
Heads and Tails: An Analysis of Visual Signals in Cats, Felis catus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Housing, Subjects, and Methods
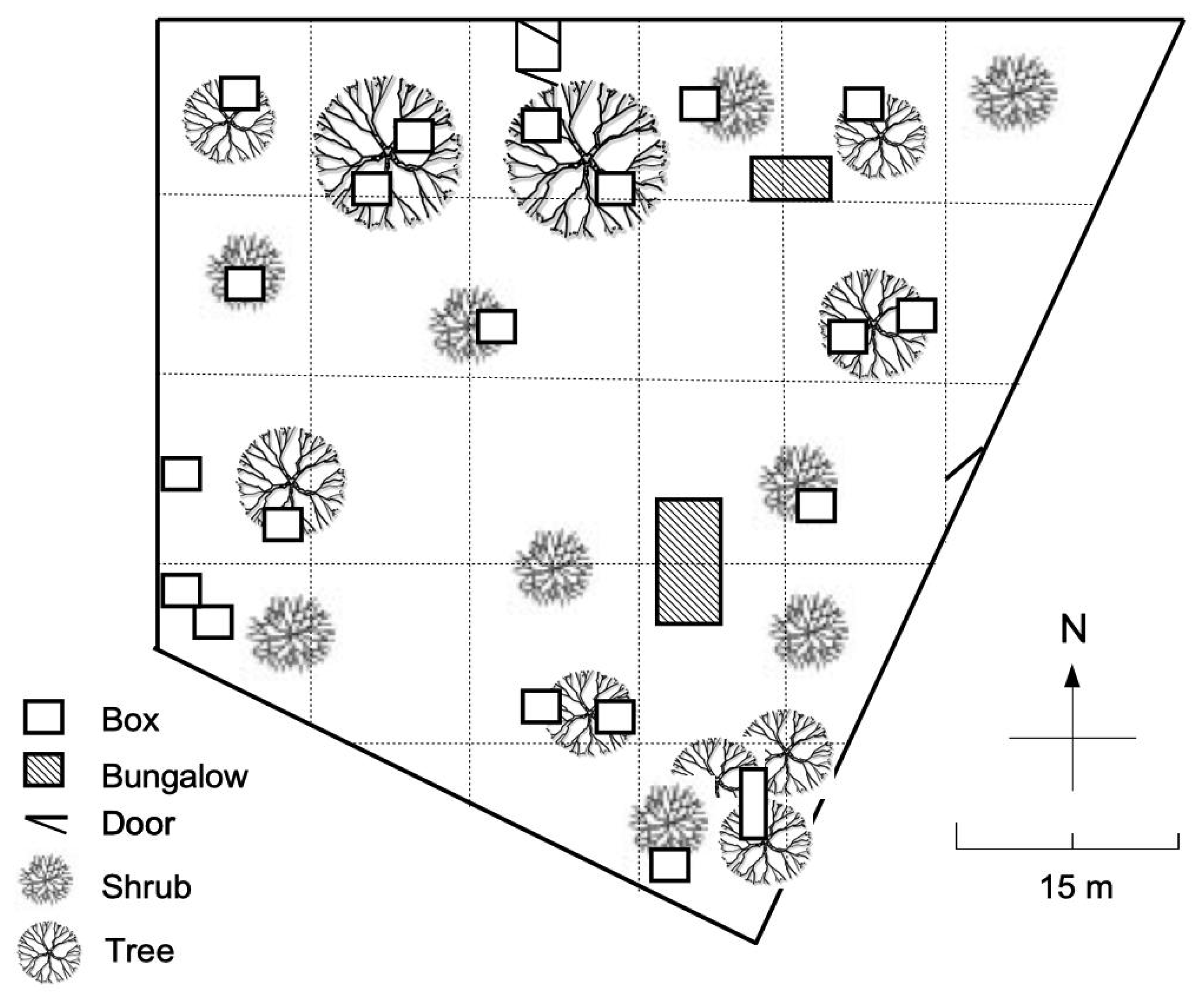
2.1. Housing and Subjects
2.1.1. Ethical Note
2.1.2. Subjects
2.2. Methods
2.2.1. Observations
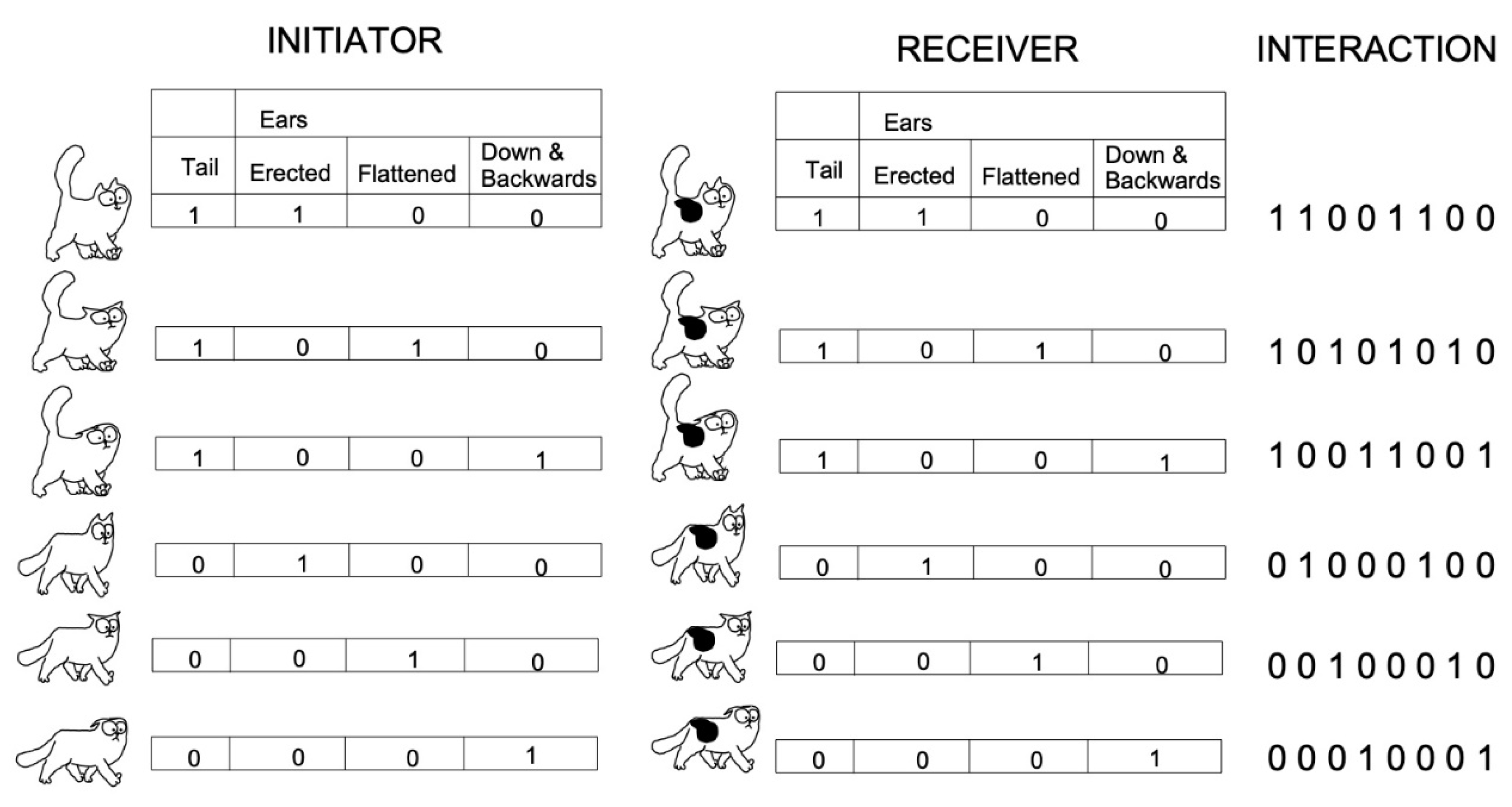
2.2.2. Variables
2.2.3. Coding Methodology
2.3. Data Processing and Analyses
3. Results
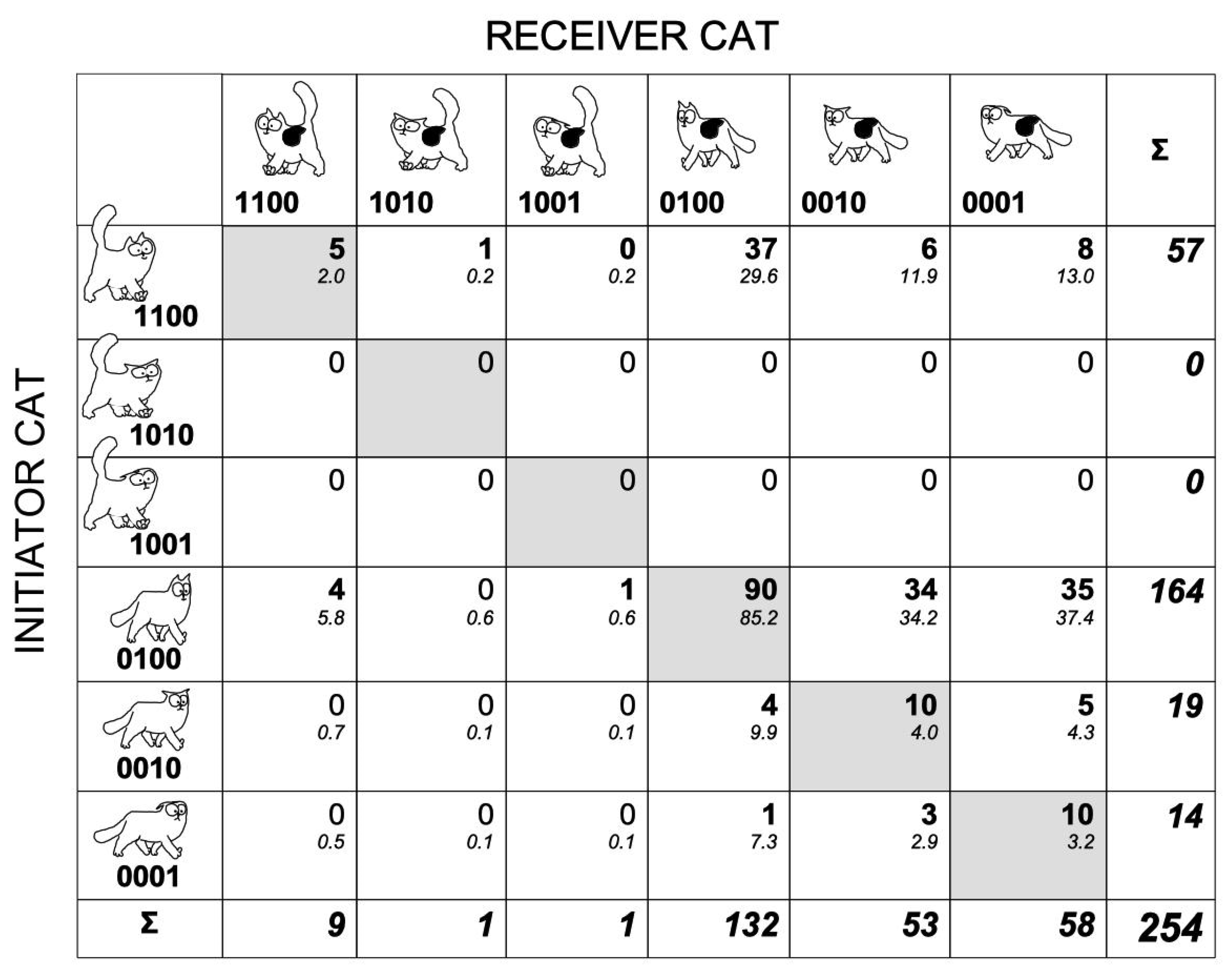
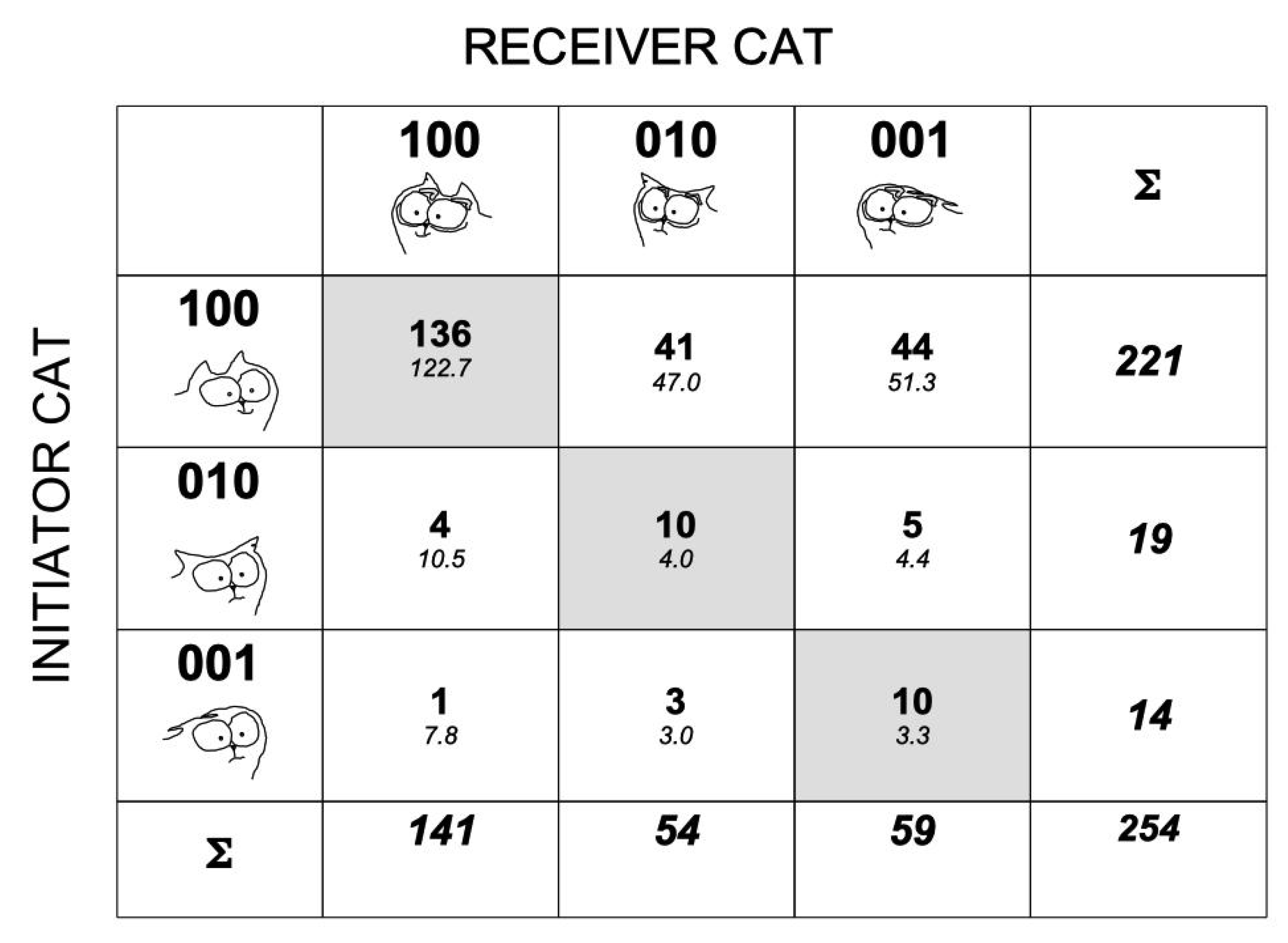
3.1. Configurations Expressed during Cat–Cat Interactions
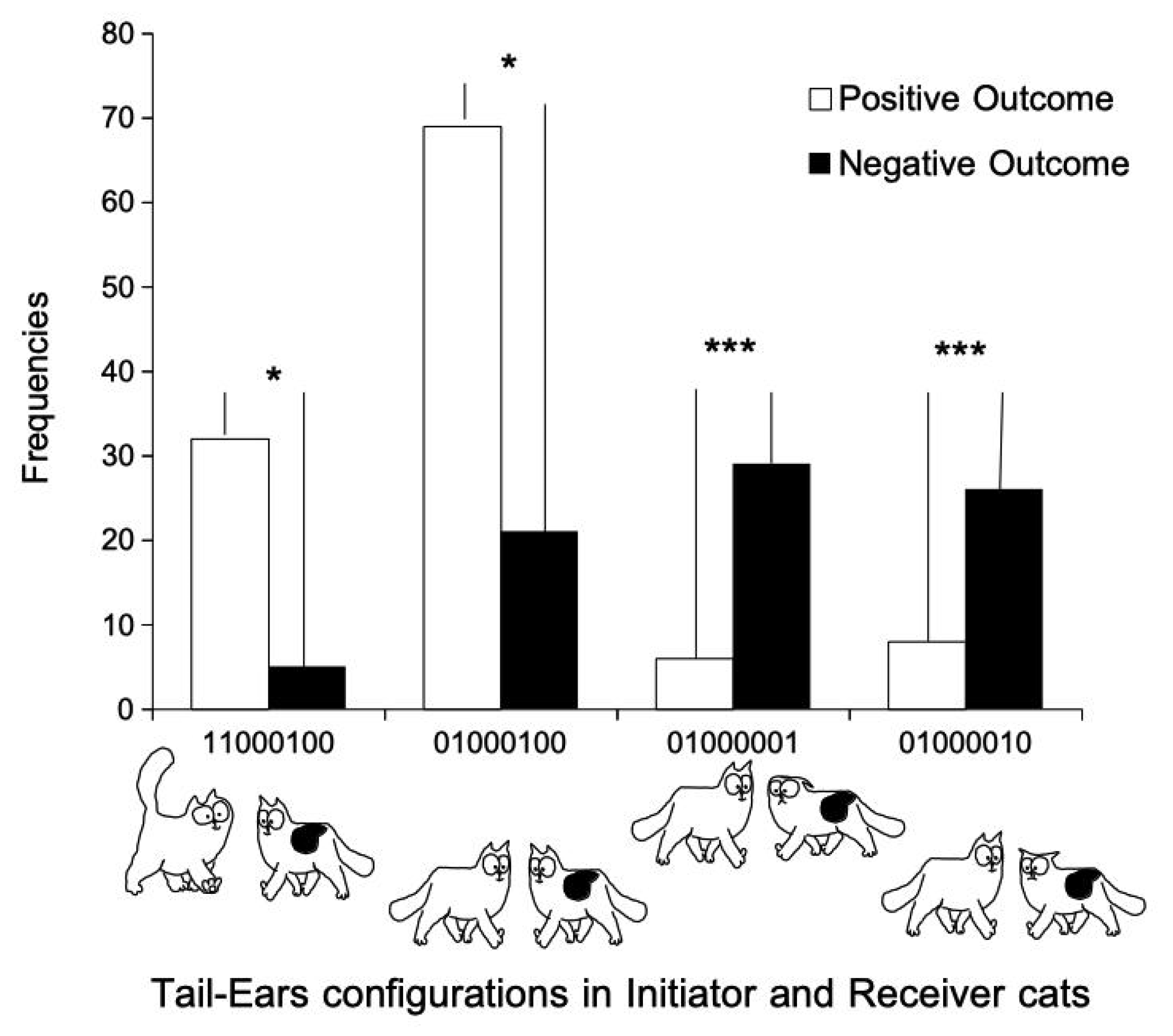
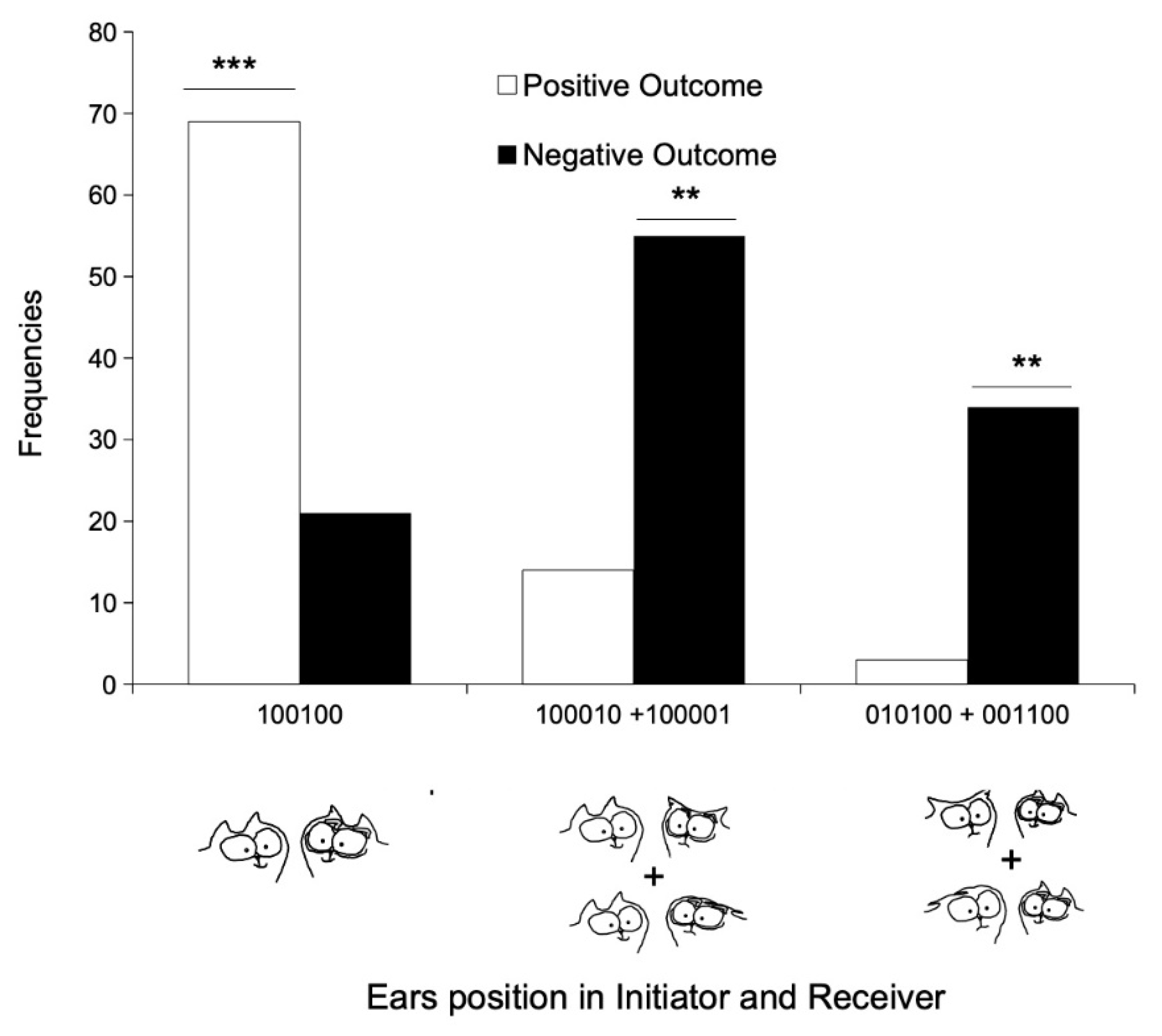
3.2. Tail–Ears Configurations at the Onset of Interactions and Outcomes of These Interactions
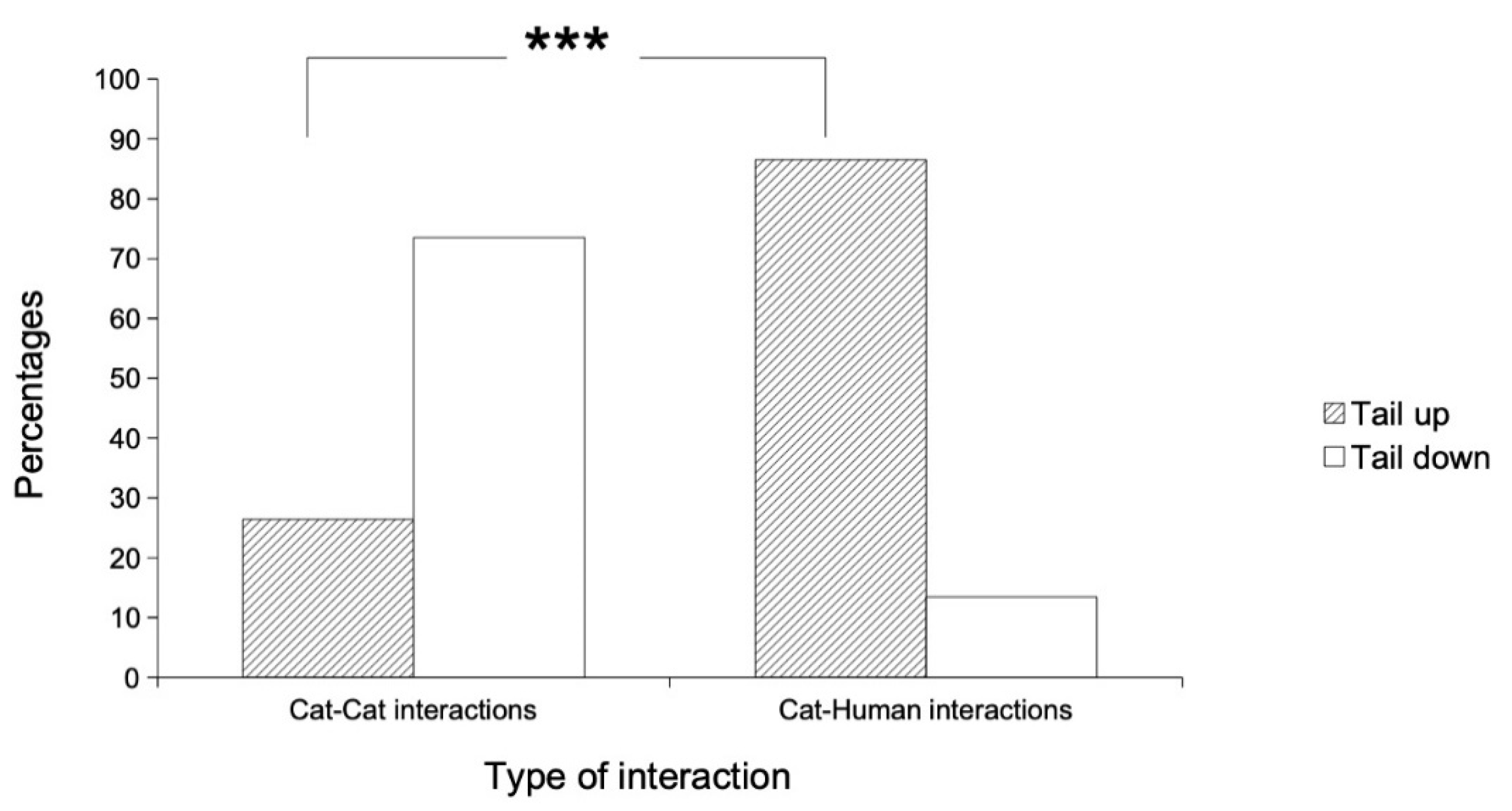
3.3. Tail Position According to Intra- and Interspecific Approaches
4. Discussion
4.1. Welfare Issues
4.2. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marler, P. Aggregation and dispersal: Two functions in primates communication. In Primates: Studies in Adaptation and Variability.; Jay, P.C., Ed.; Holt, Rinehart & Winston: New York, NY, USA, 1968; pp. 420–438. [Google Scholar]
- Menzel, E.W., Jr.; Johnson, M.K. Communication and cognitive organization in humans and other animals. Ann. N. Y. Acad. Sci. 1976, 280, 131–142. [Google Scholar] [CrossRef]
- Tinbergen, N. Derived activities; their causation, biological significance, origin, and emancipation during evolution. Quart. Rev. Biol. 1952, 27, 1–32. [Google Scholar] [CrossRef]
- Tinbergen, N. Social releasers and the experimental method required for their study. Wilson Bull. 1948, 60, 6–51. [Google Scholar]
- Golani, I. Homeostatic motor processes in mammalian interactions: A choreography of display. In Perspectives in Ethology; Bateson, P.P.G., Klopfer, P.H., Eds.; Plenum Press: New York, NY, USA, 1976; Volume 2, pp. 69–134. [Google Scholar]
- Green, S. Variation of vocal pattern with social situation in the Japanese monkey (Macaca fuscata): A field study. Primate Behav. 1975, 4, 1–102. [Google Scholar]
- Partan, S.; Marler, P. Communication goes multimodal. Science 1999, 283, 1272–1273. [Google Scholar] [CrossRef]
- Baraud, I.; Pierre, J.-S.; Deputte, B.L.; Blois-Heulin, C. Informative value of vocalizations during multimodal interactions in red-capped mangabeys (Cercocebus torquatus torquatus). In Discovering Hidden Temporal Patterns in Behaviour and Interaction; T-pattern Detection and Analysis with THEME™; Magnusson, M., Burgoon, J.K., Csaarrubea, M., Eds.; Springer: Berlin, Germany, 2016; pp. 255–277. [Google Scholar]
- Kiley-Worthington, M. The tail movements of ungulates, canids, and felids with particular reference to their causation and function as displays. Behaviour 1975, 46, 69–115. [Google Scholar] [CrossRef]
- Hausfater, G. Tail carriage in baboons (Papio cynocephalus). Folia Primatol. 1977, 27, 41–59. [Google Scholar] [CrossRef]
- Schenkel, R. Ausdrucks-Studien an Wolfen. Behave 1947, 1, 81–129. [Google Scholar] [CrossRef]
- Harrington, F.; Asa, C. Wolf communication. In Wolves: Behavior, Ecology, and Conservation; Mech, L., Boitani, L., Eds.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 66–103. [Google Scholar]
- Lisberg, A.E.; Snowdon, C.T. The effects of sex, gonadectomy and status on investigation patterns of unfamiliar conspecific urine in domestic dogs. Canis Fam. Anim. Behav. 2009, 77, 1147–1154. [Google Scholar] [CrossRef]
- Ekman, P.; Friesen, W.V. Facial Action Coding System: A Technique for the Measurement of Facial Movement; Consulting Psychologists Press: Palo Alto, CA, USA, 1978. [Google Scholar]
- Marler, P.; Tenaza, T. Signalling behavior of apes with special reference to vocalization. In How Animals Communicate; Sebeok, T.A., Ed.; Indiana University Press: Bloomington, IN, USA, 1977; Chapter 36; pp. 965–1033. [Google Scholar]
- Parr, L.A.; Waller, B.M.; Burrows, A.M.; Gothard, K.M.; Vick, S.J. MaqFACS: A muscle-based facial movement coding system for the macaque monkey. Am. J. Phys. Anthropol. 2010, 143, 625–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalla Costa, E.; Minero, M.; Lebelt, D.; Stucke, D.; Canali, E.; Leach, M.C. Development of the horse grimace scale (HGS) as a pain assessment tool in horses undergoing routine castration. PLoS ONE 2014, 9, e92281. [Google Scholar] [CrossRef] [Green Version]
- Dolensek, N.; Gehrlach, D.A.; Klein, A.S.; Gogolla, N. Facial expressions of emotion states and their neuronal correlates in mice. Science 2020, 368, 89–94. [Google Scholar] [CrossRef]
- Langford, D.J.; Bailey, A.L.; Chanda, M.L.; Clarke, S.E.; Drummond, T.E.; Echols, S. Coding of facial expressions of pain in the laboratory mouse. Nat. Methods 2010, 7, 447–449. [Google Scholar] [CrossRef]
- Fox, W. Behaviour of Wolves, Dogs and Related Canids; Jonathan Cape: London, UK, 1971. [Google Scholar]
- Waring, G.H. Horse Behavior; Noyes Publications/William Andrew Publishing: Norwich, NY, USA, 2003; 442p. [Google Scholar]
- Rosenblatt, J.S.; Schneirla, T.C. The behaviour of cats. In The Behaviour of Domestic Animals; Hafez, E.S.E., Ed.; Baillière Tindall & Cox: London, UK, 1962; Chapter 16; pp. 453–488. [Google Scholar]
- Leyhausen, P. Cat Behavior: The Predatory and Social Behavior of Domestic and Wild Cats; Garland STPM Press: New York, NY, USA, 1979; 340p. [Google Scholar]
- Macdonald, D.W. The ecology of carnivore social behaviour. Nature 1983, 301, 379–384. [Google Scholar] [CrossRef]
- Kerby, G.; Macdonald, D.W. Cat society and the consequences of colony size. In The Domestic Cat: The Biology of Its Behavior, 1st ed.; Turner, D.C., Bateson, P., Eds.; Cambridge University Press: Cambridge, UK, 1988; Chapter 6; pp. 67–81. [Google Scholar]
- Bradshaw, J.W.S. Sociality in cats: A comparative review. J. Vet. Behav. 2016, 11, 113–124. [Google Scholar] [CrossRef]
- Darwin, C. The Expression of the Emotions in Man and Animals, 3rd ed.; Oxford University Press: Oxford, UK, 1998; 372p. [Google Scholar]
- Bennett, V.; Gourkow, N.; Mills, D.S. Facial correlates of emotional behaviour in the domestic cat (Felis catus). Behav. Proc. 2017, 141, 342–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caeiro, C.C.; Burrows, A.M.; Waller, B.M. Development and application of CatFACS: Are human cat adopters influenced by cat facial expressions? Appl. Anim. Behav. Sci. 2017, 189, 66–78. [Google Scholar] [CrossRef]
- Evangelista, M.C.; Watanabe, R.; Leung, V.S.Y.; Monteiro, B.P.; O’Toole, E.; Pang, D.S.J.; Steagall, P.V. Facial expressions of pain in cats: The development and validation of a feline grimace scale. Sci. Rep. 2019, 9, 19128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finka, L.R.; Luna, S.P.; Brondani, J.T.; Tzimiropoulos, Y.; McDonagh, J.; Farnworth, M.J.; Ruta, M.; Mills, D.S. Geometric morphometrics for the study of facial expressions in nonhuman animals, using the domestic cat as an exemplar. Sci. Rep. 2019, 38, 9883. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.; Cameron-Beaumont, C. The signalling repertoire of the domestic cat and its undomesticated relatives. In The Domestic Cat: The Biology of Its Behavior, 2nd ed.; Turner, D.C., Bateson, P., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 67–93. [Google Scholar]
- Cafazzo, S.; Natoli, E. The social function of tail up in the domestic cat (Felis silvestris catus). Behav. Process. 2009, 80, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Wedl, M.; Bauera, B.; Gracey, D.; Grabmayer, C.; Spielauer, E.; Day, J.; Kotrschal, K. Factors influencing the temporal patterns of dyadic behaviours and interactions between domestic cats and their owners. Behav. Process. 2011, 86, 58–67. [Google Scholar] [CrossRef]
- Cameron-Beaumont, C.L. Visual and Tactile Communication in the Domestic Cat (Felis silvestris catus) and Undomesticated Small Felids. Ph.D. Thesis, University of Southampton, Southampton, UK, 1997. [Google Scholar]
- Tinbergen, N. The Study of Instinct; Oxford University Press: New York, NY, USA, 1951; 228p. [Google Scholar]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–265. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W. H. Freeman: San Francisco, CA, USA, 1981; p. 887. [Google Scholar]
- Siegel, S.; Castellan, N.J. Non Parametric Statistics for the Behavioral Sciences; McGraw-Hill Kogakusha: Tokyo, Japan, 1988; 399p. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 1st ed.; W. H. Freeman: San Francisco, CA, USA, 1969; 887p. [Google Scholar]
- Leyhausen, P. The communal organization of solitary mammals. Symp. Zool. Soci. London 1965, 14, 249–263. [Google Scholar]
- Barone, R. Anatomie Comparée des Manmifères Domestiques; Tome Vigot: Paris, France, 1989; 984p. [Google Scholar]
- Boissy, A.; Aubert, A.; Désiré, L.; Greiveldinger, L.; Delval, E.; Veissier, I. Cognitive sciences to relate ear postures to emotions in sheep. Anim. Welf. 2011, 20, 47–56. [Google Scholar]
- Correia-Caeiro, C.; Guo, K.; Mills, D.S. Bodily emotional expressions are a primary source of information for dogs, but not for humans. Anim. Cogn. 2021, 24, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, I.S. Dominance: The baby and the bathwater. Behav. Brain Sci. 1981, 4, 419–429. [Google Scholar] [CrossRef]
- Bradshaw, J.W.S.; Blackwell, E.J.; Casey, R.A. Dominance in domestic dogs—Useful construct or bad habit? J. Vet. Behav. 2009, 4, 135–144. [Google Scholar] [CrossRef]
- Bouissou, M.F. Observations sur la hiérarchie sociale chez les bovins domestiques. Ann. Biol. Anim. Bioch. Biophys. 1965, 5, 327–339. [Google Scholar] [CrossRef]
- Bateson, P.; Turner, D.C. Question about cats. In The Domestic Cat: The Biology of Its Behavior, 2nd ed.; Turner, D.C., Bateson, P., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 230–237. [Google Scholar]
- Budiansky, S. The Character of Cats; Viking: New York, NY, USA, 2002; 227p. [Google Scholar]
- Bradshaw, J.W.S.; Nott, H.M.R. Social and communication behaviour of companion dogs. In The Domestic Dog: Its Evolution, Behaviour and Interactions with People; Serpell, J., Ed.; Cambridge University Press: Cambridge, UK, 1995; Chapter 8; pp. 115–1130. [Google Scholar]
- Tavernier, C.; Ahmed, S.; Houpt, K.A.; Yeon, S.C. Feline vocal communication. J. Vet. Sci. 2020, 21, e18. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deputte, B.L.; Jumelet, E.; Gilbert, C.; Titeux, E. Heads and Tails: An Analysis of Visual Signals in Cats, Felis catus. Animals 2021, 11, 2752. https://doi.org/10.3390/ani11092752
Deputte BL, Jumelet E, Gilbert C, Titeux E. Heads and Tails: An Analysis of Visual Signals in Cats, Felis catus. Animals. 2021; 11(9):2752. https://doi.org/10.3390/ani11092752
Chicago/Turabian StyleDeputte, Bertrand L., Estelle Jumelet, Caroline Gilbert, and Emmanuelle Titeux. 2021. "Heads and Tails: An Analysis of Visual Signals in Cats, Felis catus" Animals 11, no. 9: 2752. https://doi.org/10.3390/ani11092752
APA StyleDeputte, B. L., Jumelet, E., Gilbert, C., & Titeux, E. (2021). Heads and Tails: An Analysis of Visual Signals in Cats, Felis catus. Animals, 11(9), 2752. https://doi.org/10.3390/ani11092752