
Some Aspects of Development and Histological Structure of the Visual System of Nothobranchius Guentheri

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Object of Study
2.2. Embryology
2.3. Histology
2.4. Microscopy
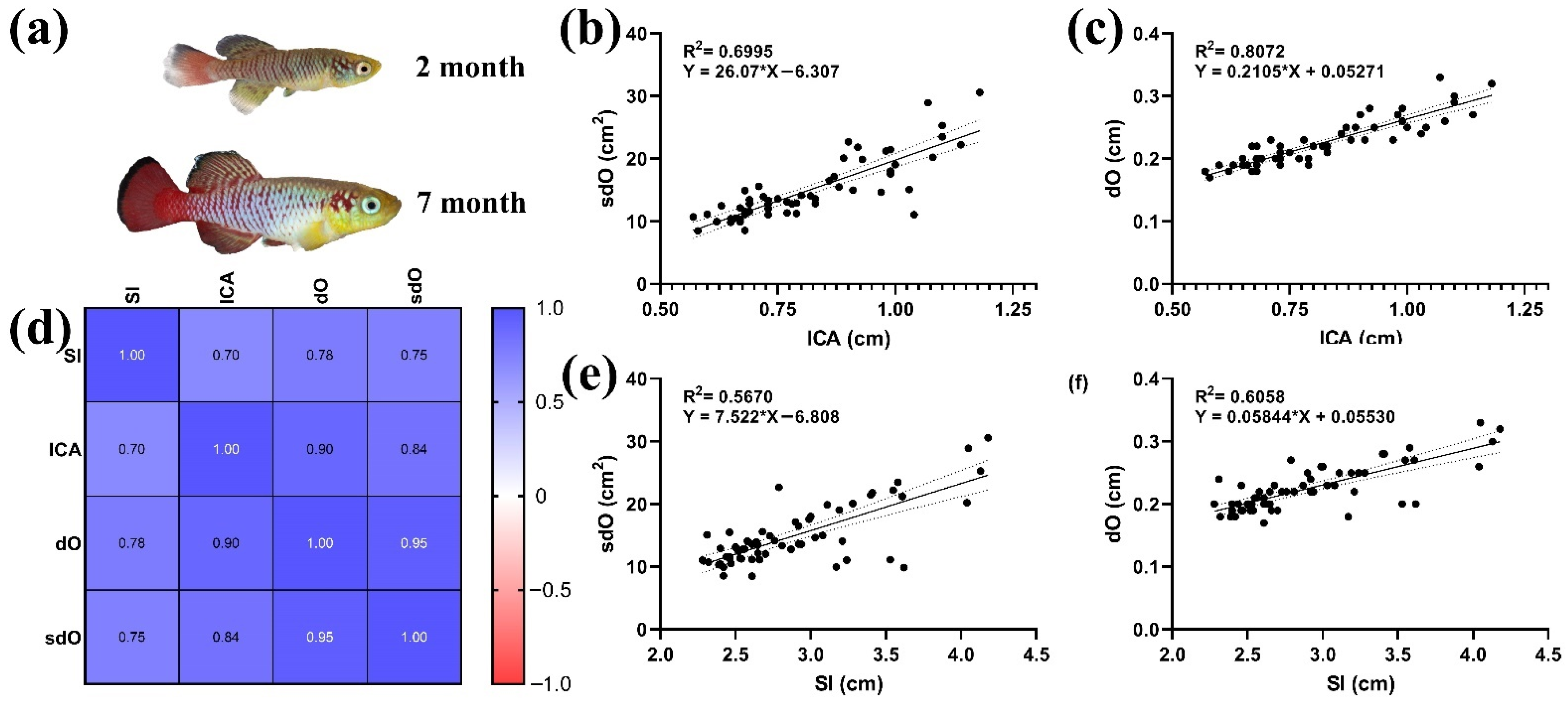
2.5. Fish Photography and Morphometric Measurements
2.6. Eye Elemental Composition
2.7. Statistics
3. Results
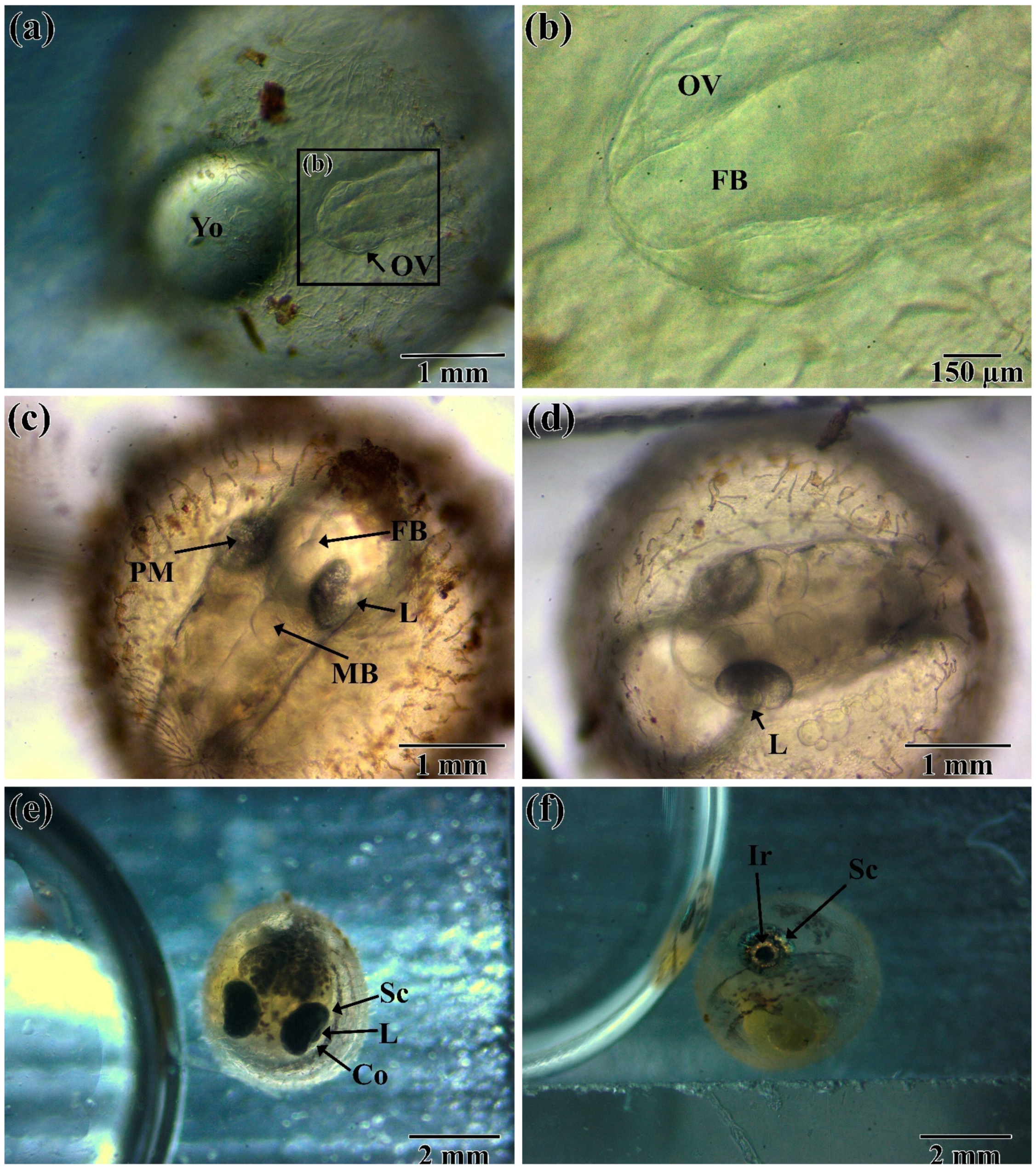
3.1. Embryogenesis
3.2. Post-Hatching Development
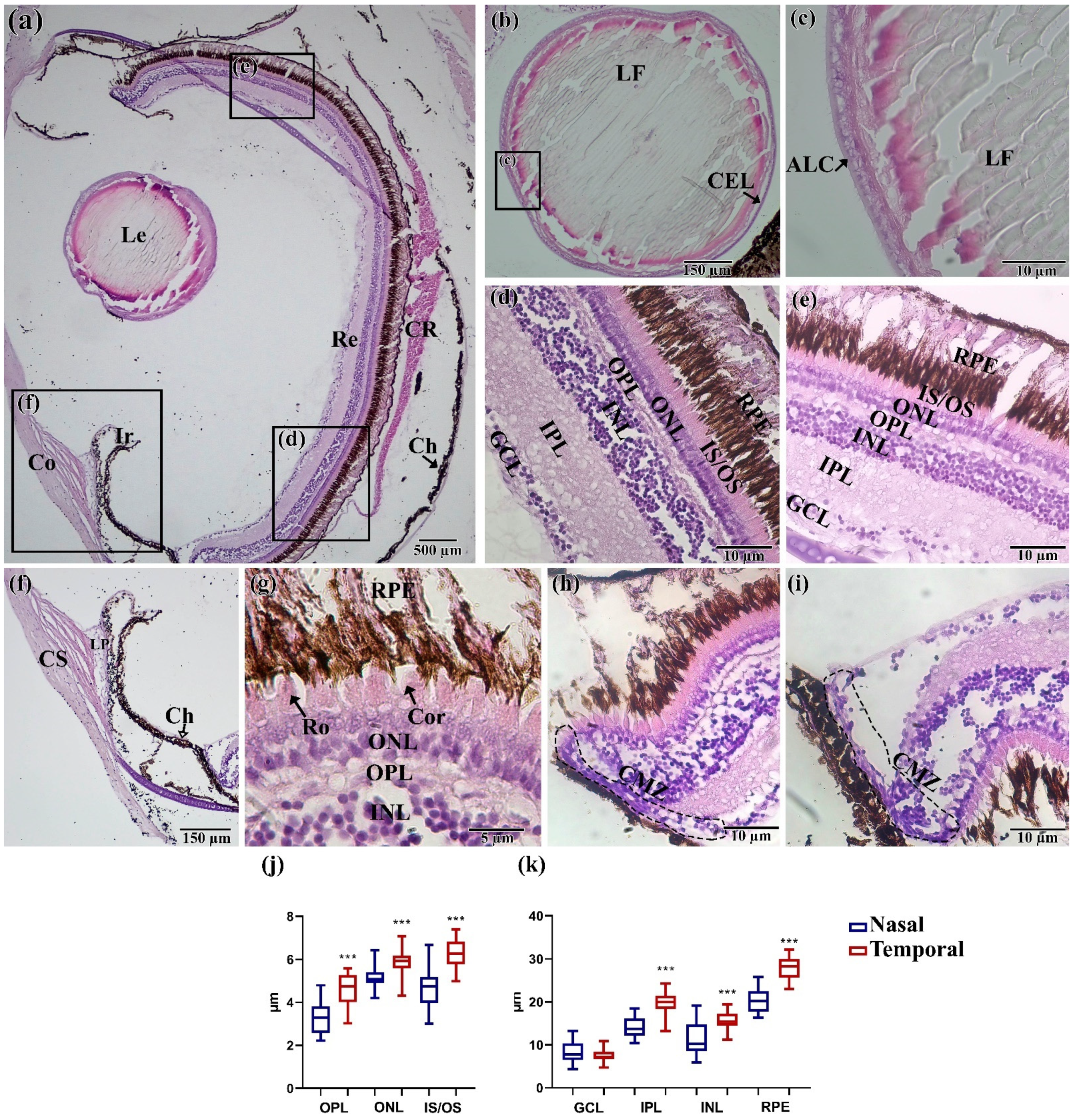
3.3. Histology Assay
3.4. Morphoanatomical Measurements
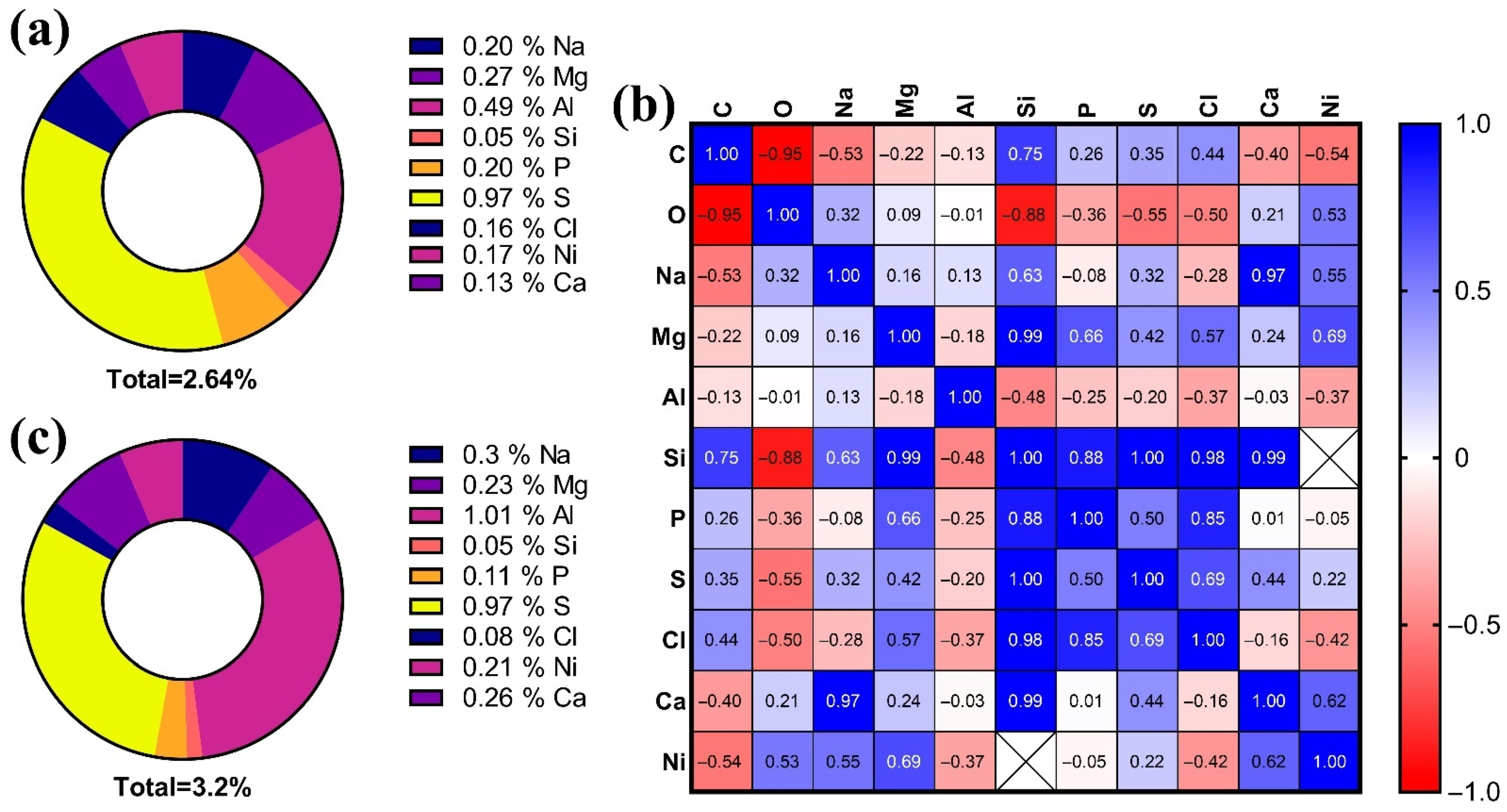
3.5. Elemental Composition of the Lens
- For 2-month old: S > Al > Mg > Na > P > Ni > Cl > Ca > Si;
- For 7-month old: Al > S > Na > Ca > Mg > Ni > P > Cl > Si.
4. Discussion
4.1. Embryology and Post-Hatching
4.2. Histological Assay
4.3. Morphoanatomical Characteristic
4.4. Elemental Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Busserolles, F.; Fitzpatrick, J.L.; Paxton, J.R.; Marshall, N.J.; Collin, S.P. Eye-Size Variability in Deep-Sea Lanternfishes (Myctophidae): An Ecological and Phylogenetic Study. PLoS ONE 2013, 8, e58519. [Google Scholar] [CrossRef]
- Seritrakul, P.; Gross, J.M. Genetic and epigenetic control of retinal development in zebrafish. Curr. Opin. Neurobiol. 2019, 59, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Demir, N. Ichthyology (Ihtiyoloji); Nobel Publishers: Ankara, Turkey, 2009; p. 252. [Google Scholar]
- Furness, A.I. The evolution of an annual life cycle in killifish: Adaptation to ephemeral aquatic environments through embryonic diapause. Biol. Rev. 2016, 91, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Lucas-Sánchez, A.; Almaida-Pagán, P.F.; Mendiola, P.; de Costa, J. Nothobranchius as a model for aging studies. A review. Aging Dis. 2014, 5, 281. [Google Scholar] [PubMed]
- Herrera, M.; Jagadeeswaran, P. Annual fish as a genetic model for aging. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2004, 59, B101–B107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krysanov, E.; Demidova, T. Extensive karyotype variability of African fish genus Nothobranchius (Cyprinodontiformes). Comp. Cytogenet. 2018, 12, 387. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.G. Observations on the biology of Nothobranchius guentheri (Pfeffer) (Cyprinodontidae), an annual fish from the coastal region of east Africa. Afr. J. Trop. Hydrobiol. Fish. 1972, 2, 33–43. [Google Scholar]
- Wildekamp, R.H. A world of killies: Atlas of the oviparous cyprinidontiform fishes of the world. J. Am. Killifish Assoc. 2004, 4, 37–76. [Google Scholar]
- Di Girolamo, N.; Coroneo, M.; Wakefield, D. Epidermal Growth Factor Receptor Signaling Is Partially Responsible for the Increased Matrix Metalloproteinase-1 Expression in Ocular Epithelial Cells after UVB Radiation. Am. J. Pathol. 2005, 167, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Li, G. Effects of resveratrol on longevity, cognitive ability and aging-related histological markers in the annual fish Nothobranchius guentheri. Exp. Gerontol. 2012, 47, 940–949. [Google Scholar] [CrossRef]
- Ray, N.J. Biophysical chemistry of the ageing eye lens. Biophys. Rev. 2015, 7, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Xiaolin, H.; Yuhui, H. Determination of 19 elements in human eye lenses. Biol. Trace. Elem. Res. 1996, 55, 89–98. [Google Scholar]
- Shukla, N.; Moitra, J.; Trivedi, R. Determination of lead, zinc, potassium, calcium, copper and sodium in human cataract lenses. Sci. Total Environ. 1996, 181, 161–165. [Google Scholar] [CrossRef]
- Wistow, G. The human crystallin gene families. Hum. Genom. 2012, 6, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guvatova, Z.; Fedorova, M.; Vershinina, Y.; Pudova, E.; Lipatova, A.; Volodin, V.; Gladysh, N.; Tokarev, A.; Kornev, A.; Pavlov, V.; et al. De Novo Transcriptome Profiling of Brain Tissue from the Annual Killifish Nothobranchius guentheri. Life 2021, 11, 137. [Google Scholar] [CrossRef]
- Tzadik, O.E.; Curtis, J.S.; Granneman, J.E.; Kurth, B.N.; Pusack, T.J.; Wallace, A.A.; Hollander, D.J.; Peebles, E.B.; Stallings, C.D. Chemical archives in fishes beyond otoliths: A review on the use of other body parts as chronological recorders of microchemical constituents for expanding interpretations of environmental, ecological, and life-history changes. Limnol. Oceanogr. Methods 2017, 15, 238–263. [Google Scholar] [CrossRef]
- Wallace, A.A.; Hollander, D.J.; Peebles, E.B. Stable Isotopes in Fish Eye Lenses as Potential Recorders of Trophic and Geographic History. PLoS ONE 2014, 9, e108935. [Google Scholar] [CrossRef] [Green Version]
- Young, M.J.; Larwood, V.; Clause, J.K.; Bell-Tilcock, M.; Whitman, G.; Johnson, R.C.; Feyrer, F. Eye lenses reveal on-togenetic trophic and habitat shifts in an imperiled fish, Clear Lake Hitch Lavinia exilicauda chi. Can. J. Fish. Aquat. Sci. 2021. accepted. [Google Scholar] [CrossRef]
- Kerr, L.A.; Campana, S.E. Chemical Composition of Fish Hard Parts as a Natural Marker of Fish Stocks; Academic Press: Cambridge, MA, USA, 2014; pp. 205–234. [Google Scholar] [CrossRef]
- Podrabsky, J.; Hand, S.C. Physiological strategies during animal diapause: Lessons from brine shrimp and annual killifish. J. Exp. Biol. 2015, 218, 1897–1906. [Google Scholar] [CrossRef] [Green Version]
- Furness, A.I.; Lee, K.; Reznick, D. Adaptation in a variable environment: Phenotypic plasticity and bet-hedging during egg diapause and hatching in an annual killifish. Evolution 2015, 69, 1461–1475. [Google Scholar] [CrossRef]
- Genade, T.; Benedetti, M.; Terzibasi, E.; Roncaglia, P.; Valenzano, D.R.; Cattaneo, A.; Cellerino, A. Annual fishes of the genus Nothobranchius as a model system for aging research. Aging Cell 2005, 4, 223–233. [Google Scholar] [CrossRef]
- El-Bakary, N.E.; Abumandour, M.M. Visual adaptations of the eye of the gilthead sea bream (Sparus aurata). Vet. Res. Commun. 2017, 41, 257–262. [Google Scholar] [CrossRef]
- Howland, H.C.; Howland, M. A standard nomenclature for the axes and planes of vertebrate eyes. Vis. Res. 2008, 48, 1926–1927. [Google Scholar] [CrossRef] [Green Version]
- Suvarna, S.K.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Chervova, L.S.; Lapshin, D.N. Behavioral control of the efficiency of pharmacological anesthesia in fish. J. Ichthyol. 2011, 51, 1126–1132. [Google Scholar] [CrossRef]
- García-Melo, J.E.; García-Melo, L.J.; Garcia-Melo, J.D.; Rojas-Briñez, D.K.; Guevara, G.; Maldonado-Ocampo, J.A. Photafish system: An affordable device for fish photography in the wild. Zootaxa 2019, 4554, 141–172. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Kweon, H.S.; Park, E.H. Improved photographic technique for enhancing the morphological resolution of small juvenile fishes. Ichthyol. Res. 1999, 46, 423–425. [Google Scholar] [CrossRef]
- Lee, Y.; Drake, A.C.; Thomas, N.O.; Ferguson, L.G.; Chappell, P.E.; Shay, K.P. Dietary resveratrol increases mid-life fecundity of female Nothobranchius guentheri. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 208, 71–76. [Google Scholar] [CrossRef]
- Petrtyl, M.; Kalous, L.; MEMİŞ, D. Comparison of manual measurements and computer-assisted image analysis in fish morphometry. Turk. J. Vet. Anim. Sci. 2014, 38, 88–94. [Google Scholar] [CrossRef]
- Ding, L.; Kuhne, W.W.; Hinton, D.E.; Song, J.; Dynan, W.S. Quantifiable Biomarkers of Normal Aging in the Japanese Medaka Fish (Oryzias latipes). PLoS ONE 2010, 5, e13287. [Google Scholar] [CrossRef]
- Nunziata, C.A. Water incubating Nothobranchius eggs. J. Am. Killifish Assoc. 1989, 22, 3–22. [Google Scholar]
- Shedd, T.R.; Widder, M.W.; Toussaint, M.W.; Sunkel, M.C.; Hull, E. Evaluation of the annual killifish Nothobranchius guentheri as a tool for rapid acute toxicity screening. Environ. Toxicol. Chem. Int. J. 1999, 18, 2258–2261. [Google Scholar]
- Hull, E. The Role of Light in Controlling the Onset and Duration of Embryonic Diapause in Nothobranchius guentheri. Ph.D. Thesis, New York University, New York, NY, USA, 1986. [Google Scholar]
- Matias, J.R.; Markofsky, J. The survival of embryos of the annual fishNothobranchius guentheri exposed to temperature extremes and the subsequent effects on embryonic Diapause. J. Exp. Zool. 1978, 204, 219–227. [Google Scholar] [CrossRef]
- Braunbeck, T.; Lammer, E. Fish embryo toxicity assays. Ger. Fed. Environ. Agency 2006, 298, 7–119. [Google Scholar]
- Tozzini, E.T.; Baumgart, M.; Battistoni, G.; Cellerino, A. Adult neurogenesis in the short-lived teleost Nothobranchius furzeri: Localization of neurogenic niches, molecular characterization and effects of aging. Aging Cell 2012, 11, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Vanhunsel, S.; Bergmans, S.; Beckers, A.; Etienne, I.; Van Houcke, J.; Seuntjens, E.; Arckens, L.; De Groef, L.; Moons, L. The killifish visual system as an in vivo model to study brain aging and rejuvenation. Aging Mech. Dis. 2021, 7, 1–17. [Google Scholar] [CrossRef]
- Johns, P.R. Growth of the adult goldfish eye. III. Source of the new retinal cells. J. Comp. Neurol. 1977, 176, 343–357. [Google Scholar] [CrossRef] [Green Version]
- A Raymond, P.; Barthel, L.K.; Bernardos, R.L.; Perkowski, J.J. Molecular characterization of retinal stem cells and their niches in adult zebrafish. BMC Dev. Biol. 2006, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Berois, N.; Arezo, M.J.; Papa, G.N.; Chalar, C. Life Cycle, Reproduction, and Development in Annual Fishes. In Annual Fishes: Life History Strategy, Diversity and Evolution; CRC Press: Boca Raton, FL, USA, 2016; pp. 33–45. [Google Scholar]
- Iwamatsu, T. The Biology of the Medaka; Scientist Co.: Tokyo, Japan, 1993; p. 324. (In Japanese) [Google Scholar]
- Kroll, W. Morphological and behavioral embryology and spontaneous diapause in the African killifish, Aphyosemion gardneri. Environ. Biol. Fishes 1984, 11, 21–28. [Google Scholar] [CrossRef]
- Easter, S.S., Jr.; Nicola, G.N. The development of vision in the zebrafish (Danio rerio). Dev. Biol. 1996, 180, 646–663. [Google Scholar] [CrossRef] [Green Version]
- Howland, H.C.; Merola, S.; Basarab, J.R. The allometry and scaling of the size of vertebrate eyes. Vis. Res. 2004, 44, 2043–2065. [Google Scholar] [CrossRef]
- Acra, A.; Jurdi, M.; Mu’allem, H.; Karahagopian, Y.; Raffoul, Z. Water Disinfection by Solar Radiation: Assessment and Ap-Plication; IDRC: Ottawa, ON, CA, 1990; p. 89. [Google Scholar]
- Podrabsky, J.; Tingaud-Sequeira, A.; Cerdà, J. Metabolic Dormancy and Responses to Environmental Desiccation in Fish Embryos. In Dormancy and Resistance in Harsh Environments; Springer: Berlin/Heidelberg, Germany, 2010; pp. 203–226. [Google Scholar] [CrossRef]
- Motamedi, M.; Shamsaldini, F.; Teimori, A.; Hesni, M.A. Histomicroscopy and normal anatomy of the adult killifish Aphanius hormuzensis (Teleostei; Aphaniidae) from the Persian Gulf coastal environment. Microsc. Res. Tech. 2018, 82, 466–480. [Google Scholar] [CrossRef]
- Menke, A.L.; Spitsbergen, J.M.; Wolterbeek, A.P.M.; Woutersen, R.A. Normal Anatomy and Histology of the Adult Zebrafish. Toxicol. Pathol. 2011, 39, 759–775. [Google Scholar] [CrossRef]
- Tarboush, R.; Chapman, G.; Connaughton, V. Ultrastructure of the distal retina of the adult zebrafish, Danio rerio. Tissue Cell 2012, 44, 264–279. [Google Scholar] [CrossRef]
- Gleeson, M.; Connaughton, V.; Arneson, L.S. Induction of hyperglycaemia in zebrafish (Danio rerio) leads to mor-phological changes in the retina. Acta Diabetol. 2007, 44, 157–163. [Google Scholar] [CrossRef]
- Liu, T.; Liu, S.; Ma, L.; Li, F.; Zheng, Z.; Chai, R.; Hou, Y.; Xie, Y.; Li, G. Oogenesis, vitellogenin-mediated ovarian degeneration and immune response in the annual fish Nothobranchius guentheri. Fish Shellfish Immunol. 2017, 66, 86–92. [Google Scholar] [CrossRef]
- Cooper, E.; Zapata, A.G.; Barrutia, M.G.; Ramirez, J. Aging changes in lymphopoietic and myelopoietic organs of the annual cyprinodont fish, Nothobranchius guentheri. Exp. Gerontol. 1983, 18, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Cui, P.; Zhang, S.; Li, Z. Aging asymmetry: Systematic survey of changes in age-related biomarkers in the annual fish Nothobranchius guentheri. Fish Physiol. Biochem. 2016, 43, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S.; Slack, B.E.; Uchiyama, J.; Zhdanova, I.V. Zebrafish as a Genetic Model in Biological and Behavioral Gerontology: Where Development Meets Aging in Vertebrates—A Mini-Review. Gerontology 2009, 55, 430–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkstra, P.D.; Maguire, S.M.; Harris, R.M.; Rodriguez, A.A.; DeAngelis, R.S.; Flores, S.A.; Hofmann, H.A. The melanocortin system regulates body pigmentation and social behaviour in a colour polymorphic cichlid fish. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegelbecker, A.; Remele, K.; Pfeifhofer, H.W.; Sefc, K.M. Wasteful carotenoid coloration and its effects on territorial behavior in a cichlid fish. Hydrobiologia 2020, 848, 3683–3698. [Google Scholar] [CrossRef]
- Hrbek, T.; Larson, A. The evolution of diapause in the killifish family Rivulidae (Atherinomorpha, Cyprinodontiformes): A molecular phylogenetic and biogeographic perspective. Evolution 1999, 53, 1200–1216. [Google Scholar] [CrossRef]
- Schmitz, L.; Wainwright, P.C. Nocturnality constrains morphological and functional diversity in the eyes of reef fishes. BMC Evol. Biol. 2011, 11, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maan, M.E.; Sefc, K.M. Colour variation in cichlid fish: Developmental mechanisms, selective pressures and evolutionary consequences. Semin. Cell Dev. Biol. 2013, 24, 516–528. [Google Scholar] [CrossRef] [Green Version]
- Quaeck-Davies, K.; Bendall, V.A.; MacKenzie, K.M.; Hetherington, S.; Newton, J.; Trueman, C.N. Teleost and elas-mobranch eye lenses as a target for life-history stable isotope analyses. PeerJ 2018, 6, e4883. [Google Scholar] [CrossRef] [PubMed]
- Bird, C.S.; Veríssimo, A.; Magozzi, S.; Abrantes, K.; Aguilar, A.; Al-Reasi, H.; Barnett, A.; Bethea, D.M.; Biais, G.; Borrell, A.; et al. A global perspective on the trophic geography of sharks. Nat. Ecol. Evol. 2018, 2, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Greiling, T.M.; Houck, S.A.; Clark, J.I. The zebrafish lens proteome during development and aging. Mol. Vis. 2009, 15, 2313–2325. [Google Scholar] [PubMed]
- Bron, A.J.; Vrensen GF, J.M.; Koretz, J.; Maraini, G.; Harding, J.J. The ageing lens. Ophthalmologica 2000, 214, 86–104. [Google Scholar] [CrossRef]
- Langford-Smith, A.; Tilakaratna, V.; Lythgoe, P.R.; Clark, S.J.; Bishop, P.N.; Day, A.J. Age and Smoking Related Changes in Metal Ion Levels in Human Lens: Implications for Cataract Formation. PLoS ONE 2016, 11, e0147576. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, S.; Vasavada, A.R.; Praveen, M.R.; Ananthan, R.; Reddy, G.B.; Tripathi, H.; Ganatra, D.A.; Arora, A.I.; Patel, A.R. Exploration of Molecular Factors Impairing Superoxide Dismutase Isoforms Activity in Human Senile Cataractous Lenses. Investig. Opthalmol. Vis. Sci. 2013, 54, 6224–6233. [Google Scholar] [CrossRef] [Green Version]
- Yanshole, L.V.; Cherepanov, I.V.; Snytnikova, O.A.; Yanshole, V.V.; Sagdeev, R.Z.; Tsentalovich, Y.P. Cata-ract-specific posttranslational modifications and changes in the composition of urea-soluble protein fraction from the rat lens. Mol. Vis. 2013, 19, 2196. [Google Scholar]
- Ramana, B.V.; Raju, T.N.; Kumar, V.V.; Reddy, P.U.M. Defensive Role of Quercetin Against Imbalances of Calcium, Sodium, and Potassium in Galactosemic Cataract. Biol. Trace Element Res. 2007, 119, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Duncan, G.; Wormstone, I.M.; Wormstone, M. Calcium cell signalling and cataract: Role of the endoplasmic reticulum. Eye 1999, 13, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, M.; Xu, F.; Suhonen, H.; Gourain, V.; Yang, L.; Ho, N.Y.; Helfen, L.; Schröck, A.; Etard, C.; Grabher, C.; et al. Melanosomes in pigmented epithelia maintain eye lens transparency during zebrafish embryonic development. Sci. Rep. 2016, 6, 25046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dove, S.G.; Kingsford, M.J. Use of otoliths and eye lenses for measuring trace-metal incorporation in fishes: A biogeo-graphic study. Mar. Biol. 1998, 130, 377–387. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | O | Na | Mg | Al | Si | P | S | Cl | Ca | Ni | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 mo. | Mean | 64.77 | 32.58 | 0.2 | 0.27 | 0.49 | 0.05 | 0.2 | 0.97 | 0.16 | 0.13 | 0.17 |
| SD | 2.81 | 3.56 | 0.02 | 0.08 | 0.36 | 0.005 | 0.06 | 0.71 | 0.08 | 0.05 | 0.02 | |
| Min | 62.23 | 28.61 | 0.18 | 0.19 | 0.16 | 0.05 | 0.12 | 0.56 | 0.1 | 0.09 | 0.15 | |
| Max | 67.57 | 35.88 | 0.23 | 0.39 | 0.81 | 0.06 | 0.27 | 2.04 | 0.28 | 0.22 | 0.19 | |
| CoF (%) | 4.34 | 10.95 | 12.99 | 32.60 | 73.55 | 10.83 | 33.91 | 73.76 | 49.84 | 42.98 | 11.76 | |
| 7 mo. | Mean | 64.09 | 32.74 | 0.30 | 0.22 | 1.01 | 0.05 | 0.105 | 0.96 | 0.07 | 0.26 | 0.2 |
| SD | 3.55 | 3.079 | 0.15 | 0.16 | 0.7 | 0 | 0.03 | 0.229 | 0.01 | 0.22 | 0.09 | |
| Min | 60.84 | 28.56 | 0.15 | 0.05 | 0.33 | 0.05 | 0.07 | 0.69 | 0.06 | 0.09 | 0.08 | |
| Max | 68.11 | 35.16 | 0.52 | 0.37 | 1.75 | 0.05 | 0.13 | 1.23 | 0.1 | 0.58 | 0.29 | |
| CoF (%) | 5.54 | 9.4 | 52.57 | 71.73 | 69.37 | 0.000 | 28.57 | 23.67 | 22.04 | 85.02 | 43.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikiforov-Nikishin, D.L.; Irkha, V.A.; Kochetkov, N.I.; Kalita, T.L.; Nikiforov-Nikishin, A.L.; Blokhin, E.E.; Antipov, S.S.; Makarenkov, D.A.; Zhavnerov, A.N.; Glebova, I.A.; et al. Some Aspects of Development and Histological Structure of the Visual System of Nothobranchius Guentheri. Animals 2021, 11, 2755. https://doi.org/10.3390/ani11092755
Nikiforov-Nikishin DL, Irkha VA, Kochetkov NI, Kalita TL, Nikiforov-Nikishin AL, Blokhin EE, Antipov SS, Makarenkov DA, Zhavnerov AN, Glebova IA, et al. Some Aspects of Development and Histological Structure of the Visual System of Nothobranchius Guentheri. Animals. 2021; 11(9):2755. https://doi.org/10.3390/ani11092755
Chicago/Turabian StyleNikiforov-Nikishin, Dmitry L., Vladimir A. Irkha, Nikita I. Kochetkov, Tatyana L. Kalita, Alexei L. Nikiforov-Nikishin, Eduard E. Blokhin, Sergei S. Antipov, Dmitry A. Makarenkov, Alexey N. Zhavnerov, Irina A. Glebova, and et al. 2021. "Some Aspects of Development and Histological Structure of the Visual System of Nothobranchius Guentheri" Animals 11, no. 9: 2755. https://doi.org/10.3390/ani11092755
APA StyleNikiforov-Nikishin, D. L., Irkha, V. A., Kochetkov, N. I., Kalita, T. L., Nikiforov-Nikishin, A. L., Blokhin, E. E., Antipov, S. S., Makarenkov, D. A., Zhavnerov, A. N., Glebova, I. A., Smorodinskaya, S. V., & Chebotarev, S. N. (2021). Some Aspects of Development and Histological Structure of the Visual System of Nothobranchius Guentheri. Animals, 11(9), 2755. https://doi.org/10.3390/ani11092755