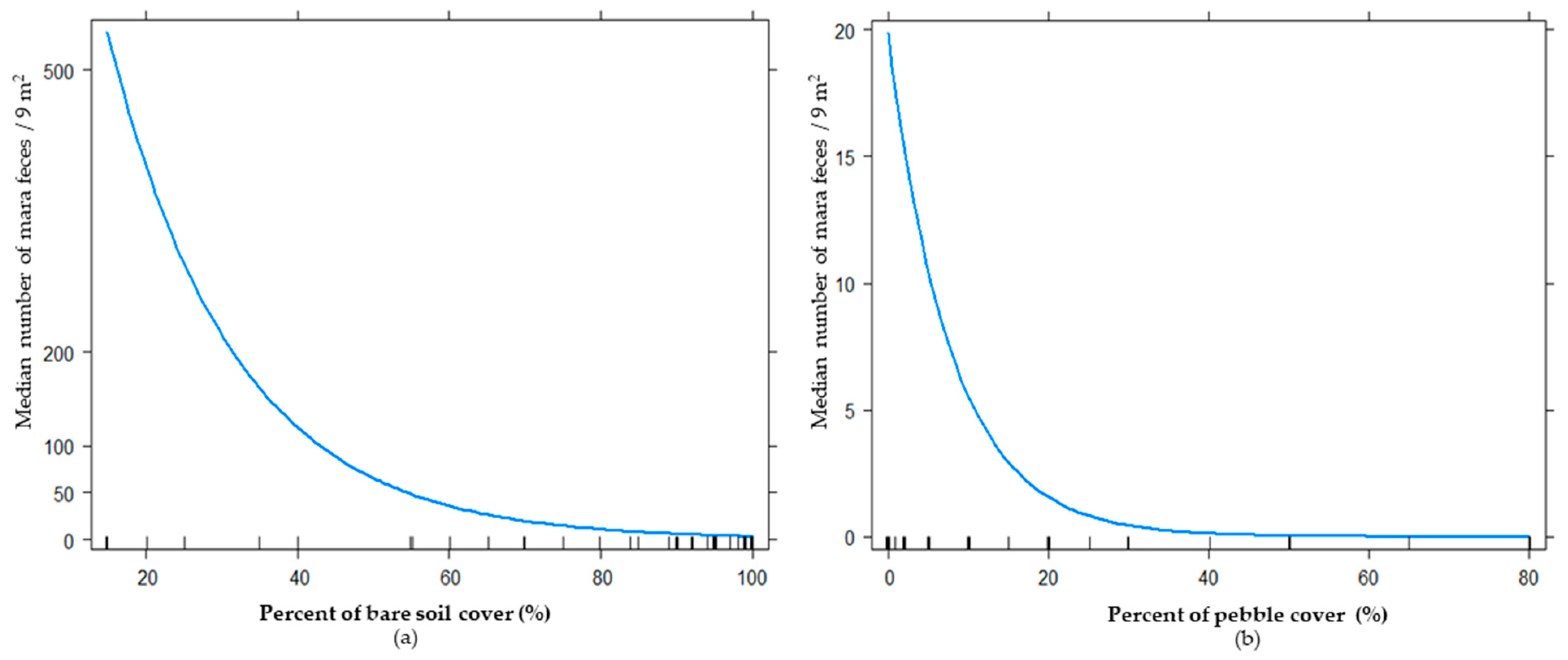
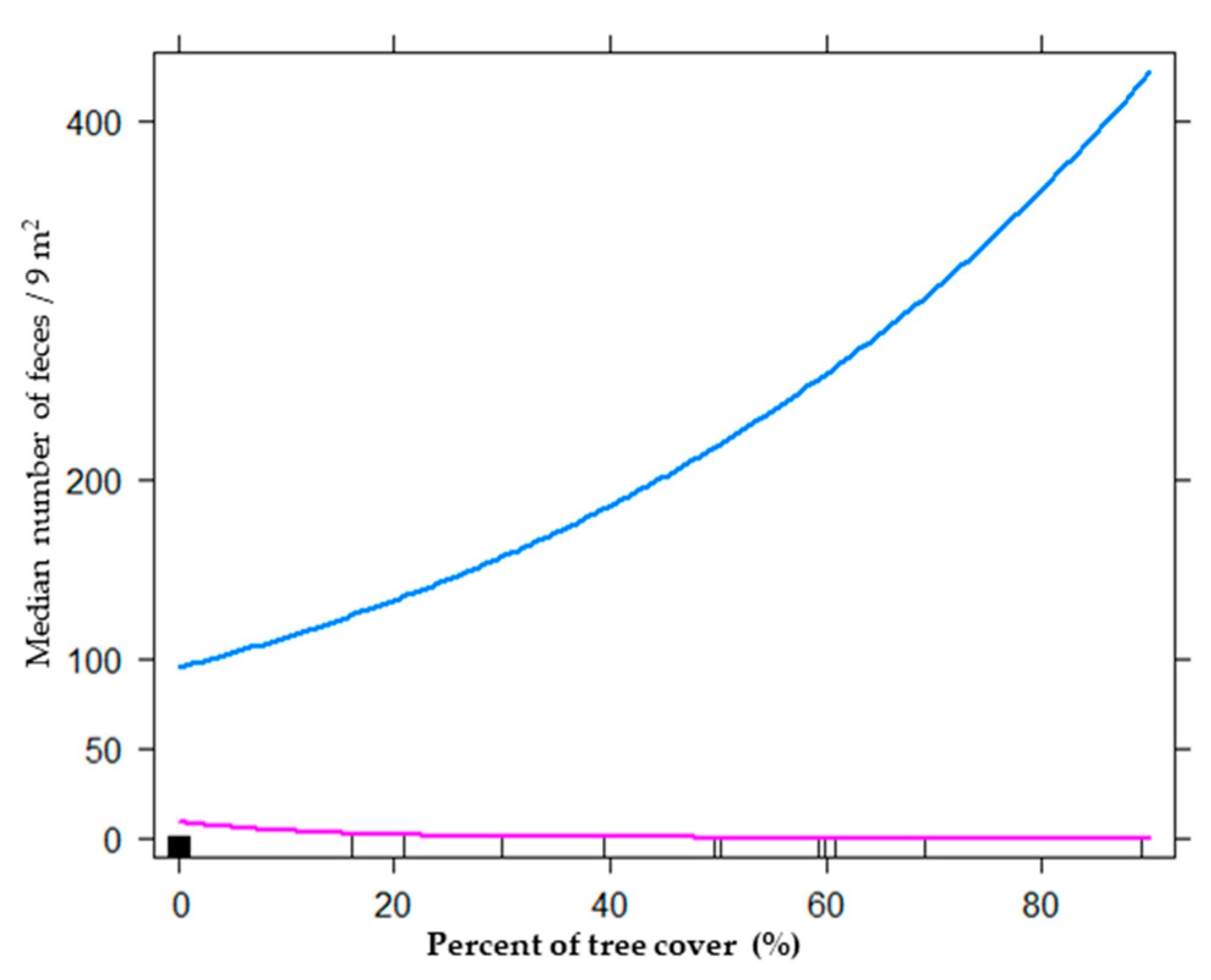
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Law, B.; Dickman, C. The Use of Habitat Mosaics by Terrestrial Vertebrate Fauna: Implications for Conservation and Management. Biodivers. Conserv. 1998, 7, 323–333. [Google Scholar] [CrossRef]
- de Gabriel Hernando, M.; Karamanlidis, A.; Grivas, K.; Krambokoukis, L.; Papakostas, G.; Beecham, J. Habitat Use and Selection Patterns Inform Habitat Conservation Priorities of an Endangered Large Carnivore in Southern Europe. Endang. Species. Res. 2021, 44, 203–215. [Google Scholar] [CrossRef]
- Alonso Roldán, V.; Udrizar Sauthier, D.E.; Giannoni, S.M.; Campos, C.M. Dolichotis patagonum. In Categorización 2019 de los Mamíferos de Argentina Según su Riesgo de Extinción. Lista Roja de los Mamíferos de Argentina; SAyDS–SAREM: Buenos Aires, Argentina, 2019; Volume 337. (In Spanish) [Google Scholar]
- Taber, A.B.; Macdonald, D.W. Communal Breeding in the Mara, Dolichotis patagonum. J. Zool. 1992, 227, 439–452. [Google Scholar] [CrossRef]
- Gatica, A.; Denkiewicz, N.; Ochoa, A. Breeding Behavior of Mara [Dolichotis patagonum (Zimmermann, 1780)] in the Monte-Chaco Ecotone of Argentina. Mammal. Study 2019, 44, 1. [Google Scholar] [CrossRef]
- Dubost, G.; Genest, H. Le Comportement Social d’une Colonie de Maras Dolichotis patagonum Z. Dans Le Parc de Branféré. Z. Für Tierpsychol. 1974, 35, 225–302. [Google Scholar] [CrossRef]
- Campos, C.M.; Tognelli, M.F.; Ojeda, R.A. Dolichotis patagonum. Mamm. Species 2001, 652, 1–5. [Google Scholar] [CrossRef]
- Chagas, K.; Vassallo, A.; Becerra, F.; Echeverría, A.; Loguercio, M.; Rocha-Barbosa, O. Locomotion in the Fastest Rodent, the Mara Dolichotis patagonum (Caviomorpha; Caviidae; Dolichotinae). Mastozool. Neotrop. 2019, 26, 65–79. [Google Scholar]
- Sombra, M.S.; Mangione, A.M. Obsessed with Grasses? The Case of Mara Dolichotis patagonum (Caviidae: Rodentia). Rev. Chil. De Hist. Nat. 2012, 78, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Reus, M.L.; Peco, B.; de los Ríos, C.; Giannoni, S.M.; Campos, C.M. Trophic Interactions between Two Medium-Sized Mammals: The Case of the Native Dolichotis patagonum and the Exotic Lepus europaeus in a Hyper-Arid Ecosystem. Acta Theriol. 2013, 58, 205–214. [Google Scholar] [CrossRef]
- Roach, N. Dolichotis patagonum. In The IUCN Red List of Threatened Species; 2016; Available online: https://www.iucnredlist.org/species/6785/22190337 (accessed on 17 July 2021).
- Rood, J.P. Ecological and Behavioural Comparisons of Three Genera of Argentine Cavies. Anim. Behav. Monogr. 1972, 5, 3–83. [Google Scholar] [CrossRef]
- Ojeda, R.A. Familia Caviidae Waterhouse. In Los Mamíferos de Argentina, Sistemática y Distribución; Barquez, R.M., Díaz, M.M., Ojeda, Y.R.A., Eds.; SAREM: Mendoza, Argentina, 2006; pp. 206–210. (In Spanish) [Google Scholar]
- Kufner, M.B.; Chambouleyron, M. Actividad Espacial de Dolichotis patagonum en Relación a la Estructura de la Vegetación en el Monte Argentino. Stud. Neotrop. Fauna Environ. 1991, 26, 249–255. (In Spanish) [Google Scholar] [CrossRef]
- Baldi, R. Breeding Success of the Endemic Mara Dolichotis patagonum in Relation to Habitat Selection: Conservation Implications. J. Arid Environ. 2007, 68, 9–19. [Google Scholar] [CrossRef]
- Rodríguez, D. Modeling Habitat Use of the Threatened and Endemic Mara (Dolichotis patagonum, Rodentia, Caviidae) in Agricultural Landscapes of Monte Desert. J. Arid Environ. 2009, 73, 444–448. [Google Scholar] [CrossRef]
- Tabeni, S.; Spirito, F.; Ojeda, R.A. Conservation of Small and Medium-Sized Mammals Following Native Woodland Regrowth: A Case Study in a Long-Term UNESCO Biosphere Reserve, Argentina. J. Arid Environ. 2013, 88, 250–253. [Google Scholar] [CrossRef]
- Curry, B.; Moore, W.; Bauer, J.; Cosgriff, K.; Lipscombe, N. Modelling Impacts of Wildlife Tourism on Animal Communities: A Case Study from Royal Chitwan National Park, Nepal. J. Sustain. Tour. 2001, 9, 514–529. [Google Scholar] [CrossRef]
- Buckley, R. Impacts Positive and Negative: Links between Ecotourism and Environment. Environ. Impacts Ecotourism 2004, 2, 5–14. [Google Scholar]
- Severcan, Ç.; Yamaç, E. The Effects of Flock Size and Human Presence on Vigilance and Feeding Behavior in the Eurasian Coot (Fulica atra L.) during Breeding Season. Acta Ethologica 2011, 14, 51–56. [Google Scholar] [CrossRef]
- Semeniuk, C.A.D. A Linked Model of Animal Ecology and Human Behavior for the Management of Wildlife Tourism. Ecol. Model. 2010, 221, 2699–2713. [Google Scholar] [CrossRef]
- Heil, L.; Fernández-Juricic, E.; Renison, D.; Cingolani, A.M.; Blumstein, D.T. Avian responses to tourism in the biogeographically isolated high Córdoba Mountains, Argentina. Biodivers. Conserv. 2006, 16, 1009–1026. [Google Scholar] [CrossRef]
- Frid, A.; Dill, L. Human-Caused Disturbance Stimuli as a Form of Predation Risk. Conserv. Ecol. 2002, 6, 11. [Google Scholar] [CrossRef]
- Stankowich, T. Ungulate Flight Responses to Human Disturbance: A Review and Meta-Analysis. Biol. Conserv. 2008, 141, 2159–2173. [Google Scholar] [CrossRef]
- Christ, C.; Hillel, O.; Matus, S.; Sweeting, J. Tourism and Biodiversity: Mapping Tourism’s Global Footprint; Conservation International (CI): Washington, DC, USA, 2003; pp. 1–53. [Google Scholar]
- Barros, A.; Monz, C.; Pickering, C. Is Tourism Damaging Ecosystems in the Andes? Current Knowledge and an Agenda for Future Research. Ambio 2015, 44, 82–98. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wu, B.; Cai, L. Tourism Development of World Heritage Sites in China: A Geographic Perspective. Tour. Manag. 2008, 29, 308–319. [Google Scholar] [CrossRef]
- Cortez, E.; Borghi, C.E.; Giannoni, S.M. Plan. de Manejo Parque Provincial Ischigualasto, Fase I y II; Ente Autárquico Ischigualasto, Gobierno de San Juan: San Juan, Argentina, 2005; p. 225. (In Spanish) [Google Scholar]
- Giaccardi, M.; Alcober, O.; Colombi, C.; Duran, V.; Giannoni, S.M. Plan. de Manejo Del Parque Natural Provincial Ischigualasto. Patrimonio Natural de La Humanidad, Provincia de San Juan; Ente Autárquico Ischigualasto, Gobierno de San Juan: San Juan, Argentina, 2015; Volume 1, pp. 1–290. (In Spanish) [Google Scholar]
- Márquez, J.; Carretero, E.M.; Dalmasso, A.; Pastrán, G.; Ortiz, G. Las Áreas Protegidas de la Provincia de San Juan (Argentina) II. La Vegetación del Parque Provincial Ischigualasto. Multequina 2005, 14, 1–27. (In Spanish) [Google Scholar]
- Acebes, P.; Traba, J.; Peco, B.; Reus, M.L.; Giannoni, S.M.; Malo, J.E. Abiotic Gradients Drive Floristic Composition and Structure of Plant Communities in the Monte Desert. Rev. Chil. Hist. Nat. 2010, 83, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Adamczewski, J.Z.; Gates, C.C.; Soutar, B.M.; Hudson, R.J. Limiting Effects of Snow on Seasonal Habitat Use and Diets of Caribou (Rangifer tarandus groenlandicus) on Coats Island, Northwest Territories, Canada. Can. J. Zool. 1988, 66, 1986–1996. [Google Scholar] [CrossRef]
- Bailey, R.E.; Putman, R.J. Estimation of Fallow Deer (Dama dama) Populations from Faecal Accumulation. J. Appl. Ecol. 1981, 18, 697–702. [Google Scholar] [CrossRef]
- Ovejero, R.J.; Acebes, P.; Malo, J.E.; Traba, J.; Torres, M.E.M.; Borghi, C.E. Lack of Feral Livestock Interference with Native Guanaco during the Dry Season in a South American Desert. Eur. J. Wildl. Res. 2011, 57, 1007–1015. [Google Scholar] [CrossRef]
- Cappa, F.M.; Borghi, C.E.; Giannoni, S.M. How Roads Affect the Spatial Use of the Guanaco in a South American Protected Area: Human Connectivity vs Animal Welfare. Diversity 2019, 11, 110. [Google Scholar] [CrossRef] [Green Version]
- Kaseloo, P.A. Synthesis of Noise Effects on Wildlife Populations. In Proceedings of the 2005 International Conference on Ecology and Transportation, North Carolina State University, Raleigh, NC, USA, 29 August–5 September 2006; pp. 33–35. [Google Scholar]
- Dyke, F.; Brocke, R.; Shaw, H.; Ackerman, B.; Hemker, T.; Lindzey, F. Reactions of Mountain Lions to Logging and Human Activity. J. Wildl. Manag. 1986, 50, 95–102. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. A Practical Information-Theoretic Approach. In Model Selection and Multimodel Inference, 2nd ed; Springer: New York, NY, USA, 2002; pp. 1–488. [Google Scholar]
- Barton, K. MuMIn: Multi-Model. Inference. R Package Version 1.43.6. 2019. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 10 April 2019).
- Harrell, F.E., Jr. Rms: Regression Modeling Strategies. R package Version 5.1-3.1. 2019. Available online: https://CRAN.R-project.org/package=rms (accessed on 10 April 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 1 April 2019).
- Daciuk, J. Notas Faunísticas y Biotecnológicas de Península Valdés y Patagonia. XII. Mamíferos Colectados y Observados en la Península Valdés y Zona Litoral de Los Golfos San José y Nuevo (Provincia de Chubut, República Argentina). Physis 1974, 33, 23–39. (In Spanish) [Google Scholar]
- Kotliar, N.B.; Wiens, J.A. Multiple Scales of Patchiness and Patch Structure: A Hierarchical Framework for the Study of Heterogeneity. Oikos 1990, 59, 253–260. [Google Scholar] [CrossRef]
- Pedlar, J.H.; Fahrig, L.; Merriam, H.G. Raccoon Habitat Use at 2 Spatial Scales. J. Wildl. Manag. 1997, 61, 102–112. [Google Scholar] [CrossRef]
- Acebes, P.; Malo, J.E.; Traba, J. Trade-Offs between Food Availability and Predation Risk in Desert Environments: The Case of Polygynous Monomorphic Guanaco (Lama guanicoe). J. Arid Environ. 2013, 97, 136–142. [Google Scholar] [CrossRef]
- Ojeda, R.A.; Mares, M.A. Conservation of South American Mammals: Argentina as a Paradigm. Mamm. Biol. South. Am. 1982, 6, 505–521. [Google Scholar]
- García Fernández, J.J.; Ojeda, R.; Fraga, R.M.; Diaz, G.B.; Baigun, R.J. Libro Rojo de Mamíferos y Aves Amenazados de la Argentina; FUCEMA-Administración de Parques Nacionales: Buenos Aires, Argentina, 1997; ISBN 987-96325-0-8. (In Spanish) [Google Scholar]
- Berger, J.; Cunningham, C. Size-related Effects on Search Times in North American Grassland Female Ungulates. Ecology 1988, 69, 177–183. [Google Scholar] [CrossRef]
- Rogala, J.K.; Hebblewhite, M.; Whittington, J.; White, C.A.; Coleshill, J.; Musiani, M. Human Activity Differentially Redistributes Large Mammals in the Canadian Rockies National Parks. Ecol. Soc. 2011, 16, 16. [Google Scholar] [CrossRef] [Green Version]
- Cappa, F.M.; Giannoni, S.M.; Borghi, C.E. Effects of Roads on the Behaviour of the Largest South American Artiodactyl (Lama guanicoe) in an Argentine Reserve. Anim. Behav. 2017, 131, 131–136. [Google Scholar] [CrossRef]
- Suraci, J.P.; Clinchy, M.; Zanette, L.Y.; Wilmers, C.C. Fear of Humans as Apex Predators has Landscape-Scale Impacts from Mountain Lions to Mice. Ecol. Lett. 2019, 22, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, R.A.; Chillo, V.; Díaz, G. Libro Rojo de Los Mamíferos Amenazados de La Argentina, Sociedad Argentina para el Estudio de los Mamíferos (SAREM); Sociedad Argentina para el Estudio de los Mamíferos (SAREM): Mendoza, Argentina, 2012. (In Spanish) [Google Scholar]
- Borghi, C.E.; Campos, C.M.; Ortuño, N.; Beninato, V.; Andino, N.; Campos, V.; de los Ríos, C.; Cappa, F.; Giannoni, S.M. Efeitos Indiretos Sobre a Fauna do Corredor Bioceánico Central em uma Área Protegida do Deserto do Monte: PP Ischigualasto. Ecol. Estrad. Tendências E Pesqui. Lavras UFLA Lavras 2012, 314, 237–252. (In Spanish) [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Levels | K | AICc | ΔAICc | D2 |
|---|---|---|---|---|
| Plant Community | ||||
| Pebble Cover + Bare Soil | 4 | 486.82 | 0 | 15.42% |
| Null Model | 2 | 497.00 | 10.18 | 0% |
| Plant Strata | ||||
| Pebble Cover + Bare Soil | 4 | 486.82 | 0 | 15.42% |
| Null Model | 2 | 497.00 | 10.18 | 0% |
| Biological Form | ||||
| Tourism Activity + Pebble Cover + Bare Soil +Trees + Tourism Activity × Pebble Cover + Tourism Activity × Trees | 8 | 476.80 | 0 | 31.31% |
| Null Model | 2 | 497.00 | 10.18 | 0% |
| Floristic Composition | ||||
| Tourism Activity + Pebble Cover + Prosopis + Bare Soil + Tourism Activity × Pebble Cover + Tourism Activity × Prosopis | 8 | 476.80 | 0 | 31.31% |
| Null Model | 2 | 497.00 | 10.18 | 0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beninato, V.A.; Borghi, C.E.; Andino, N.; Pérez, M.A.; Giannoni, S.M. Effects of Tourism on the Habitat Use by a Threatened Large Rodent at a World Heritage Site. Animals 2021, 11, 2278. https://doi.org/10.3390/ani11082278
Beninato VA, Borghi CE, Andino N, Pérez MA, Giannoni SM. Effects of Tourism on the Habitat Use by a Threatened Large Rodent at a World Heritage Site. Animals. 2021; 11(8):2278. https://doi.org/10.3390/ani11082278
Chicago/Turabian StyleBeninato, Veronica A., Carlos E. Borghi, Natalia Andino, Mauricio A. Pérez, and Stella M. Giannoni. 2021. "Effects of Tourism on the Habitat Use by a Threatened Large Rodent at a World Heritage Site" Animals 11, no. 8: 2278. https://doi.org/10.3390/ani11082278
APA StyleBeninato, V. A., Borghi, C. E., Andino, N., Pérez, M. A., & Giannoni, S. M. (2021). Effects of Tourism on the Habitat Use by a Threatened Large Rodent at a World Heritage Site. Animals, 11(8), 2278. https://doi.org/10.3390/ani11082278