
Genetic Variability Trend of Lusitano Horse Breed Reared in Italy

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and Laboratory Methods
2.2. Genetic Diversity Measures
2.3. Inbreeding Analysis
2.4. Population Structure
3. Results
3.1. Genetic Diversity
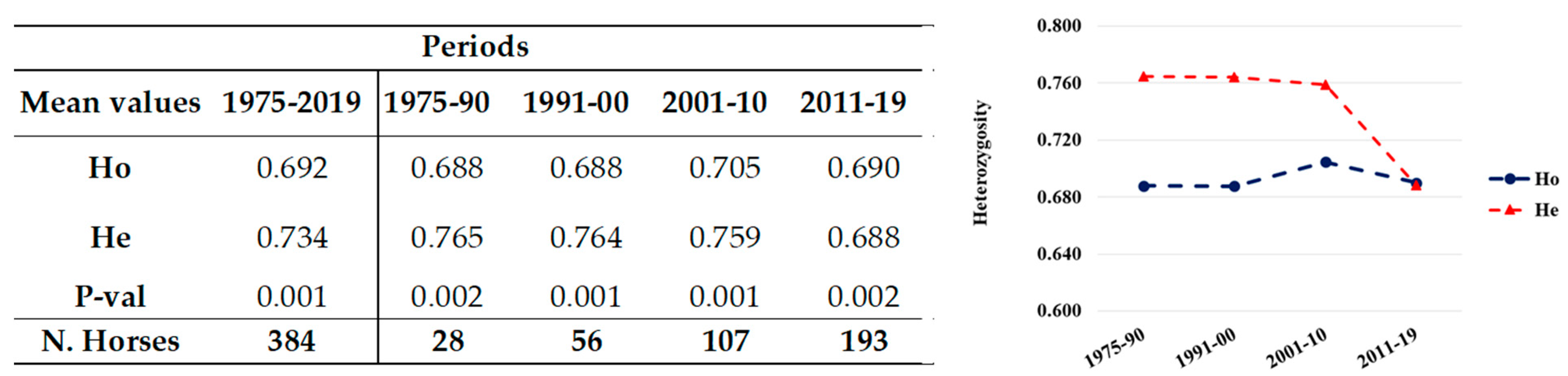
3.1.1. Period 1975–2019
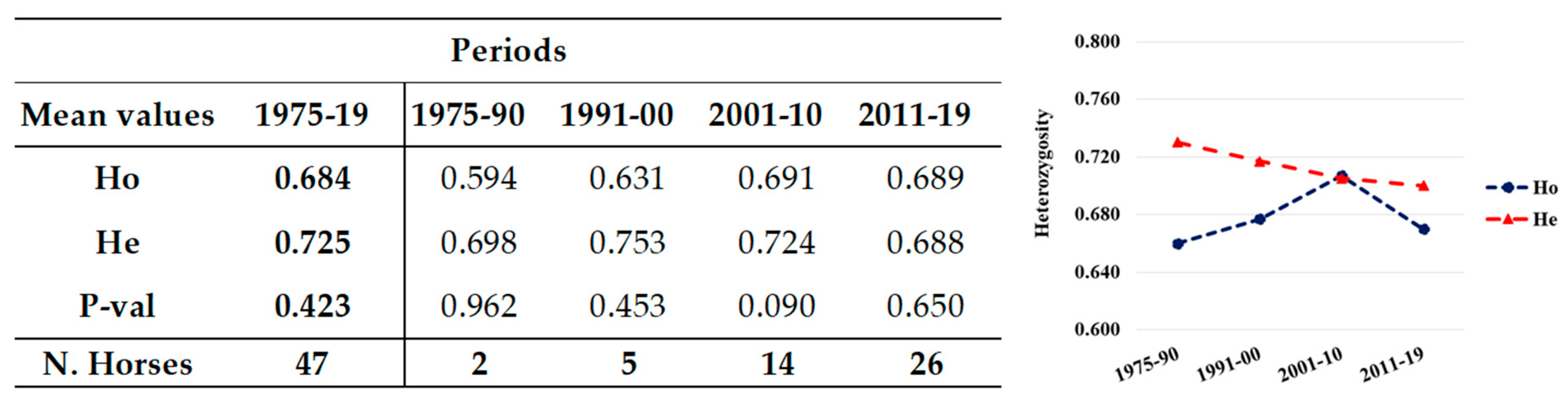
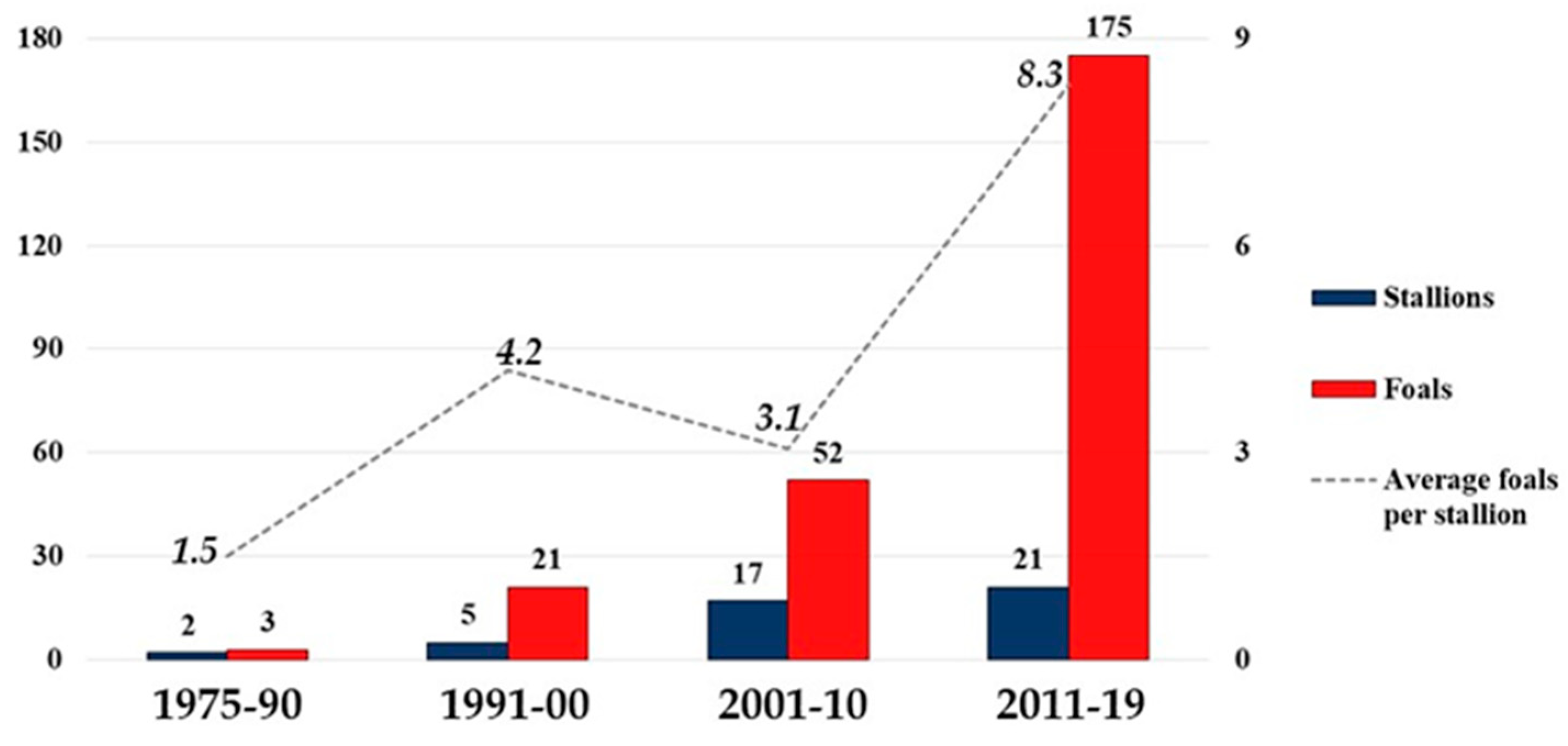
3.1.2. Periods 1975–1990, 1991–2000, 2001–2010, 2011–2019
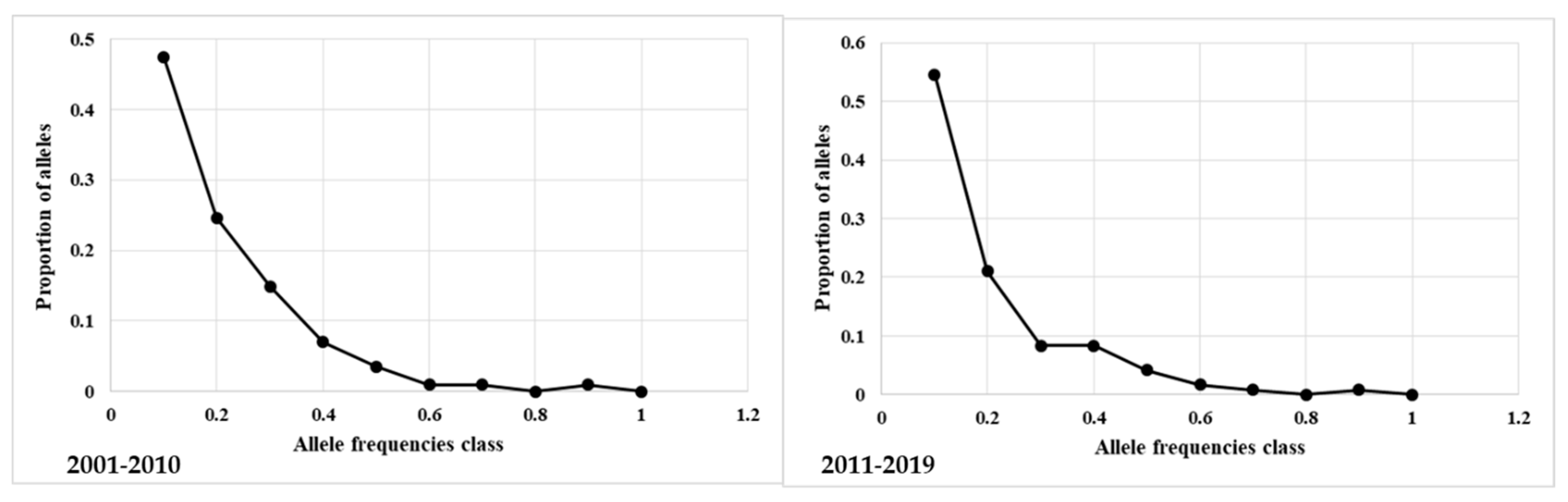
3.2. Inbreeding Analysis
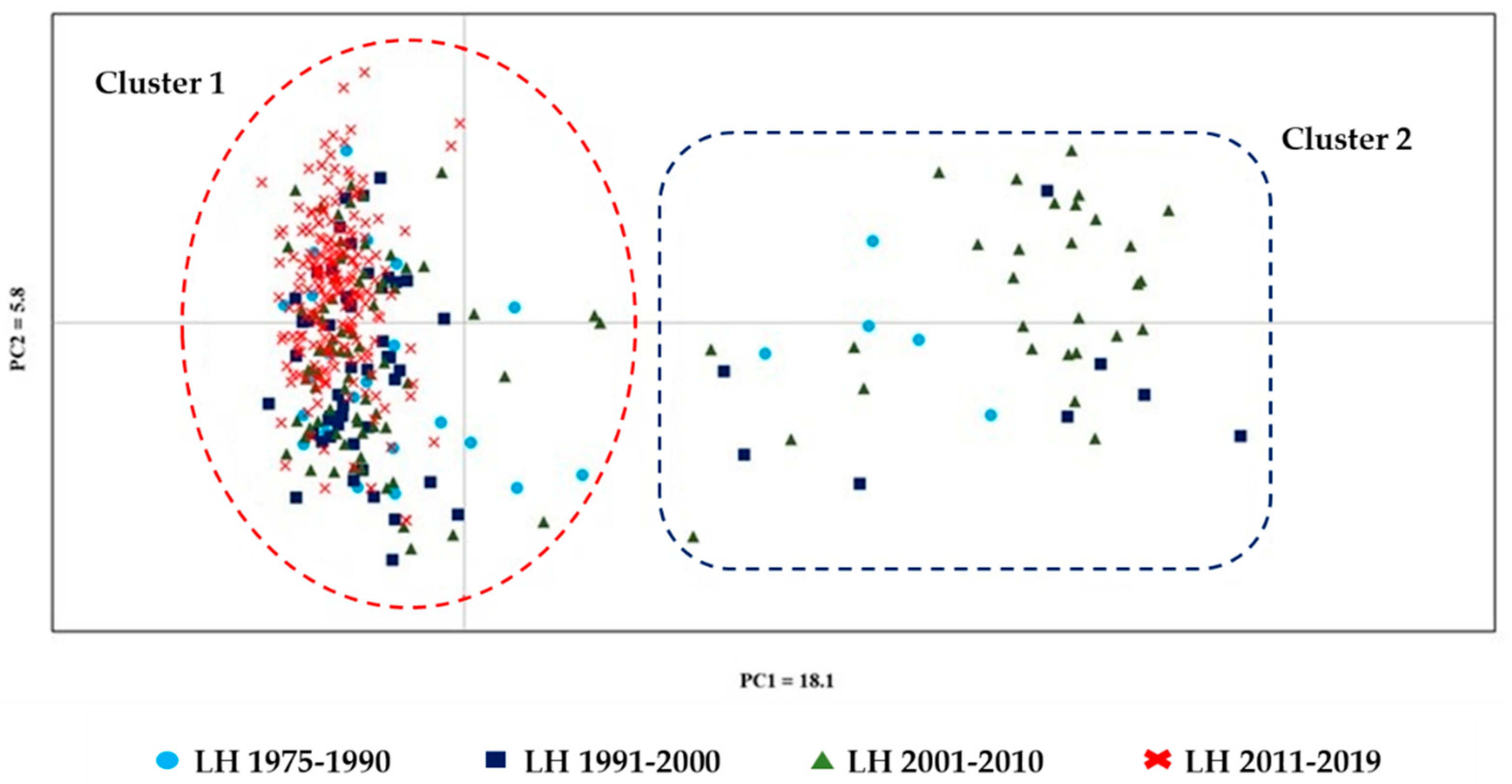
3.3. Population Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vicente, A.A.; Carolino, N.; Gama, L.T. Genetic diversity in the Lusitano horse breed assessed by pedigree analysis. Livest. Sci. 2012, 148, 16–25. [Google Scholar] [CrossRef] [Green Version]
- da Silva Faria, R.A.; Andrade Vicente, A.P.; Duarte Guedes dos Santos, R.I.; Marchi Maiorano, A.; Curi, R.A.; Loyola Chardulo, L.A.; Vasconcelos Silva, J.A. Genetic diversity of Lusitano Horse in Brazil using pedigree information. J. Equine Vet. Sci. 2018, 69, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Vicente, A.P. Characterization and Selection of the Lusitano Horse Breed. Ph.D. Thesis, Faculty of Veterinary Medicine, Lisboa, Portugal, 2015. [Google Scholar]
- Cassetti, G.; Associazione Italiana Cavallo Puro Sangue Lusitano (AICL), Rome, Italy. Personal communication, 2021.
- Luís, C.; Juras, R.; Oom, M.M.; Cothran, E.G. Genetic diversity and relationships of Portuguese and other horse breeds based on protein and microsatellite loci variation. Anim. Genet. 2007, 38, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Vicente, A.A.; Carolino, N.; Gama, L.T. Demographic parameters in Lusitano horse. Arch. Zootec. 2009, 58 (Suppl. S1), 501–504. [Google Scholar]
- Juras, R.; Cothran, E.G. Microsatellites in Lithuanian native horse breeds: Usefulness for parentage testing. Biologija 2004, 4, 6–9. [Google Scholar]
- Zuccaro, A.; Bordonaro, S.; Criscione, A.; Guastella, A.M.; Perrotta, G.; Blasi, M.; D’Urso, G.; Marletta, D. Genetic diversity and admixture analysis of Sanfratellano and three other Italian horse breeds assessed by microsatellite markers. Animal 2008, 2, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Leroy, G.; Callède, L.; Verrier, E.; Mériaux, J.-C.; Ricard, A.; Danchin-Burge, C.; Rognon, X. Genetic diversity of a large set of horse breeds raised in France assessed by microsatellite polymorphism. Genet. Sel. Evol. 2009, 41, 5. [Google Scholar] [CrossRef] [Green Version]
- Bömcke, E.; Gengler, N.; Cothran, E. Genetic variability in the Skyros pony and its relationship with other Greek and foreign horse breeds. Genet. Mol. Biol. 2011, 34, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortés, O.; Dunner, S.; Gama, L.T.; Martínez, A.M.; Delgado, J.V.; Ginja, C.; Jiménez, L.M.; Jordana, J.; Luis, C.; Oom, M.M.; et al. The legacy of Columbus in American horse populations assessed by microsatellite markers. J. Anim. Breed. Genet. 2017, 134, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, M.C.; Strillacci, M.G.; Valiati, P.; Rogliano, E.; Bagnato, A.; Longeri, M. Genetic variability of Akhal-Teke horses bred in Italy. PeerJ 2018, 6, e4889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosenza, M.; La Rosa, V.; Rosati, R.; Chiofalo, V. Genetic diversity of the Italian thoroughbred horse population. Ital. J. Anim. Sci. 2019, 18, 538–545. [Google Scholar] [CrossRef]
- Van de Goor, L.H.; Panneman, H.; van Haeringen, W.A. A proposal for standardization in forensic equine DNA typing: Allele nomenclature for 17 equine-specific STR loci. Anim. Genet. 2010, 41, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Van de Goor, L.H.P.; van Haeringen, W.A.; Lenstra, J.A. Population studies of 17 equine STR for forensic and phylogenetic analysis. Anim. Genet. 2011, 42, 627–633. [Google Scholar] [CrossRef] [PubMed]
- ISAG International Society for Animal Genetics. Available online: http://www.isag.us/Docs/EquineGenParentage2016.pdf (accessed on 30 November 2021).
- Applied Biosystems by Life Technologies User Guide. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/manuals/4346135_Stockmarks_Kits_UG.pdf (accessed on 30 November 2021).
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. Genepop ‘007: A complete re-implementation of the Genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Yang, R.; Boyle, T.J.B. POPGENE Version 1.31 Microsoft Window-Based Freeware for Population Genetic Analysis. 1997. Available online: https://sites.ualberta.ca/~fyeh/popgene.html (accessed on 30 November 2021).
- Goudet, J. FSTAT, a Program to Estimate and Test Gene Diversities and Fixation Indices. 2002. Available online: https://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 30 November 2021).
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Cornuet, J.M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef] [PubMed]
- Luikart, G.L.; Allendrof, F.W.; Cornuet, J.M.; Sherwin, W.B. Distortion of allele frequency distributions provides a test for recent population bottlenecks. J. Hered. 1998, 89, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A Computer Program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Ustyantseva, A.V.; Khrabrova, L.A.; Abramova, N.V.; Ryabova, T.N. Genetic characterization of Akhal-Teke horse subpopulations using 17 microsatellite loci. IOP Conf. Ser. Earth Environ. Sci. 2019, 341, 012070. [Google Scholar] [CrossRef]
- Szwaczkowski, T.; Greguła-Kania, M.; Stachurska, A.; Borowska, A.; Jaworski, Z.; Gruszecki, T.M. Inter-and intra-genetic diversity in the Polish Konik horse: Implications for the conservation program. Can. J. Anim. Sci. 2016, 96, 570–580. [Google Scholar] [CrossRef]
- Conant, E.K.; Juras, R.; Cothran, E.G. A microsatellite analysis of five Colonial Spanish horse populations of the southeastern United States. Anim. Genet. 2012, 43, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Kalashnikov, V.; Khrabrova, L.; Blohina, N.; Zaitcev AKalashnikova, T. Dynamics of the inbreeding coefficient and homozygosity in Thoroughbred Horses in Russia. Animals 2020, 10, 1217. [Google Scholar] [CrossRef] [PubMed]
- Głażewska, I.; Gralak, B.; Naczk, A.M.; Prusak, B. Genetic diversity and population structure of Polish Arabian horses assessed through breeding and microsatellite data. Anim. Sci. J. 2018, 89, 735–742. [Google Scholar] [CrossRef]
- Di Rienzo, A.; Peterson, A.C.; Garza, J.C.; Valdes, A.M.; Slatkin, M.; Freimer, N.B. Mutational processes of simple-sequence repeat loci in human populations. Proc. Natl. Acad. Sci. USA 1994, 91, 3166–3170. [Google Scholar] [CrossRef] [Green Version]
- Spencer, C.C.; Neigel, J.E.; Leberg, P.L. Experimental evaluation of the usefulness of microsatellite DNA for detecting demographic bottlenecks. Mol. Ecol. 2000, 9, 1517–1528. [Google Scholar] [CrossRef]
- Sharma, R.; Kumar, B.; Arora, R.; Ahlawat, S.; Mishra, A.K.; Tantia, M.S. Genetic diversity estimates point to immediate efforts for conserving the endangered Tibetan sheep of India. Meta Gene 2016, 8, 14–20. [Google Scholar] [CrossRef]
- Funk, S.M.; Guedaoura, S.; Juras, R.; Raziq, A.; Landolsi, F.; Luís, C.; Martínez Martínez, A.; Musa Mayaki, A.; Mujica, F.; Oom, M.M.; et al. Major inconsistencies of inferred population genetic structure estimated in a large set of domestic horse breeds using microsatellites. Ecol. Evol. 2020, 10, 4261–4279. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Horses | Total Period | Generation Interval Periods | |||
|---|---|---|---|---|---|
| 1975–2019 | 1975–1990 | 1991–2000 | 2001–2010 | 2010–2019 | |
| Total population: Related and unrelated | 384 | 28 | 56 | 107 | 193 |
| Unrelated | 47 | 2 | 5 | 14 | 26 |
| Mean Value | Periods | |||||
|---|---|---|---|---|---|---|
| 1975–2019 | 1975–1990 | 1991–2000 | 2001–2010 | 2011–2019 | ||
| Total | Fis | 0.057 | 0.102 | 0.101 | 0.072 | −0.002 |
| Unrelated | Fis | 0.056 | 0.208 | 0.206 | 0.045 | 0.019 |
| 1975–1990 | 1991–2000 | 2001–2010 | 2011–2019 | |
|---|---|---|---|---|
| 1975–1990 | 6.781 | 6.770 | 6.733 | 6.811 * |
| 1991–2000 | −0.036 | 6.831 | 6.756 | 6.850 * |
| 2001–2010 | −0.004 | −0.005 | 6.694 | 6.784 * |
| 2011–2019 | 0.166 * | 0.180 * | 0.182 * | 6.509 |
| Dams Mother of | Sires Father of | |||||||
|---|---|---|---|---|---|---|---|---|
| 1975–1990 | 1991–2000 | 2001–2010 | 2011–2019 | 1975–1990 | 1991–2000 | 2001–2010 | 2011–2019 | |
| 1 foal | 1 | 7 | 20 | 35 | 2 | 6 | 10 | 24 |
| 2 foals | - | 6 | 7 | 21 | 1 | 2 | 4 | 10 |
| 3 foals | - | 1 | 4 | 8 | - | 1 | 1 | - |
| 4 foals | - | - | 1 | 8 | - | - | 1 | 3 |
| 5 foals | - | - | - | 6 | - | - | 1 | 1 |
| 6 foals | - | - | - | 3 | - | 1 | 2 | 1 |
| 7 foals | - | - | - | 1 | - | - | 1 | 1 |
| ≥8 foals | - | - | - | - | - | - | - | 4 |
| Total | 1 | 14 | 32 | 82 | 3 | 10 | 27 | 41 |
| Sire ID | N. Dams | N. Foal | N. Full sibs |
|---|---|---|---|
| A | 39 | 50 | 22 |
| B | 28 | 46 | 33 |
| C | 14 | 22 | 14 |
| D | 11 | 11 | 0 |
| E | 6 | 7 | 2 |
| F | 5 | 6 | 2 |
| G | 4 | 4 | 0 |
| H | 3 | 4 | 2 |
| I | 3 | 4 | 2 |
| J | 2 | 2 | 0 |
| K | 2 | 2 | 0 |
| L | 2 | 2 | 0 |
| M | 2 | 5 | 5 |
| N | 1 | 2 | 1 |
| Total (14) | 122 | 167 | 83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cozzi, M.C.; Valiati, P.; Longeri, M.; Ferreira, C.; Abreu Ferreira, S. Genetic Variability Trend of Lusitano Horse Breed Reared in Italy. Animals 2022, 12, 98. https://doi.org/10.3390/ani12010098
Cozzi MC, Valiati P, Longeri M, Ferreira C, Abreu Ferreira S. Genetic Variability Trend of Lusitano Horse Breed Reared in Italy. Animals. 2022; 12(1):98. https://doi.org/10.3390/ani12010098
Chicago/Turabian StyleCozzi, Maria Cristina, Paolo Valiati, Maria Longeri, Carlos Ferreira, and Sofia Abreu Ferreira. 2022. "Genetic Variability Trend of Lusitano Horse Breed Reared in Italy" Animals 12, no. 1: 98. https://doi.org/10.3390/ani12010098
APA StyleCozzi, M. C., Valiati, P., Longeri, M., Ferreira, C., & Abreu Ferreira, S. (2022). Genetic Variability Trend of Lusitano Horse Breed Reared in Italy. Animals, 12(1), 98. https://doi.org/10.3390/ani12010098