
The Fatty Acid-Based Erythrocyte Membrane Lipidome in Dogs with Chronic Enteropathy

 , ,
, ,  , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diagnostic Investigations and Therapeutic Trials
2.3. Fatty Acid-Based RBC Membrane Lipidome Analysis
2.4. Evaluation of the Fatty Acid Cluster, Homeostasis Indexes, and Enzymes Activity Indexes
2.5. Statistical Analysis
3. Results
3.1. Study Population
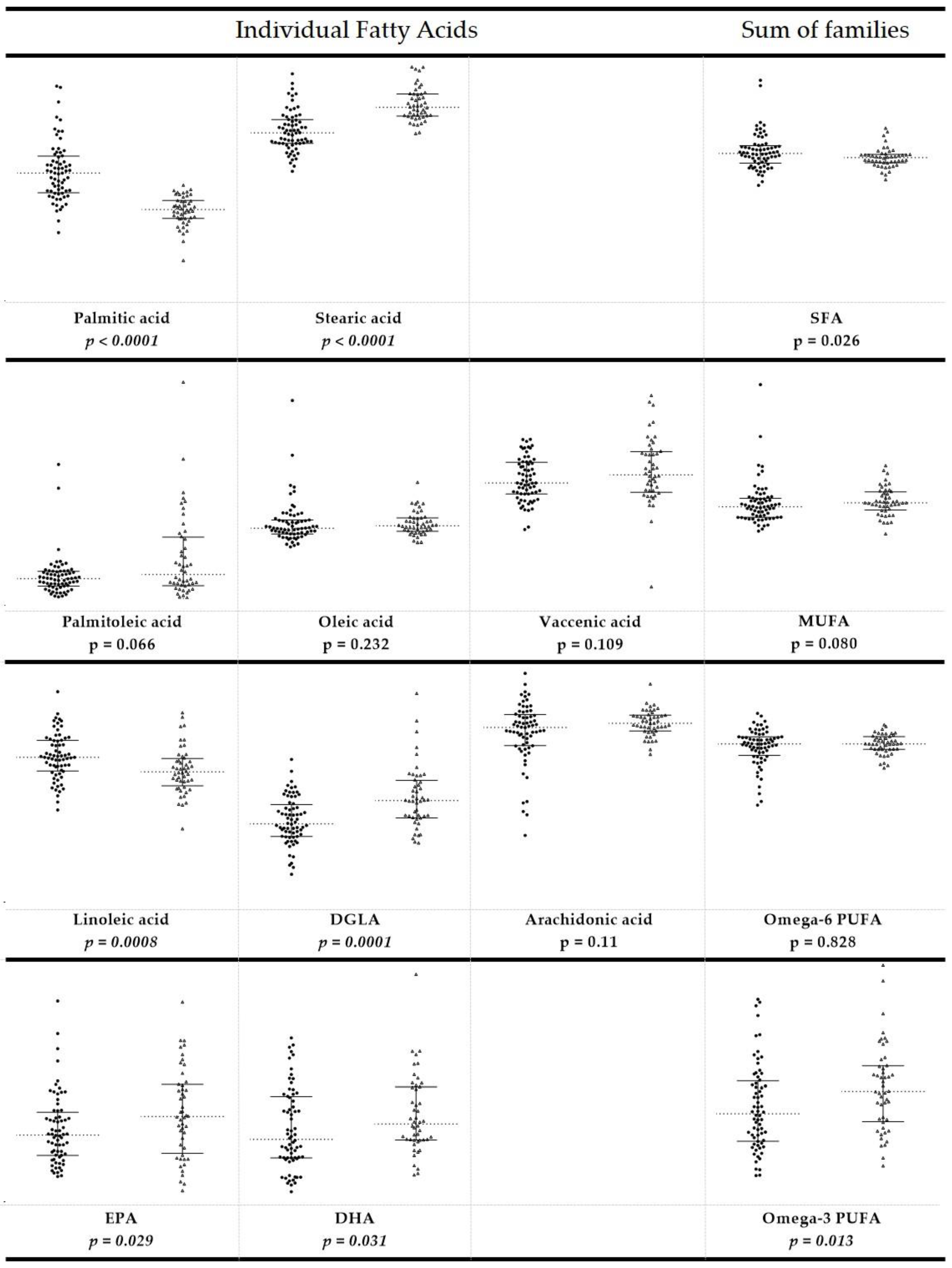
3.2. Fatty Acid-Based RBC Membrane Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dandrieux, J.R.S. Inflammatory bowel disease versus chronic enteropathy in dogs: Are they one and the same? J. Small Anim. Pr. 2016, 57, 589–599. [Google Scholar] [CrossRef]
- The WSAVA International Gastrointestinal Standardization Group; Washabau, R.J.; Day, M.J.; Willard, M.D.; Hall, E.; Jergens, A.E.; Mansell, J.; Minami, T.; Bilzer, T.W. Endoscopic, Biopsy, and Histopathologic Guidelines for the Evaluation of Gastrointestinal Inflammation in Companion Animals. J. Vet. Intern. Med. 2010, 24, 10–26. [Google Scholar] [CrossRef]
- Allenspach, K.; Wieland, B.; Gröne, A.; Gaschen, F. Chronic Enteropathies in Dogs: Evaluation of Risk Factors for Negative Outcome. J. Vet. Intern. Med. 2007, 21, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Craven, M.; Simpson, J.W.; Ridyard, A.E.; Chandler, M.L. Canine inflammatory bowel disease: Retrospective analysis of diagnosis and outcome in 80 cases (1995–2002). J. Small Anim. Pr. 2004, 45, 336–342. [Google Scholar] [CrossRef]
- Nakashima, K.; Hiyoshi, S.; Ohno, K.; Uchida, K.; Goto-Koshino, Y.; Maeda, S.; Mizutani, N.; Takeuchi, A.; Tsujimoto, H. Prognostic factors in dogs with protein-losing enteropathy. Vet. J. 2015, 205, 28–32. [Google Scholar] [CrossRef] [PubMed]
- German, A.; Hall, E.J.; Day, M.J. Chronic intestinal inflammation and intestinal disease in dogs. J. Vet. Intern. Med. 2003, 17, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.J. Gastrointestinal aspects of food allergy: A review. J. Small Anim. Pr. 1994, 35, 145–152. [Google Scholar] [CrossRef]
- Suchodolski, J.S.; Markel, M.E.; Garcia-Mazcorro, J.; Unterer, S.; Heilmann, R.M.; Dowd, S.; Kachroo, P.; Ivanov, I.; Minamoto, Y.; Dillman, E.M.; et al. The Fecal Microbiome in Dogs with Acute Diarrhea and Idiopathic Inflammatory Bowel Disease. PLoS ONE 2012, 7, e51907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scaioli, E.; Liverani, E.; Belluzzi, A. The Imbalance between n-6/n-3 Polyunsaturated Fatty Acids and Inflammatory Bowel Disease: A Comprehensive Review and Future Therapeutic Perspectives. Int. J. Mol. Sci. 2017, 18, 2619. [Google Scholar] [CrossRef] [Green Version]
- Dumusc, S.; Ontsouka, E.; Schnyder, M.; Hartnack, S.; Albrecht, C.; Bruckmaier, R.; Burgener, I. Cyclooxygenase-2 and 5-Lipoxygenase in Dogs with Chronic Enteropathies. J. Vet. Intern. Med. 2014, 28, 1684–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalak, A.; Mosińska, P.; Fichna, J. Polyunsaturated Fatty Acids and Their Derivatives: Therapeutic Value for Inflammatory, Functional Gastrointestinal Disorders, and Colorectal Cancer. Front. Pharmacol. 2016, 7, 459. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, M.J.C.R.; Caramujo, M.J. The Various Roles of Fatty Acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef] [Green Version]
- Nicolson, G.L.; Ash, M. Lipid Replacement Therapy: A natural medicine approach to replacing damaged lipids in cellular membranes and organelles and restoring function. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 1657–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreri, C.; Chatgilialoglu, C. Nutrilipidomics. In Membrane Lipidomics for Personalized Health; John Wiley & Sons Ltd: Hoboken, NJ, USA, 2015; pp. 127–133. ISBN 978. [Google Scholar]
- Ferreri, C.; Masi, A.; Sansone, A.; Giacometti, G.; LaRocca, A.V.; Menounou, G.; Scanferlato, R.; Tortorella, S.; Rota, D.; Conti, M.; et al. Fatty Acids in Membranes as Homeostatic, Metabolic and Nutritional Biomarkers: Recent Advancements in Analytics and Diagnostics. Diagnostics 2016, 7, 1. [Google Scholar] [CrossRef]
- Ferreri, C.; Chatgilialoglu, C. Role of fatty acid-based functional lipidomics in the development of molecular diagnostic tools. Expert Rev. Mol. Diagn. 2012, 12, 767–780. [Google Scholar] [CrossRef]
- Hu, C.; van der Heijden, R.; Wang, M.; van der Greef, J.; Hankemeier, T.; Xu, G. Analytical strategies in lipidomics and applications in disease biomarker discovery. J. Chromatogr. B 2009, 877, 2836–2846. [Google Scholar] [CrossRef] [PubMed]
- Alhouayek, M.; Ameraoui, H.; Muccioli, G.G. Bioactive lipids in inflammatory bowel diseases–From pathophysiological alterations to therapeutic opportunities. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2021, 1866, 158854. [Google Scholar] [CrossRef]
- Barnig, C.; Bezema, T.; Calder, P.C.; Charloux, A.; Frossard, N.; Garssen, J.; Haworth, O.; Dilevskaya, K.; Levi-Schaffer, F.; Lonsdorfer, E.; et al. Activation of Resolution Pathways to Prevent and Fight Chronic Inflammation: Lessons from Asthma and Inflammatory Bowel Disease. Front. Immunol. 2019, 10, 1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhner, S.; Nagel, E.; Korber, J.; Vogelsang, H.; Linn, T.; Pichlmayr, R. Ileal and colonic fatty acid profiles in patients with active Crohn’s disease. Gut 1994, 35, 1424–1428. [Google Scholar] [CrossRef] [Green Version]
- Fernández–Bañares, F.; Esteve–Comas, M.; Mañé, J.; Navarro, E.; Bertrán, X.; Cabré, E.; Bartolí, R.; Boix, J.; Pastor, C.; Gassull, M.A. Changes in Mucosal Fatty Acid Profile in Inflammatorybowel Disease and in Experimental Colitis: A Common Response to Bowel Inflammation. Clin. Nutr. 1997, 16, 177–183. [Google Scholar] [CrossRef]
- Hwang, D.H.; Kim, J.; Lee, J.Y. Mechanisms for the activation of Toll-like receptor 2/4 by saturated fatty acids and inhibition by docosahexaenoic acid. Eur. J. Pharmacol. 2016, 785, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Kalenyak, K.; Heilmann, R.M.; Van De Lest, C.H.A.; Brouwers, J.F.; Burgener, I.A. Comparison of the systemic phospholipid profile in dogs diagnosed with idiopathic inflammatory bowel disease or food-responsive diarrhea before and after treatment. PLoS ONE 2019, 14, e0215435. [Google Scholar] [CrossRef] [PubMed]
- Ambrosini, Y.M.; Neuber, S.; Borcherding, D.; Seo, Y.-J.; Segarra, S.; Glanemann, B.; Garden, O.A.; Müller, U.; Adam, M.G.; Dang, V.; et al. Treatment with Hydrolyzed Diet Supplemented with Prebiotics and Glycosaminoglycans Alters Lipid Metabolism in Canine Inflammatory Bowel Disease. Front. Vet. Sci. 2020, 7, 451. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acar, N.; Berdeaux, O.; Grégoire, S.; Cabaret, S.; Martine, L.; Gain, P.; Thuret, G.; Creuzot-Garcher, C.P.; Bron, A.M.; Bretillon, L. Lipid Composition of the Human Eye: Are Red Blood Cells a Good Mirror of Retinal and Optic Nerve Fatty Acids? PLoS ONE 2012, 7, e35102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dushianthan, A.; Cusack, R.; Koster, G.; Grocott, M.P.W.; Postle, A.D. Insight into erythrocyte phospholipid molecular flux in healthy humans and in patients with acute respiratory distress syndrome. PLoS ONE 2019, 14, e0221595. [Google Scholar] [CrossRef] [PubMed]
- Arab, L. Biomarkers of Fat and Fatty Acid Intake. J. Nutr. 2003, 133 (Suppl. 3), 925S–932S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrows, B.R.; Parks, E.J. Contributions of Different Fatty Acid Sources to Very Low-Density Lipoprotein-Triacylglycerol in the Fasted and Fed States. J. Clin. Endocrinol. Metab. 2006, 91, 1446–1452. [Google Scholar] [CrossRef] [Green Version]
- Timlin, M.T.; Parks, E.J. Temporal pattern of de novo lipogenesis in the postprandial state in healthy men. Am. J. Clin. Nutr. 2005, 81, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Prasinou, P.; Crisi, P.E.; Chatgilialoglu, C.; Di Tommaso, M.; Sansone, A.; Gramenzi, A.; Belà, B.; De Santis, F.; Boari, A.; Ferreri, C. The Erythrocyte Membrane Lipidome of Healthy Dogs: Creating a Benchmark of Fatty Acid Distribution and Interval Values. Front. Vet. Sci. 2020, 7, 502. [Google Scholar] [CrossRef]
- Heilmann, R.M.; Berghoff, N.; Mansell, J.; Grützner, N.; Parnell, N.K.; Gurtner, C.; Suchodolski, J.S.; Steiner, J.M. Association of fecal calprotectin concentrations with disease severity, response to treatment, and other biomarkers in dogs with chronic inflammatory enteropathies. J. Vet. Intern. Med. 2018, 32, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Süß, R.; Teuber, K.; Eibisch, M.; Schiller, J. Lipid analysis by thin-layer chromatography—A review of the current state. J. Chromatogr. A 2011, 1218, 2754–2774. [Google Scholar] [CrossRef] [PubMed]
- Sansone, A.; Tolika, E.; Louka, M.; Sunda, V.; Deplano, S.; Melchiorre, M.; Anagnostopoulos, D.; Chatgilialoglu, C.; Formisano, C.; Di Micco, R.; et al. Hexadecenoic Fatty Acid Isomers in Human Blood Lipids and Their Relevance for the Interpretation of Lipidomic Profiles. PLoS ONE 2016, 11, e0152378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, H.J.; Chung, H.K.; Moon, J.; Shin, M.J. Relationship between the Estimates of Desaturase Activities and Cardiometabolic Phenotypes in Koreans. J Clin. Biochem. Nutr. 2011, 49, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanferlato, R.; Bortolotti, M.; Sansone, A.; Chatgilialoglu, C.; Polito, L.; De Spirito, M.; Maulucci, G.; Bolognesi, A.; Ferreri, C. Hexadecenoic Fatty Acid Positional Isomers and De Novo PUFA Synthesis in Colon Cancer Cells. Int. J. Mol. Sci. 2019, 20, 832. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, K.; Odahara, S.; Nakamura, M.; Koido, S.; Katahira, K.; Shiraishi, H.; Ohkusa, T.; Fujise, K.; Tajiri, H. The Fatty Acid Profile of the Erythrocyte Membrane in Initial-Onset Inflammatory Bowel Disease Patients. Dig. Dis. Sci. 2013, 58, 1235–1243. [Google Scholar] [CrossRef]
- Riezzo, G.; Ferreri, C.; Orlando, A.; Martulli, M.; D’Attoma, B.; Russo, F. Lipidomic analysis of fatty acids in erythrocytes of coeliac patients before and after a gluten-free diet intervention: A comparison with healthy subjects. Br. J. Nutr. 2014, 112, 1787–1796. [Google Scholar] [CrossRef] [Green Version]
- Ueda, Y.; Kawakami, Y.; Kunii, D.; Okada, H.; Azuma, M.; Le, D.S.N.; Yamamoto, S. Elevated concentrations of linoleic acid in erythrocyte membrane phospholipids in patients with inflammatory bowel disease. Nutr. Res. 2008, 28, 239–244. [Google Scholar] [CrossRef]
- Abbott, S.K.; Else, P.; Atkins, T.A.; Hulbert, A. Fatty acid composition of membrane bilayers: Importance of diet polyunsaturated fat balance. Biochim. Biophys. Acta (BBA) Biomembr. 2012, 1818, 1309–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baur, L.A.; O’Connor, J.; Pan, D.A.; Wu, B.; O’Connor, M.J.; Storlien, L.H. Relationships between the fatty acid composition of muscle and erythrocyte membrane phospholipid in young children and the effect of type of infant feeding. Lipids 2000, 35, 77–82. [Google Scholar] [CrossRef]
- Makrides, M.; Neumann, M.A.; Byard, R.; Simmer, K.; Gibson, R. Fatty acid composition of brain, retina, and erythrocytes in breast- and formula-fed infants. Am. J. Clin. Nutr. 1994, 60, 189–194. [Google Scholar] [CrossRef]
- Bueno-Hernández, N.; Dominguez-López, A.; Barreto-Zuñiga, R.; Sánchez-Muñoz, F.; Yamamoto-Furusho, J.K. Quantification of low expressed SCD1 gene in colonic mucosa from patients with active ulcerative colitis. Inflamm. Bowel Dis. 2011, 17, e155. [Google Scholar] [CrossRef]
- Chen, C.; Shah, Y.M.; Morimura, K.; Krausz, K.W.; Miyazaki, M.; Richardson, T.A.; Morgan, E.; Ntambi, J.M.; Idle, J.; Gonzalez, F.J. Metabolomics Reveals that Hepatic Stearoyl-CoA Desaturase 1 Downregulation Exacerbates Inflammation and Acute Colitis. Cell Metab. 2008, 7, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Gu, X.; Dai, W.; Ye, J.; Lu, F.; Chai, Y.; Fan, G.; Gonzalez, F.J.; Duan, G.; Qi, Y. A lipidomics investigation into the intervention of celastrol in experimental colitis. Mol. BioSyst. 2016, 12, 1436–1444. [Google Scholar] [CrossRef]
- Jezernik, G.; Potočnik, U. Comprehensive genetic study of fatty acids helps explain the role of noncoding inflammatory bowel disease associated SNPs and fatty acid metabolism in disease pathogenesis. Prostagland. Leukot. Essent. Fat. Acids 2018, 130, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, D.; Zhang, K.; Hung, C.; Glasgow, S.; Aruni, A.W.; Unternaehrer, J.; Payne, K.; Langridge, W.H.R.; De Leon, M. Palmitic acid is a toll-like receptor 4 ligand that induces human dendritic cell secretion of IL-1β. PLoS ONE 2017, 12, e0176793. [Google Scholar] [CrossRef]
- Schnyder, M.; Oevermann, A.; Doherr, M.G.; Luckschander, N.; Zurbriggen, A.; Burgener, I.A. Dysregulation of Toll-Like Receptors in Dogs with Chronic Enteropathies. J. Inflamm. Bowel Dis. Disord. 2018, 3, 1–9. [Google Scholar] [CrossRef]
- Burgener, I.; König, A.; Allenspach, K.; Sauter, S.; Boisclair, J.; Doherr, M.; Jungi, T. Upregulation of Toll-Like Receptors in Chronic Enteropathies in Dogs. J. Vet. Intern. Med. 2008, 22, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; Peritore, A.F.; Piras, C.; Cuzzocrea, S.; Crupi, R. Palmitoylethanolamide and Related ALIAmides: Prohomeostatic Lipid Compounds for Animal Health and Wellbeing. Vet. Sci. 2020, 7, 78. [Google Scholar] [CrossRef] [PubMed]
- Febo, E.; Crisi, P.E.; Oddi, S.; Pietra, M.; Galiazzo, G.; Piscitelli, F.; Gramenzi, A.; Di Prinzio, R.; Di Tommaso, M.; Bernabò, N.; et al. Circulating Endocannabinoids as Diagnostic Markers of Canine Chronic Enteropathies: A Pilot Study. Front. Vet. Sci. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Balvers, M.G.; Hendriks, H.F.; Wilpshaar, T.; Van Heek, T.; Witkamp, R.F.; Meijerink, J. Docosahexaenoyl serotonin emerges as most potent inhibitor of IL-17 and CCL-20 released by blood mononuclear cells from a series of N -acyl serotonins identified in human intestinal tissue. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2017, 1862, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Ito, Z.; Uchiyama, K.; Takami, S.; Koido, S.; Odahara, S.; Kubota, T.; Ohkusa, T. Utility of Fatty Acid Profile of Erythrocyte Membrane in Identifying Diagnostic Markers of Crohn’s Disease. Am. J. Gastroenterol. 2015, 110, S775. [Google Scholar] [CrossRef]
- Solakivi, T.; Kaukinen, K.; Kunnas, T.; Lehtimäki, T.; Mäki, M.; Nikkari, S.T. Serum fatty acid profile in celiac disease patients before and after a gluten-free diet. Scand. J. Gastroenterol. 2009, 44, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Chambrier, C.; Garcia, I.; Bannier, E.; Gerard-Boncompain, M.; Bouletreau, P. Specific changes in n -6 fatty acid metabolism in patients with chronic intestinal failure. Clin. Nutr. 2002, 21, 67–72. [Google Scholar] [CrossRef]
- Steel, D.M.; Ryd, W.; Ascher, H.; Strandvik, B. Abnormal Fatty Acid Pattern in Intestinal Mucosa of Children with Celiac Disease Is Not Reflected in Serum Phospholipids. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N.; Khalili, H.; Song, M.; Higuchi, L.M.; Lochhead, P.; Richter, J.M.; Chan, A.T. Genetic Polymorphisms in Fatty Acid Metabolism Modify the Association Between Dietary n3. Inflamm. Bowel Dis. 2017, 23, 1898–1904. [Google Scholar] [CrossRef] [PubMed]
- Costea, I.; Mack, D.R.; Lemaitre, R.N.; Israel, D.; Marcil, V.; Ahmad, A.; Amre, D.K. Interactions Between the Dietary Polyunsaturated Fatty Acid Ratio and Genetic Factors Determine Susceptibility to Pediatric Crohn’s Disease. Gastroenterology 2014, 146, 929–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costea, I.; Mack, D.R.; Israel, D.; Morgan, K.; Krupoves, A.; Seidman, E.; Deslandres, C.; Lambrette, P.; Grimard, G.; Levy, E.; et al. Genes Involved in the Metabolism of Poly-Unsaturated Fatty-Acids (PUFA) and Risk for Crohn’s Disease in Children & Young Adults. PLoS ONE 2010, 5, e15672. [Google Scholar] [CrossRef]
- Marion-Letellier, R.; Savoye, G.; Beck, P.L.; Panaccione, R.; Ghosh, S. Polyunsaturated Fatty Acids in Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2013, 19, 650–661. [Google Scholar] [CrossRef]
- Trebble, T.M.; Arden, N.K.; Wootton, S.; Calder, P.; Mullee, M.A.; Fine, D.; Stroud, M.A. Fish oil and antioxidants alter the composition and function of circulating mononuclear cells in Crohn disease. Am. J. Clin. Nutr. 2004, 80, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Esteve, M.; Ramírez, M.; Fernández-Bañares, F.; Lacruz, A.A.; Gil, A.; Cabre, E.; Gonzalez-Huix, F.; Moreno, J.; Humbert, P.; Guilera, M. Plasma polyunsaturated fatty acid pattern in active inflammatory bowel disease. Gut 1992, 33, 1365–1369. [Google Scholar] [CrossRef] [Green Version]
- Esteve, M.; Núñez, M.C.; Fernández-Bañares, F.; Abad-Lacruz, A.; Gil, A.; Cabre, E.; Gonzalez-Huix, F.; Bertrán, X.; Gassull, M. Àngel Abnormal plasma polyunsaturated fatty acid pattern in non-active inflammatory bowel disease. Gut 1993, 34, 1370–1373. [Google Scholar] [CrossRef] [Green Version]
- Daniluk, U.; Daniluk, J.; Kucharski, R.; Kowalczyk, T.; Pietrowska, K.; Samczuk, P.; Filimoniuk, A.; Kretowski, A.; Lebensztejn, D.; Ciborowski, M. Untargeted Metabolomics and Inflammatory Markers Profiling in Children With Crohn’s Disease and Ulcerative Colitis—A Preliminary Study. Inflamm. Bowel Dis. 2019, 25, 1120–1128. [Google Scholar] [CrossRef]
- Scoville, E.; Allaman, M.M.; Adams, D.W.; Motley, A.K.; Peyton, S.C.; Ferguson, S.L.; Horst, S.N.; Williams, C.; Beaulieu, D.B.; Schwartz, D.A.; et al. Serum Polyunsaturated Fatty Acids Correlate with Serum Cytokines and Clinical Disease Activity in Crohn’s Disease. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Wiese, D.M.; Horst, S.N.; Brown, C.T.; Allaman, M.M.; Hodges, M.E.; Slaughter, J.; Druce, J.P.; Beaulieu, D.B.; Schwartz, D.A.; Wilson, K.T.; et al. Serum Fatty Acids Are Correlated with Inflammatory Cytokines in Ulcerative Colitis. PLoS ONE 2016, 11, e0156387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergeant, S.; Rahbar, E.; Chilton, F.H. Gamma-linolenic acid, Dihommo-gamma linolenic, Eicosanoids and Inflammatory Processes. Eur. J. Pharmacol. 2016, 785, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Burr, G.O.; Burr, M.M. A new deficiency disease produced by the rigid exclusion of fat from the diet. J. Biol. Chem. 1929, 82, 345–367. [Google Scholar] [CrossRef]
- Revskij, D.; Haubold, S.; Viergutz, T.; Kröger-Koch, C.; Tuchscherer, A.; Kienberger, H.; Rychlik, M.; Tröscher, A.; Hammon, H.M.; Schuberth, H.-J.; et al. Dietary Fatty Acids Affect Red Blood Cell Membrane Composition and Red Blood Cell ATP Release in Dairy Cows. Int. J. Mol. Sci. 2019, 20, 2769. [Google Scholar] [CrossRef] [Green Version]
- Perona, J.S. Membrane lipid alterations in the metabolic syndrome and the role of dietary oils. Biochim. Biophys. Acta (BBA) Biomembr. 2017, 1859, 1690–1703. [Google Scholar] [CrossRef] [PubMed]
- Symons, J.; Cho, K.-J.; Chang, J.T.; Du, G.; Waxham, M.N.; Hancock, J.F.; Levental, I.; Levental, K.R. Lipidomic atlas of mammalian cell membranes reveals hierarchical variation induced by culture conditions, subcellular membranes, and cell lineages. Soft Matter 2021, 17, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R.; Portero–Otín, M.; Riba, D.; Ruiz, C.; Prat, J.; Bellmunt, M.J.; Barja, G. Mitochondrial Membrane Peroxidizability Index Is Inversely Related to Maximum Life Span in Mammals. J. Lipid Res. 1998, 39, 1989–1994. [Google Scholar] [CrossRef]
- Rubio, C.; Martinez-Subiela, S.; Ruiz, J.H.; Tvarijonaviciute, A.; Cerón, J.; Allenspach, K. Serum biomarkers of oxidative stress in dogs with idiopathic inflammatory bowel disease. Vet. J. 2017, 221, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.; Ye, J.; Zhou, Z.; Ke, X.; Yu, X.; Huang, K. Fatty Acid Metabolism in Immune Cells: A Bioinformatics Analysis of Genes Involved in Ulcerative Colitis. DNA Cell Biol. 2020, 39, 1573–1582. [Google Scholar] [CrossRef]
- Robichaud, P.-P.; Munganyiki, J.E.; Boilard, E.; Surette, M.E. Polyunsaturated fatty acid elongation and desaturation in activated human T-cells: ELOVL5 is the key elongase. J. Lipid Res. 2018, 59, 2383–2396. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N. Folic acid and polyunsaturated fatty acids improve cognitive function and prevent depression, dementia, and Alzheimer’s disease—But how and why? Prostagland. Leukot. Essent. Fat. Acids 2008, 78, 11–19. [Google Scholar] [CrossRef]
- Ued, F.V.; Mathias, M.G.; Toffano, R.B.D.; Barros, T.T.; Almada, M.O.R.V.; Salomão, R.G.; Coelho-Landell, C.A.; Hillesheim, E.; Camarneiro, J.M.; Camelo-Junior, J.S.; et al. Vitamin B2 and Folate Concentrations are Associated with ARA, EPA and DHA Fatty Acids in Red Blood Cells of Brazilian Children and Adolescents. Nutrition 2019, 11, 2918. [Google Scholar] [CrossRef] [Green Version]
- Umhau, J.C.; Dauphinais, K.M.; Patel, S.H.; Nahrwold, D.A.; Hibbeln, J.R.; Rawlings, R.R.; George, D.T. The relationship between folate and docosahexaenoic acid in men. Eur. J. Clin. Nutr. 2005, 60, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Selhub, J. Folate, vitamin B12 and vitamin B6 and one carbon metabolism. J. Nutr. Heal. Aging 2002, 6, 39–42. [Google Scholar]
- Ferreri, C.; Grabovskiy, S.A.; Aoun, M.; Melchiorre, M.; Kabal’Nova, N.; Feillet-Coudray, C.; Fouret, G.; Coudray, C.; Chatgilialoglu, C. Trans Fatty Acids: Chemical Synthesis of Eicosapentaenoic Acid Isomers and Detection in Rats Fed a Deodorized Fish Oil Diet. Chem. Res. Toxicol. 2012, 25, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Menounou, G.; Giacometti, G.; Scanferlato, R.; Dambruoso, P.; Sansone, A.; Tueros, I.; Amézaga, J.; Chatgilialoglu, C.; Ferreri, C. Trans Lipid Library: Synthesis of Docosahexaenoic Acid (DHA) Monotrans Isomers and Regioisomer Identification in DHA-Containing Supplements. Chem. Res. Toxicol. 2018, 31, 191–200. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | Healthy Dogs (n = 68) | Chronic Enteropathy (n = 48) | p Value |
|---|---|---|---|
| Males | 30 | 31 | 0.04 |
| Females | 38 | 17 | |
| Age | 41 (2–156) | 46 (4–144) | 0.83 |
| Bodyweight | 22.0 (2.6–43.0) | 22.0 (4.0–36.3) | 0.43 |
| Variable | FRE (n = 28) | ARE (n = 5) | IRE/NRE (n = 15) | p Value |
|---|---|---|---|---|
| BCS (1–9) | 5 (2–6) | 3 (2–5) | 4 (1–7) | 0.175 |
| CCECAI | 3 (1–12) | 6 (5–13) | 6 (2–19) | 0.003 * |
| CRP (0–1 mg/dL) | 0.6 (0.0–12.4) | 1.0 (0.9–2.3) | 1.0 (0.0–10.0) | 0.340 |
| Folate (6.5–11.5 µg/L) | 8.9 (2.2–24.0) | 7.9 (2.6–11.0) | 7.7 (1.0–24.0) | 0.891 |
| Cobalamin (250–730 ng/L) | 397 (150–824) | 284 (242–575) | 307 (150–994) | 0.356 |
| Variable | Healthy Dogs (n = 68) | Chronic Enteropathy (n = 48) | |
|---|---|---|---|
| Interquartile (IQ) Range | Number (Percentage) of Dogs with Values above the IQ Range | Number (Percentage) of Dogs with Values below the IQ Range | |
| Palmitic Acid | 13.03–17.43 | 0 (0%) | 42 (87.5%) |
| Stearic Acid | 18.95–21.81 | 41 (85.4%) | 0 (0%) |
| Saturated Fatty Acids | 33.22–37.44 | 4 (8.3%) | 11 (22.9%) |
| Palmitoleic Acid | 0.19–0.33 | 24 (50%) | 11 (22.9%) |
| Oleic Acid | 8.50–10.16 | 14 (29.1%) | 8 (16.6%) |
| Vaccenic Acid | 1.77–2.28 | 18 (37.5%) | 11 (22.9%) |
| Monounsaturated Fatty Acids | 10.43–12.72 | 17 (35.4%) | 5 (10.4%) |
| Linoleic Acid | 13.15–16.22 | 6 (12.5%) | 25 (52.0%) |
| Dihomo-gamma-linolenic Acid | 1.06–1.57 | 28 (58.3%) | 4 (8.3%) |
| Arachidonic Acid | 31.43–37.64 | 10 (20.8%) | 2 (4.1%) |
| ω-6 Polyunsaturated Fatty Acids | 47.27–53.20 | 12 (25.0%) | 7 (14.5%) |
| EPA | 0.46–0.89 | 22 (45.8%) | 11 (22.9%) |
| DHA | 0.70–1.68 | 14 (29.1%) | 4 (8.3%) |
| ω-3 Polyunsaturated Fatty Acids | 1.20–2.40 | 21 (43.7%) | 5 (10.4%) |
| Polyunsaturated Fatty Acids | 49.97–55.29 | 12 (25.0%) | 7 (14.5%) |
| SFA/MUFA | 2.83–3.28 | 8 (16.6%) | 25 (52.0%) |
| ω-6/ω-3 | 19.60–40.18 | 4 (8.3%) | 14 (29.1%) |
| PUFA Balance | 2.42–4.85 | 14 (29.1%) | 4 (8.3%) |
| Unsaturation Index | 186.8–207.2 | 14 (29.1%) | 4 (8.3%) |
| Peroxidation Index | 160.7–181.9 | 17 (35.4%) | 5 (10.4%) |
| Elongase-6 activity | 0.56–0.71 | 43 (89.5%) | 0 (0.0%) |
| Delta-6 desaturase activity | 0.07–0.11 | 32 (66.6%) | 3 (6.2%) |
| Delta-5 desaturase activity | 21.09–30.51 | 7 (14.5%) | 18 (37.5%) |
| Delta-9 desaturase activity | 0.40–0.51 | 4 (8.3%) | 20 (41.6%) |
| Fatty Acids | BCS | CCECAI | CRP | Folate | Cobalamin |
|---|---|---|---|---|---|
| Palmitic Acid | 0.029 | 0.051 | 0.313 | −0.119 | −0.128 |
| Stearic Acid | 0.092 | 0.222 | −0.018 | −0.138 | −0.035 |
| Palmitoleic Acid | 0.095 | 0.031 | 0.075 | 0.107 | −0.020 |
| Oleic Acid | −0.031 | 0.245 | 0.183 | −0.355 * | −0.148 |
| Vaccenic Acid | −0.044 | 0.223 | 0.222 | −0.347 * | −0.362 * |
| Linoleic Acid | −0.165 | −0.144 | −0.013 | 0.177 | 0.125 |
| Dihomo-gamma-linolenic Acid | −0.103 | 0.021 | −0.067 | −0.241 | −0.206 |
| Arachidonic Acid | −0.004 | −0.186 | −0.207 | 0.309 | 0.166 |
| EPA | 0.06 | −0.241 | −0.194 | 0.142 | 0.080 |
| DHA | 0.125 | −0.011 | −0.101 | 0.147 | −0.108 |
| Fatty Acid Families | |||||
| Total SFAs | 0.126 | 0.195 | 0.193 | −0.234 | −0.013 |
| Total MUFAs | −0.022 | 0.222 | 0.146 | −0.365 * | −0.204 |
| ω-6 Polyunsaturated Fatty Acids | −0.097 | −0.300 * | −0.229 | 0.330 * | 0.181 |
| ω-3 Polyunsaturated Fatty Acids | 0.134 | −0.185 | −0.079 | 0.171 | −0.032 |
| Total PUFAs | −0.077 | −0.283 | −0.276 | 0.369 * | 0.128 |
| Homeostasis Indexes | |||||
| SFA/MUFA | 0.097 | −0.116 | −0.0114 | 0.249 | 0.116 |
| ω-6/ω-3 | −0.125 | 0.142 | 0.0578 | −0.112 | 0.069 |
| PUFA Balance | 0.125 | −0.142 | −0.0578 | 0.112 | −0.069 |
| Unsaturation Index | −0.01 | −0.250 | −0.275 | 0.317 | 0.106 |
| Peroxidation Index | 0.064 | −0.250 | −0.232 | 0.319 | 0.087 |
| Enzyme indexes | |||||
| Elongase-6 activity | −0.04 | 0.081 | −0.147 | −0.099 | 0.021 |
| Delta-6 desaturase activity | 0.001 | 0.107 | −0.074 | −0.340 * | −0.358 * |
| Delta-5 desaturase activity | 0.087 | −0.042 | 0.043 | 0.327 * | 0.235 |
| Delta-9 desaturase activity | −0.059 | 0.026 | 0.114 | −0.128 | −0.034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crisi, P.E.; Luciani, A.; Di Tommaso, M.; Prasinou, P.; De Santis, F.; Chatgilialoglu, C.; Pietra, M.; Procoli, F.; Sansone, A.; Giordano, M.V.; et al. The Fatty Acid-Based Erythrocyte Membrane Lipidome in Dogs with Chronic Enteropathy. Animals 2021, 11, 2604. https://doi.org/10.3390/ani11092604
Crisi PE, Luciani A, Di Tommaso M, Prasinou P, De Santis F, Chatgilialoglu C, Pietra M, Procoli F, Sansone A, Giordano MV, et al. The Fatty Acid-Based Erythrocyte Membrane Lipidome in Dogs with Chronic Enteropathy. Animals. 2021; 11(9):2604. https://doi.org/10.3390/ani11092604
Chicago/Turabian StyleCrisi, Paolo Emidio, Alessia Luciani, Morena Di Tommaso, Paraskevi Prasinou, Francesca De Santis, Chryssostomos Chatgilialoglu, Marco Pietra, Fabio Procoli, Anna Sansone, Maria Veronica Giordano, and et al. 2021. "The Fatty Acid-Based Erythrocyte Membrane Lipidome in Dogs with Chronic Enteropathy" Animals 11, no. 9: 2604. https://doi.org/10.3390/ani11092604
APA StyleCrisi, P. E., Luciani, A., Di Tommaso, M., Prasinou, P., De Santis, F., Chatgilialoglu, C., Pietra, M., Procoli, F., Sansone, A., Giordano, M. V., Gramenzi, A., Ferreri, C., & Boari, A. (2021). The Fatty Acid-Based Erythrocyte Membrane Lipidome in Dogs with Chronic Enteropathy. Animals, 11(9), 2604. https://doi.org/10.3390/ani11092604