
Anthelmintic Efficacy of Palmarosa Oil and Curcuma Oil against the Fish Ectoparasite Gyrodactylus kobayashii (monogenean)

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fish and Parasite
2.3. In Vivo Screening of 16 EOs
2.4. Anthelmintic Efficacy of Selected EOs
2.4.1. In Vitro Assay
2.4.2. In Vivo Assay
2.5. Acute Toxicity Tests
2.6. The Physiological and Histological Effects of Eos on Goldfish
2.7. Statistical Analysis
3. Results
3.1. In Vivo Anthelmintic Efficacy of 16 EOs
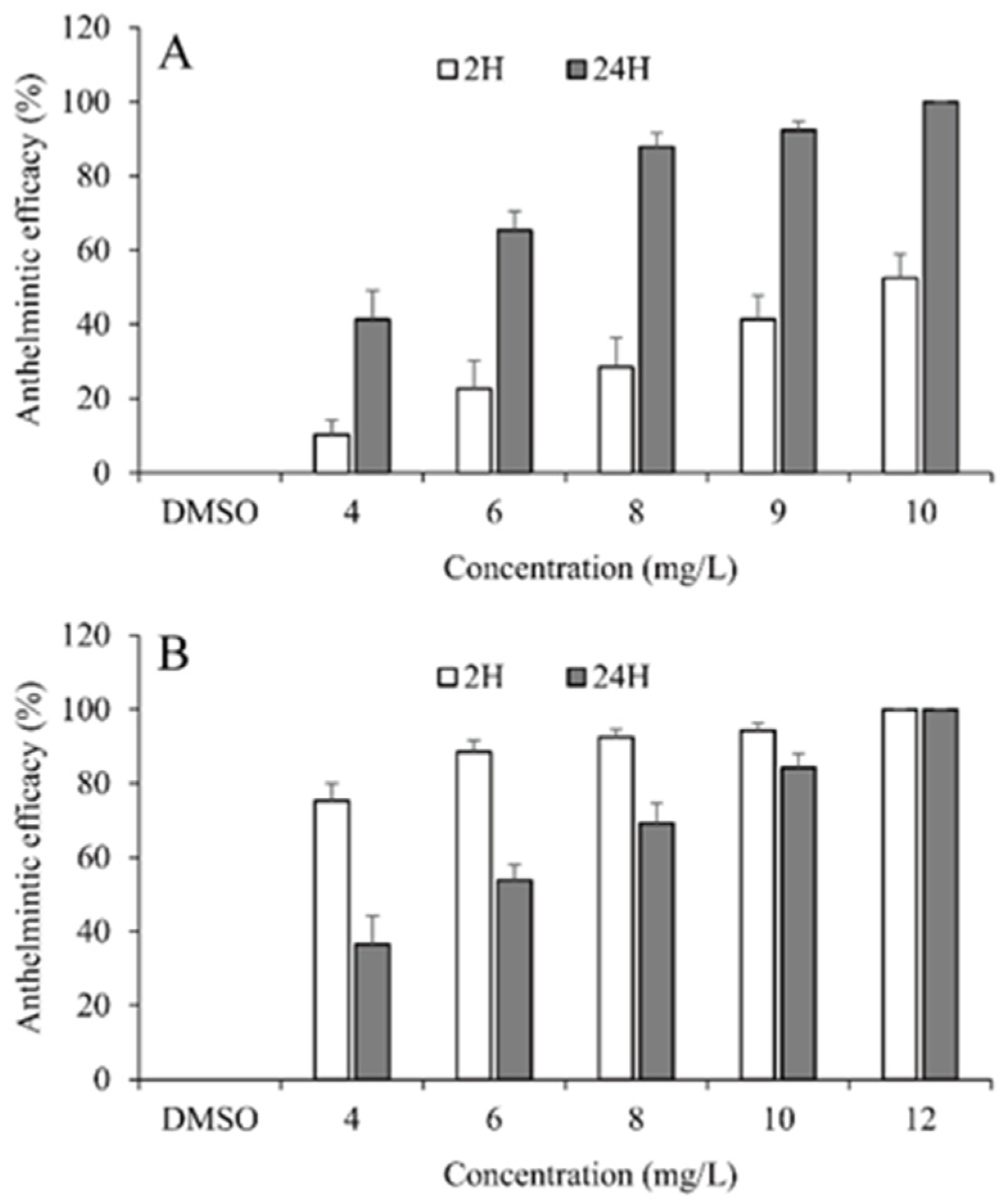
3.2. Anthelmintic Efficacy of Selected EOs
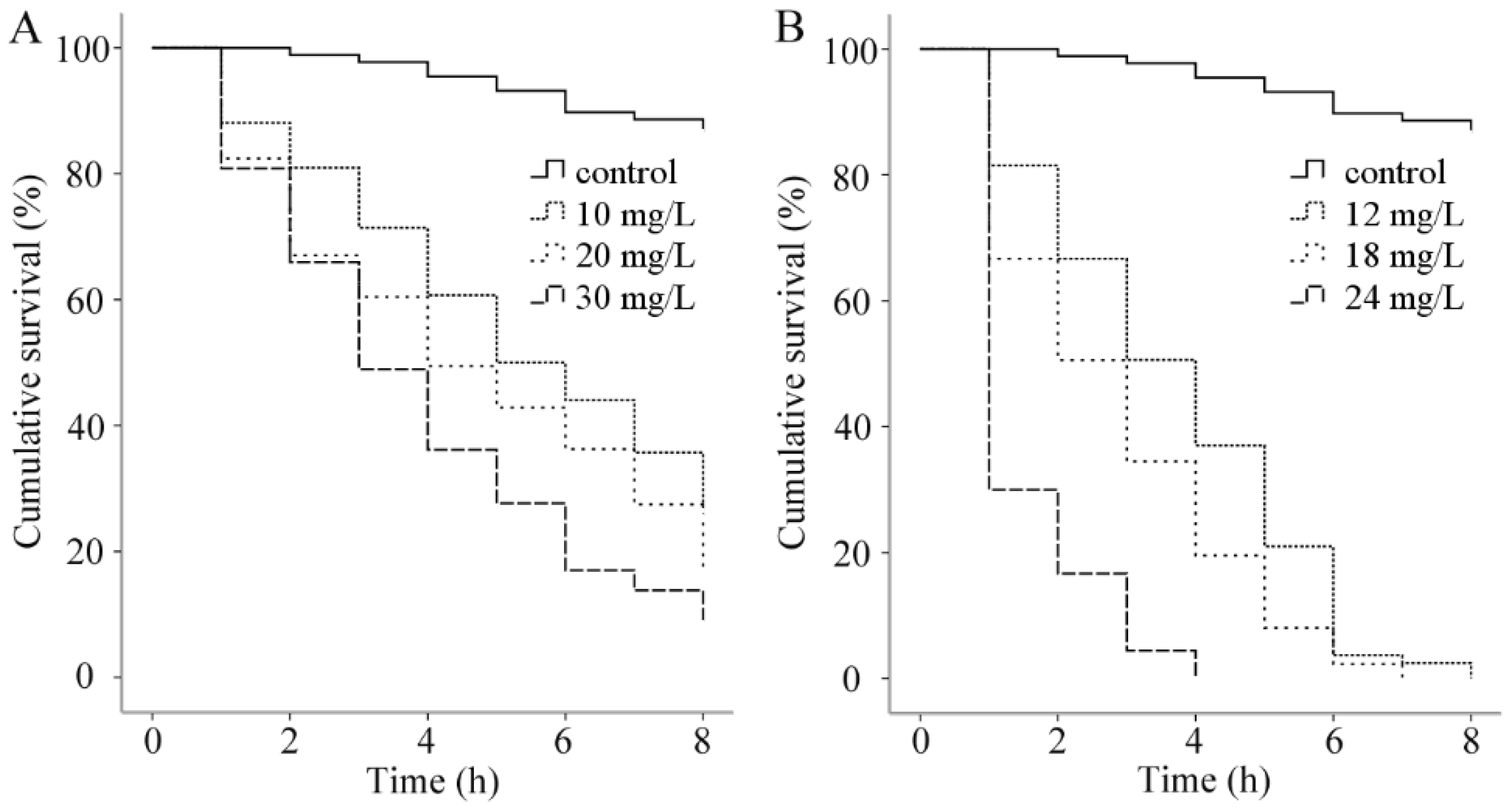
3.3. Acute Toxicity Tests against Goldfish
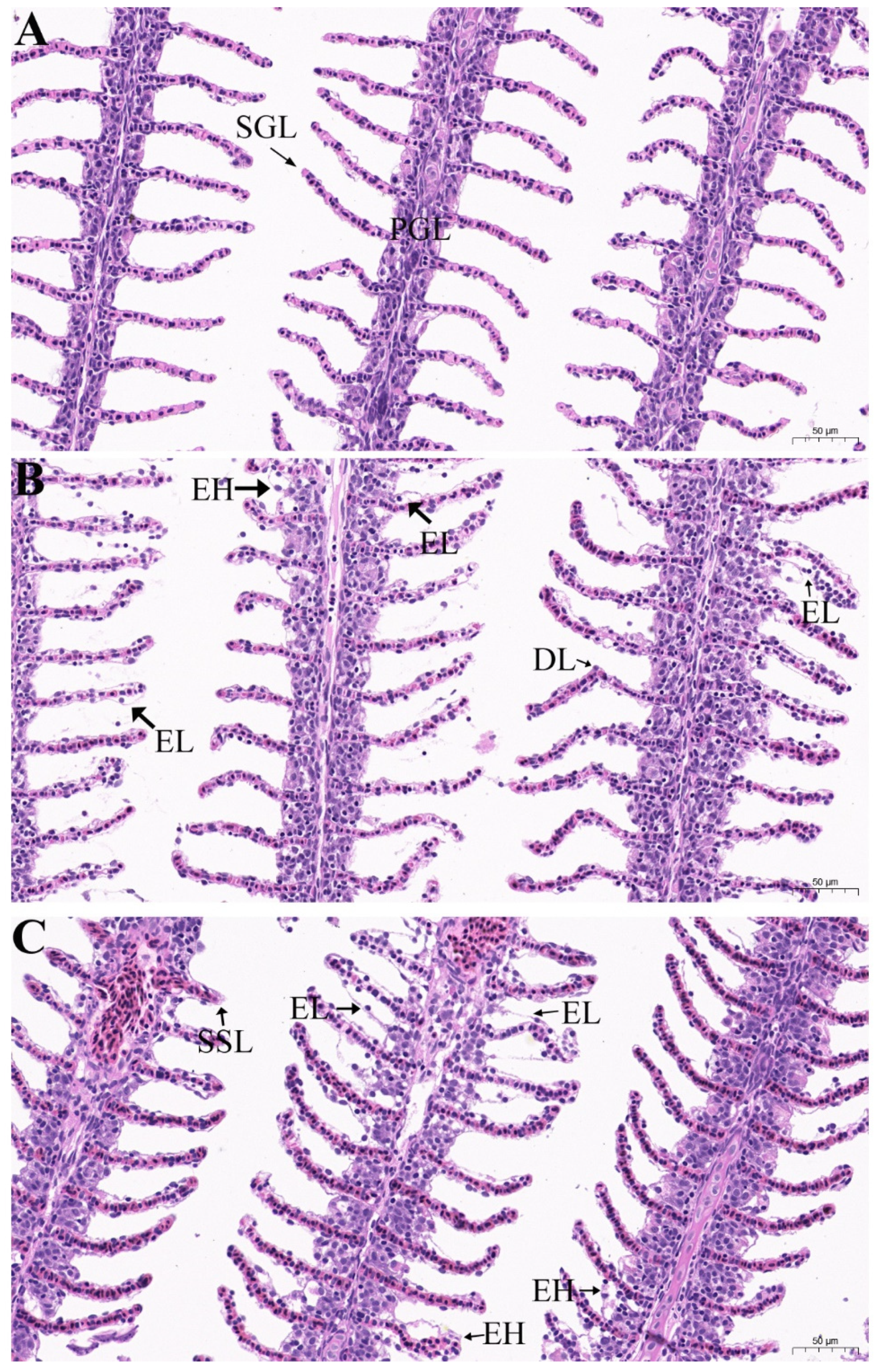
3.4. The Physiological and Histological Effects of EOs on Goldfish
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kohn, A.; Cohen, S.C.; Salgado-Maldonado, G. Checklist of Monogenea parasites of freshwater and marine fishes, amphibians and reptiles from Mexico, Central America and Caribbean. Zootaxa 2006, 1289, 1–114. [Google Scholar]
- Kuchta, R.; Řehulková, E.; Francová, K.; Scholz, T.; Morand, S.; Šimková, A. Diversity of monogeneans and tapeworms in cypriniform fishes across two continents. Int. J. Parasitol. 2020, 50, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Gonzalez, A.; Becker, J.A.; Hutson, K.S. Parasite dispersal from the ornamental goldfish trade. In Advances in Parasitology; Rollinson, D., Stothard, J.R., Eds.; Elsevier, Academic Press Inc.: San Diego, CA, USA, 2018; pp. 239–281. [Google Scholar]
- Bakke, T.A.; Cable, J.; Harris, P.D. The biology of gyrodactylid monogeneans: The “Russian-doll killers”. Adv. Parasitol. 2007, 64, 161–376. [Google Scholar] [PubMed]
- Cone, D.K.; Odense, P.H. Pathology of five species of Gyrodactylus Nordmann, 1832 (Monogenea). Can. J. Zool. 1984, 62, 1084–1088. [Google Scholar] [CrossRef]
- Tu, X.; Ling, F.; Huang, A.; Wang, G. An infection of Gyrodactylus kobayashii Hukuda, 1940 (Monogenea) associated with the mortality of goldfish (Carassius auratus) from central China. Parasitol. Res. 2015, 114, 737–745. [Google Scholar] [CrossRef]
- Pettersen, R.A.; Hytterod, S.; Vollestad, L.A.; Mo, T.A. Osmoregulatory disturbances in Atlantic salmon, Salmo salar L., caused by the monogenean Gyrodactylus salaris. J. Fish Dis. 2013, 36, 67–70. [Google Scholar] [CrossRef]
- Schelkle, B.; Shinn, A.P.; Peeler, E.; Cable, J. Treatment of gyrodactylid infections in fish. Dis. Aquat. Org. 2009, 86, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Pietrak, M.; Backman, S. Treatment of lumpfish (Cyclopterus lumpus L.) infected with Gyrodactylus cyclopteri (Scyborskaya 1948). J. Fish Dis. 2018, 41, 721–723. [Google Scholar] [CrossRef]
- Zhou, S.; Li, W.X.; Wang, Y.Q.; Zou, H.; Wu, S.G.; Wang, G.T. Anthelmintic efficacies of three common disinfectants and extracts of four traditional Chinese medicinal plants against Gyrodactylus kobayashii (Monogenea) in goldfish (Carassius auratus). Aquaculture 2017, 466, 72–77. [Google Scholar] [CrossRef]
- Rowland, S.J.; Nixon, M.; Landos, M.; Mifsud, C.; Read, P.; Boyd, P. Effects of formalin on water quality and parasitic monogeneans on silver perch (Bidyanus bidyanus Mitchell) in earthen ponds. Aquac. Res. 2006, 37, 869–876. [Google Scholar] [CrossRef]
- IARC. Monographs on the Evaluation of Carcinogenic Risks to Humans: Chemical Agents and Related Occupations. A Review of Human Carcinogens; International Agency for Research on Cancer: Lyon, France, 2012. [Google Scholar]
- Van Doan, H.; Soltani, E.; Ingelbrecht, J.; Soltani, M. Medicinal herbs and plants: Potential treatment of monogenean infections in fish. Rev. Fish. Sci. Aquac. 2020, 28, 260–282. [Google Scholar] [CrossRef]
- Mladineo, I.; Trumbic, Z.; Ormad-Garcia, A.; Palenzuela, O.; Sitja-Bobadilla, A.; Manuguerra, S.; Ruiz, C.E.; Messina, C.M. In vitro testing of alternative synthetic and natural antiparasitic compounds against the monogenean Sparicotyle chrysophrii. Pathogens 2021, 10, 980. [Google Scholar] [CrossRef] [PubMed]
- Serra, B.N.V.; Martins, M.L.; Pereira, S.A.; Chagas, E.C.; Chaves, F.C.M.; Oliveira, M.I.B.; Jeronimo, G.T. Ovicidal effect of essential oils of Lippia alba, Lippia sidoides and Lippia gracilis on the acanthocephalan Neoechinorhynchus buttnerae (Eoacanthocephala: Neoechinorhynchidae). J. Fish Dis. 2022, 45, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; el Basuini, M.F.; Zaineldin, A.I.; Yilmaz, S.; Hasan, M.T.; Ahmadifar, E.; el Asely, A.M.; Abdel-Latif, H.M.R.; Alagawany, M.; Abu-Elala, N.M.; et al. Antiparasitic and antibacterial functionality of essential oils: An alternative approach for sustainable aquaculture. Pathogens 2021, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Lam, N.S.; Long, X.; Su, X.-Z.; Lu, F. Melaleuca alternifolia (tea tree) oil and its monoterpene constituents in treating protozoan and helminthic infections. Biomed. Pharmacother. 2020, 130, 110624. [Google Scholar] [CrossRef]
- Sutili, F.; Murari, A.; Silva, L.; Gressler, L.; Heinzmann, B.; de Vargas, A.; Schmidt, D.; Baldisserotto, B. The use of Ocimum americanum essential oil against the pathogens Aeromonas hydrophila and Gyrodactylus sp. in silver catfish (Rhamdia quelen). Lett. Appl. Microbiol. 2016, 63, 82–88. [Google Scholar] [CrossRef]
- Gonzales, A.; Yoshioka, E.T.O.; Mathews, P.D.; Mertins, O.; Chaves, F.C.M.; Videira, M.; Tavares-Dias, M. Anthelminthic efficacy of Cymbopogon citratus essential oil (Poaceae) against monogenean parasites of Colossoma macropomum (Serrasalmidae), and blood and histopathological effects. Aquaculture 2020, 528, 735500. [Google Scholar] [CrossRef]
- Luz, J.G.R.; Nogueira, J.N.; Alves, C.M.G.; Videira, M.N.; Canuto, K.M.; Castro, K.N.C.; Tavares-Dias, M. Essential oil of Alpinia zerumbet (Zingiberaceae) has anthelmintic efficacy against monogenean of Colossoma macropomum (Characiformes: Serrasalmidae). Aquac. Res. 2021, 52, 5340–5349. [Google Scholar] [CrossRef]
- Alves, C.M.G.; Nogueira, J.N.; Luz, J.G.R.; Chaves, F.C.M.; Tavares-Dias, M. Essential oil of Piper callosum, Piper hispidum and Piper marginatum (Piperaceae) possesses in vitro efficacy against monogeneans of Colossoma macropomum (tambaqui). Aquac. Res. 2021, 52, 6107–6116. [Google Scholar] [CrossRef]
- Zhou, S.; Dong, J.; Liu, Y.; Yang, Q.; Xu, N.; Yang, Y.; Gu, Z.; Ai, X. Anthelmintic efficacy of 35 herbal medicines against a monogenean parasite, Gyrodactylus kobayashii, infecting goldfish (Carassius auratus). Aquaculture 2020, 521, 734992. [Google Scholar] [CrossRef]
- Tu, X.; Duan, C.X.; Wu, S.W.; Fu, S.L.; Ye, J.M. Identification of plumbagin as an effective chemotherapeutic agent for treatment of Gyrodactylus infections. Aquaculture 2021, 535, 8. [Google Scholar] [CrossRef]
- FAO. Fishery and Aquaculture Statistics 2018; FAO Yearbook; FAO: Rome, Italy, 2020. [Google Scholar]
- Leal, J.F.; Neves, M.D.G.P.M.S.; Santos, E.; Esteves, V.I. Use of formalin in intensive aquaculture: Properties, application and effects on fish and water quality. Rev. Aquac. 2018, 10, 281–295. [Google Scholar] [CrossRef]
- Parikh, L.; Agindotan, B.O.; Burrows, M.E. Antifungal activity of plant-derived essential oils on pathogens of pulse crops. Plant Dis. 2021, 105, 1692–1701. [Google Scholar] [CrossRef] [PubMed]
- Ajith, M.; Pankaj; Shakil, N.A.; Kaushik, P.; Rana, V.S. Chemical composition and nematicidal activity of essential oils and their major compounds against Meloidogyne graminicola (Rice Root-Knot Nematode). J. Essent. Oil Res. 2020, 32, 526–535. [Google Scholar] [CrossRef]
- Sinha, S.; Biswas, D.; Mukherjee, A. Antigenotoxic and antioxidant activities of palmarosa and citronella essential oils. J. Ethnopharmacol. 2011, 137, 1521–1527. [Google Scholar] [CrossRef]
- Nirmal, S.A.; Girme, A.S.; Bhalke, R.D. Major constituents and anthelmintic activity of volatile oils from leaves and flowers of Cymbopogon martini Roxb. Nat. Prod. Res. 2007, 21, 1217–1220. [Google Scholar] [CrossRef]
- Rana, V.S.; Das, M.; Blazqeuz, M.A. Essential Oil Yield, Chemical Composition, and Total Citral Content of Nine Cultivars of Cymbopogon Species from Western India. J. Herbs Spices Med. Plants 2016, 22, 289–299. [Google Scholar] [CrossRef]
- Kumar, C.B.; Kumar, A.; Rathore, G. Antibacterial activity of palmarosa oil significantly varies between Aeromonas veronii and Aeromonas caviae and exhibits selective action on tetracycline and sulfonamide resistant A. caviae. J. Appl. Microbiol. 2022, 132, 4321–4329. [Google Scholar] [CrossRef]
- Cable, J.; Harris, P.D. Gyrodactylid developmental biology: Historical review, current status and future trends. Int. J. Parasitol. 2002, 32, 255–280. [Google Scholar] [CrossRef]
- Saccol, E.M.H.; Toni, C.; Pes, T.S.; Ourique, G.M.; Gressler, L.T.; Silva, L.V.F.; Mourao, R.H.V.; Oliveira, R.B.; Baldisserotto, B.; Pavanato, M.A. Anaesthetic and antioxidant effects of Myrcia sylvatica (G. Mey.) DC. and Curcuma longa L. essential oils on tambaqui (Colossoma macropomum). Aquac. Res. 2017, 48, 2012–2031. [Google Scholar] [CrossRef]
- Teles, A.M.; Rosa, T.D.D.; Mouchrek, A.N.; Abreu-Silva, A.L.; Calabrese, K.D.S.; Almeida-Souza, F. Cinnamomum zeylanicum, Origanum vulgare, and Curcuma longa essential oils: Chemical composition, antimicrobial and antileishmanial activity. Evid.-Based Complement. Altern. Med. 2019, 2019, 2421695. [Google Scholar] [CrossRef] [Green Version]
- Hutson, K.S.; Brazenor, A.K.; Vaughan, D.B.; Trujillo-Gonzalez, A. Monogenean parasite cultures: Current techniques and recent advances. Adv. Parasitol. 2018, 99, 61–91. [Google Scholar]
- Truong Dinh, H. Reproductive strategies of parasitic flatworms (Platyhelminthes, Monogenea): The impact on parasite management in aquaculture. Aquac. Int. 2020, 28, 421–447. [Google Scholar]
- Le, T.B.; Beaufay, C.; Nghiem, D.T.; Pham, T.A.; Mingeot-Leclercq, M.P.; Quetin-Leclercq, J. Evaluation of the anti-trypanosomal activity of vietnamese essential oils, with emphasis on Curcuma longa L. and its components. Molecules 2019, 24, 1158. [Google Scholar] [CrossRef] [Green Version]
- Ajaiyeoba, E.O.; Sama, W.; Essien, E.E.; Olayemi, J.O.; Ekundayo, O.; Walker, T.M.; Setzer, W.N. Larvicidal activity of turmerone-rich essential oils of Curcuma longa leaf and rhizome from nigeria on Anopheles gambiae. Pharm. Biol. 2008, 46, 279–282. [Google Scholar] [CrossRef]
- Setzer, W.; Duong, L.; Poudel, A.; Mentreddy, S. Variation in the Chemical Composition of Five Varieties of Curcuma longa Rhizome Essential Oils Cultivated in North Alabama. Foods 2021, 10, 212. [Google Scholar] [CrossRef]
- Schmahl, G.; Mehlhorn, H. Treatment of fish parasites. 4. effects of sym. triazinone (toltrazuril) on monogenean. Parasitol. Res. 1988, 75, 132–143. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; El Basuini, M.F.; Yilmaz, S.; Abdel-Latif, H.M.R.; Alagawany, M.; Kari, Z.A.; Razab, M.K.A.A.; Hamid, N.K.A.; Moonmanee, T.; Van Doan, H. Exploring the Roles of Dietary Herbal Essential Oils in Aquaculture: A Review. Animals 2022, 12, 823. [Google Scholar] [CrossRef]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Liju, V.B.; Jeena, K.; Kuttan, R. Acute and subchronic toxicity as well as mutagenic evaluation of essential oil from turmeric (Curcuma longa L.). Food Chem. Toxicol. 2013, 53, 52–61. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Essential Oil | CAS | Source | Maximum Anthelmintic Efficacy (%) | CAE (mg/L) | CFD (mg/L) |
|---|---|---|---|---|---|
| Palmarosa oil | 8014-19-5 | Cymbopogon martinii (Roxb.) Wats. | 100 | 10 | 30 |
| Curcuma oil | 8024-37-1 | Curcuma longa Linn. | 100 | 12 | 25 |
| Cablin patchouli oil | 8014-09-3 | Pogostemon cablin (Blanco) Benth. * | 100 | 10 | 10 |
| Zedoary oil | / | Curcuma zedoaria (Christm.) Rosc. | 100 | 15 | 20 |
| Rue oil | 8014-29-7 | Ruta graveolens Linn. * | 100 | 25 | 40 |
| Tea tree oil | 68647-73-4 | Melaleuca alternifolia Cheel | 100 | 40 | 50 |
| Neem oil | / | Melia azedarace Linn. | 100 | 40 | 45 |
| Cassia oil | 8015-96-1 | Cinnamomum cassia Presl | 94.57 | 14 | 14 |
| Eucalyptus oil | 8000-48-4 | Eucalyptus globulus Labill. * | 90.84 | 100 | 100 |
| Clove oil | 8000-34-8 | Syzygium aromaticum (L.) Merr. Et Perry | 45.06 | 16 | 16 |
| Clove leaf oil | 8015-97-2 | Syzygium aromaticum (L.) Merr. Et Perry | 36.02 | 18 | 18 |
| Origanum oil | 8007-11-2 | Origanum vulgare Linn. | 23.36 | 10 | 10 |
| Anise oil | 8007-70-3 | Pimpinella anisum Linn. | 21.92 | 10 | 10 |
| Peppermint Oil | 68917-18-0 | Mentha sachalinensis (Briq. ex Miyaabe et Miyake) Kudo * | 10.45 | 40 | 40 |
| Fennel Oil | 8006-84-6 | Foeniculum vulgare Mill. | 10.1 | 40 | 40 |
| Lemon oil | 84929-31-7 | Citrus limon (L.) Burm. F. | 5.62 | 10 | 10 |
| Essential Oil | Exposure Time (h) | EC50 (95%CI, mg/L) | EC90 (95%CI, mg/L) |
|---|---|---|---|
| Palmarosa oil | 2 | 9.87 (9.24–10.72) | 15.4 (13.92–17.7) |
| 24 | 4.98 (4.11–5.66) | 8.07 (7.3–9.22) | |
| Curcuma oil | 2 | 3.48 (0–5.21) | 6.7 (4.97–10.84) |
| 24 | 5.72 (4.14–6.86) | 9.34 (8.02–12.17) |
| Essential Oil | Concentration (mg/L) | No. of Fish/Tank | No. Dead Fish (mean ± SD) | LC50 (95%CI, mg/L) | ||
|---|---|---|---|---|---|---|
| 24 h | 48 h | 24 h | 48 h | |||
| Palmarosa oil | 0 | 10 | 0 | 0 | 40.8 (38.26–43.43) | 39.15 (36.36–41.86) |
| 30 | 10 | 0 | 0.33 ± 0.58 | |||
| 35 | 10 | 1.67 ± 1.58 | 2.67 ± 0.58 | |||
| 40 | 10 | 4.33 ± 0.58 | 5.67 ± 1.15 | |||
| 45 | 10 | 7.33 ± 0.58 | 8.0 ± 1.0 | |||
| 50 | 10 | 10 | 10 | |||
| Curcuma oil | 0 | 10 | 0 | 0 | 31.73 (29.2–34.51) | 28.85 (26.06–31.5) |
| 20 | 10 | 0 | 0 | |||
| 25 | 10 | 1.67 ± 0.58 | 3.57 ± 0.58 | |||
| 30 | 10 | 3.0 ± 1.0 | 5.33 ± 0.58 | |||
| 35 | 10 | 6.67 ± 0.58 | 8.33 ± 0.58 | |||
| 40 | 10 | 10 | 10 | |||
| Treatment | 0.2% DMSO | Palmarosa Oil | Curcuma Oil |
|---|---|---|---|
| RBC (1012/L) | 0.98 ± 0.12 a | 0.94 ± 0.15 a | 1.16 ± 0.28 a |
| Hb (g/L) | 105 ± 18.01 a | 103.6 ± 10.24 a | 109.2 ± 13.08 a |
| HCT (%) | 16.18 ± 2.32 a | 15.12 ± 2.24 a | 15.52 ± 1.58 a |
| MCV (fL) | 179.2 ± 8.09 a | 177.96 ± 8.9 a | 176.16 ± 12.25 a |
| MCH (pg) | 127.94 ± 4.9 a | 128.06 ± 12.64 a | 133.5 ± 11.9 a |
| MCHC (g/L) | 667.8 ± 62.78 a | 747.4 ± 87.99 a | 731 ± 50.4 a |
| PLT (109/L) | 47.8 ± 5.89 a | 58 ± 17.25 a | 43.8 ± 18.75 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Yang, Q.; Dong, J.; Liu, Y.; Xu, N.; Yang, Y.; Ai, X. Anthelmintic Efficacy of Palmarosa Oil and Curcuma Oil against the Fish Ectoparasite Gyrodactylus kobayashii (monogenean). Animals 2022, 12, 1685. https://doi.org/10.3390/ani12131685
Zhou S, Yang Q, Dong J, Liu Y, Xu N, Yang Y, Ai X. Anthelmintic Efficacy of Palmarosa Oil and Curcuma Oil against the Fish Ectoparasite Gyrodactylus kobayashii (monogenean). Animals. 2022; 12(13):1685. https://doi.org/10.3390/ani12131685
Chicago/Turabian StyleZhou, Shun, Qiuhong Yang, Jing Dong, Yongtao Liu, Ning Xu, Yibin Yang, and Xiaohui Ai. 2022. "Anthelmintic Efficacy of Palmarosa Oil and Curcuma Oil against the Fish Ectoparasite Gyrodactylus kobayashii (monogenean)" Animals 12, no. 13: 1685. https://doi.org/10.3390/ani12131685
APA StyleZhou, S., Yang, Q., Dong, J., Liu, Y., Xu, N., Yang, Y., & Ai, X. (2022). Anthelmintic Efficacy of Palmarosa Oil and Curcuma Oil against the Fish Ectoparasite Gyrodactylus kobayashii (monogenean). Animals, 12(13), 1685. https://doi.org/10.3390/ani12131685