
Carboxylated ε-Poly-L-Lysine Supplementation of the Freezing Extender Improves the Post-Thawing Boar Sperm Quality

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Extenders
2.2. Semen Collection
2.3. Semen Processing
2.4. Evaluation of Post-Thaw Sperm Motility
2.5. Evaluation of Sperm Membrane Integrity and Acrosome Integrity
2.6. Mitochondrial Membrane Potential
2.7. Measurement of NADH-CoQ Activity
2.8. Measurement of Sperm ATP Level
2.9. Measurement of Post-Thaw Sperm MDA Content
2.10. Evaluation of Activities of CAT, SOD, and GPx in Post-Thaw Sperm
2.11. Annexin V-FITC/PI Assay
2.12. Statistical Analysis
3. Results
3.1. Addition of Carboxylated ε-Poly-L-Lysine to the Freezing Medium Increased the Post-Thaw Sperm Motility Parameters
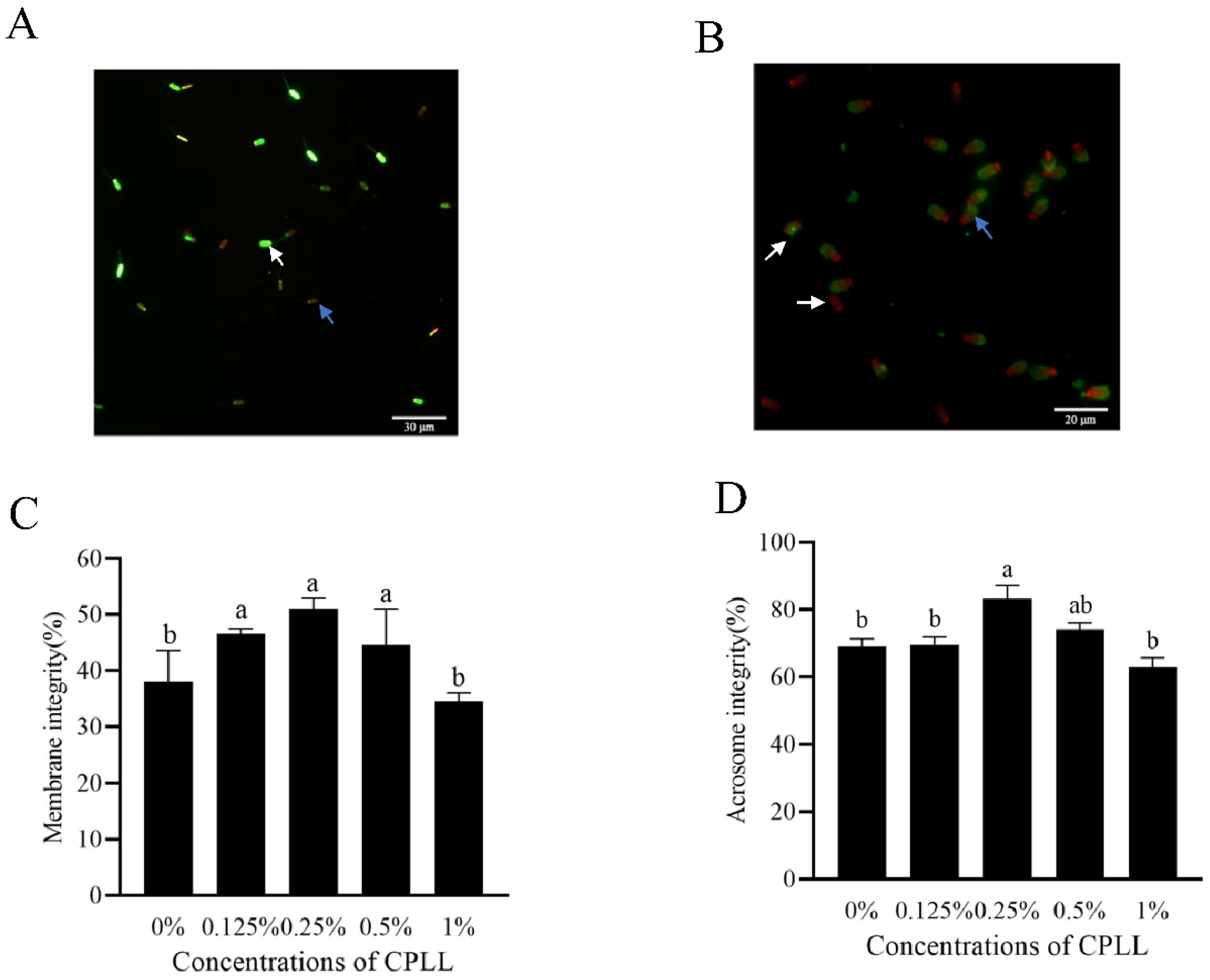
3.2. Addition of Carboxylated ε-Poly-L-Lysine to the Freezing Medium Improved the Post-Thaw Sperm Membrane Integrity and Acrosome Integrity
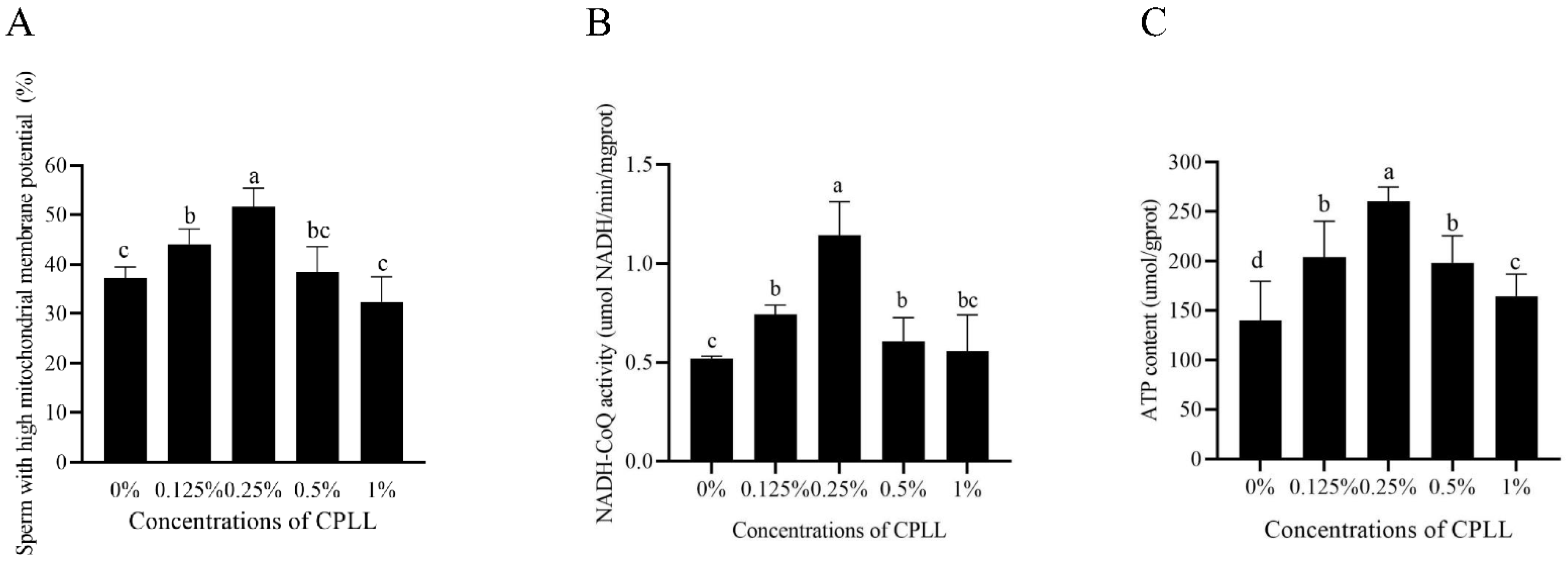
3.3. Addition of Carboxylated ε-Poly-L-Lysine to the Freezing Medium Improved the Post-Thaw Sperm Mitochondrial Function
3.4. Addition of Carboxylated ε-Poly-L-Lysine to the Freezing Medium Increased the Post-Thaw Sperm ATP Level
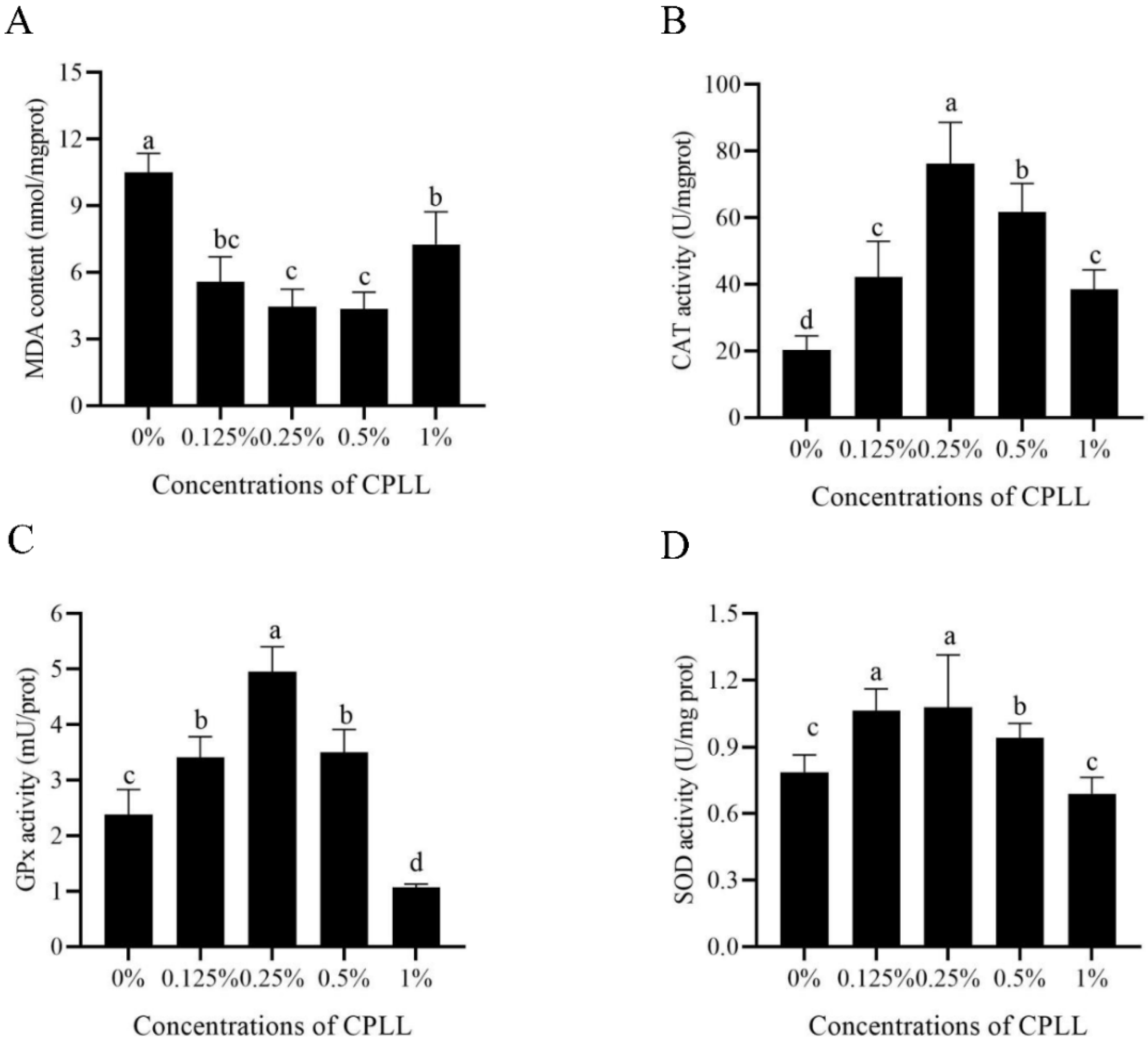
3.5. Addition of Carboxylated ε-Poly-L-Lysine to the Freezing Medium Improved the Post-Thaw Sperm Antioxidative Stress
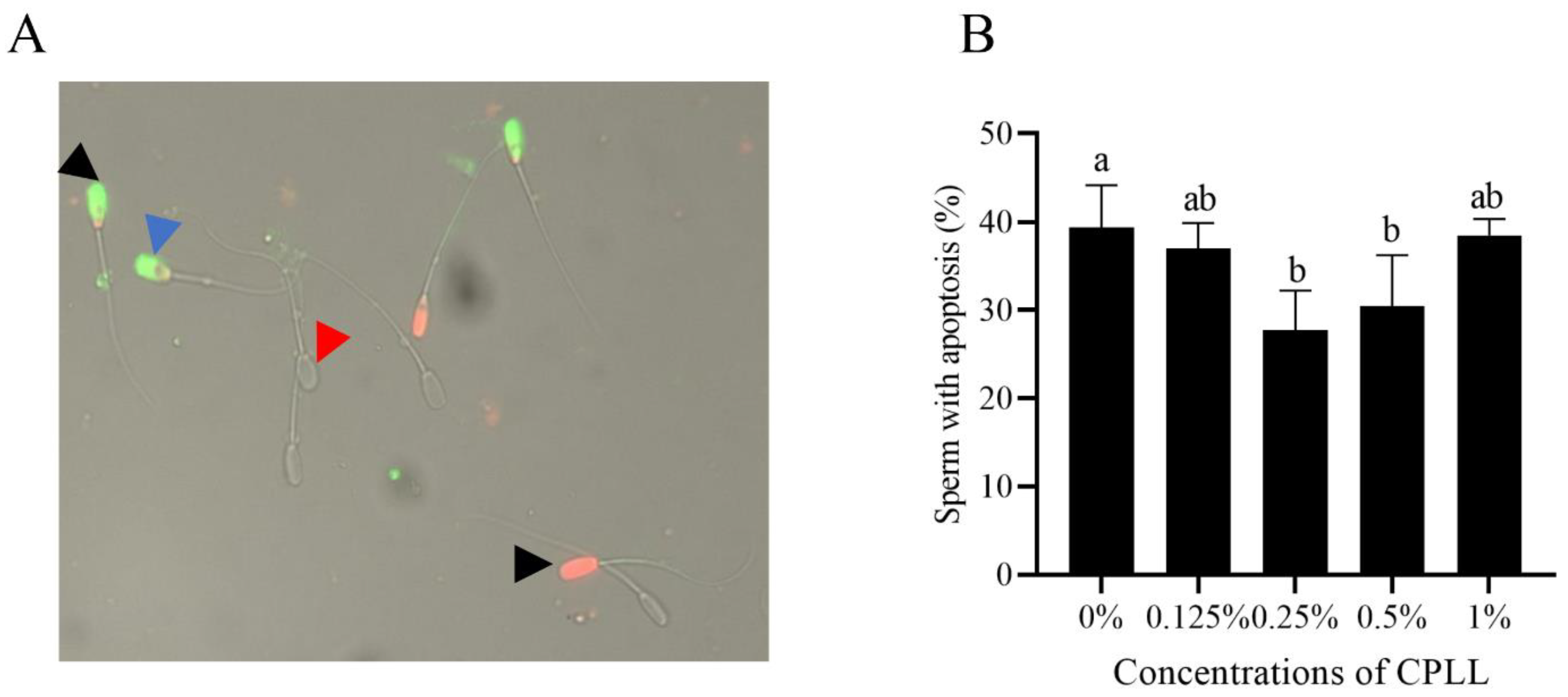
3.6. Addition of Carboxylated ε-Poly-L-Lysine to the Freezing Medium Reduced the Post-Thaw Sperm Apoptosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Gil, J.E.; Estrada, E. Artificial Insemination in Boar Reproduction. In Boar Reproduction: Fundamentals and New Biotechnological Trends; Bonet, S., Casas, I., Holt, W.V., Yeste, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 589–607. [Google Scholar]
- Knox, R.V. Artificial insemination in pigs today. Theriogenology 2016, 85, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M. Recent Advances in Boar Sperm Cryopreservation: State of the Art and Current Perspectives. Reprod. Domest. Anim. 2015, 50 (Suppl. 2), 71–79. [Google Scholar] [CrossRef] [PubMed]
- Didion, B.A.; Braun, G.D.; Duggan, M.V. Field fertility of frozen boar semen: A retrospective report comprising over 2600 AI services spanning a four year period. Anim. Reprod. Sci. 2013, 137, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Pursel, V.G.; Schulman, L.L.; Johnson, L.A. Distribution and morphology of fresh and frozen-thawed sperm in the reproductive tract of gilts after artificial insemination. Biol. Reprod. 1978, 19, 69–76. [Google Scholar] [CrossRef]
- Di Santo, M.; Tarozzi, N.; Nadalini, M.; Borini, A. Human Sperm Cryopreservation: Update on Techniques, Effect on DNA Integrity, and Implications for ART. Adv. Urol. 2012, 2012, 854837. [Google Scholar] [CrossRef]
- Maxwell, W.M.; Johnson, L.A. Membrane status of boar spermatozoa after cooling or cryopreservation. Theriogenology 1997, 48, 209–219. [Google Scholar] [CrossRef]
- Vadnais, M.L.; Althouse, G.C. Characterization of capacitation, cryoinjury, and the role of seminal plasma in porcine sperm. Theriogenology 2011, 76, 1508–1516. [Google Scholar] [CrossRef]
- Zhu, Z.; Li, R.; Fan, X.; Lv, Y.; Zheng, Y.; Hoque, S.A.M.; Wu, D.; Zeng, W. Resveratrol Improves Boar Sperm Quality via 5’AMP-Activated Protein Kinase Activation during Cryopreservation. Oxid. Med. Cell Longev. 2019, 2019, 5921503. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.H.; Kim, W.H.; Cheong, H.T.; Yang, B.K.; Park, C.K. Effect of Alpha-Linolenic Acid with Bovine Serum Albumin or Methyl-Beta-Cyclodextrin on Membrane Integrity and Oxidative Stress of Frozen-Thawed Boar Sperm. Dev. Reprod. 2019, 23, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, T.; Yoshida, S.; Teshima, H.; Shimada, M. The addition of calcium ion chelator, EGTA to thawing solution improves fertilizing ability in frozen-thawed boar sperm. Anim. Sci. J. 2011, 82, 412–419. [Google Scholar] [CrossRef]
- Robles, V.; Valcarce, D.G.; Riesco, M.F. The Use of Antifreeze Proteins in the Cryopreservation of Gametes and Embryos. Biomolecules 2019, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, T.; Shimada, M. New strategies of boar sperm cryopreservation: Development of novel freezing and thawing methods with a focus on the roles of seminal plasma. Anim. Sci. J. 2012, 83, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, W.; Li, R.; Zeng, W. Reducing the Glucose Level in Pre-treatment Solution Improves Post-thaw Boar Sperm Quality. Front. Vet. Sci. 2022, 9, 856536. [Google Scholar] [CrossRef] [PubMed]
- Fowler, A.; Toner, M. Cryo-injury and biopreservation. Ann. N. Y. Acad. Sci. 2005, 1066, 119–135. [Google Scholar] [CrossRef]
- Buhr, M.M.; Fiser, P.; Bailey, J.L.; Curtis, E.F. Cryopreservation in different concentrations of glycerol alters boar sperm and their membranes. J. Androl. 2001, 22, 961–969. [Google Scholar] [CrossRef]
- El Assal, R.; Abou-Elkacem, L.; Tocchio, A.; Pasley, S.; Matosevic, S.; Kaplan, D.L.; Zylberberg, C.; Demirci, U. Bioinspired Preservation of Natural Killer Cells for Cancer Immunotherapy. Adv. Sci. 2019, 6, 1802045. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Lee, H.R.; Yu, S.J.; Han, M.E.; Lee, D.Y.; Kim, S.Y.; Ahn, H.J.; Han, M.J.; Lee, T.I.; Kim, T.S.; et al. Hydrogel-laden paper scaffold system for origami-based tissue engineering. Proc. Natl. Acad. Sci. USA 2015, 112, 15426–15431. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, K.; Hyon, S.H. Polyampholytes as low toxic efficient cryoprotective agents with antifreeze protein properties. Biomaterials 2009, 30, 4842–4849. [Google Scholar] [CrossRef]
- Matsumura, K.; Bae, J.Y.; Kim, H.H.; Hyon, S.H. Effective vitrification of human induced pluripotent stem cells using carboxylated epsilon-poly-l-lysine. Cryobiology 2011, 63, 76–83. [Google Scholar] [CrossRef]
- Fujikawa, T.; Ando, T.; Gen, Y.; Hyon, S.H.; Kubota, C. Cryopreservation of bovine somatic cells using antifreeze polyamino-acid (carboxylated poly-L-lysine). Cryobiology 2017, 76, 140–145. [Google Scholar] [CrossRef]
- Kamoshita, M.; Kato, T.; Fujiwara, K.; Namiki, T.; Matsumura, K.; Hyon, S.H.; Ito, J.; Kashiwazaki, N. Successful vitrification of pronuclear-stage pig embryos with a novel cryoprotective agent, carboxylated epsilon-poly-L-lysine. PLoS ONE 2017, 12, e0176711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tariq, A.; Ahmad, M.; Iqbal, S.; Riaz, M.I.; Tahir, M.Z.; Ghafoor, A.; Riaz, A. Effect of carboxylated poly l-Lysine as a cryoprotectant on post-thaw quality and in vivo fertility of Nili Ravi buffalo (Bubalus bubalis) bull semen. Theriogenology 2020, 144, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, R.; Feng, C.; Liu, R.; Zheng, Y.; Hoque, S.A.M.; Wu, D.; Lu, H.; Zhang, T.; Zeng, W. Exogenous Oleic Acid and Palmitic Acid Improve Boar Sperm Motility via Enhancing Mitochondrial Beta-Oxidation for ATP Generation. Animals 2020, 10, 591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Li, R.; Lv, Y.; Zeng, W. Melatonin protects rabbit spermatozoa from cryo-damage via decreasing oxidative stress. Cryobiology 2019, 88, 1–8. [Google Scholar] [CrossRef]
- Bwanga, C.O. Cryopreservation of boar semen. I: A literature review. Acta Vet. Scand. 1991, 32, 431–453. [Google Scholar] [CrossRef]
- Knox, R.V. The Fertility of Frozen Boar Sperm When used for Artificial Insemination. Reprod. Domest. Anim. 2015, 50 (Suppl. 2), 90–97. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, T.; Abe, S.; Shimada, M. Improved conception rates in sows inseminated with cryopreserved boar spermatozoa prepared with a more optimal combination of osmolality and glycerol in the freezing extender. Anim. Sci. J. 2009, 80, 121–129. [Google Scholar] [CrossRef]
- Okazaki, T.; Akiyoshi, T.; Kan, M.; Mori, M.; Teshima, H.; Shimada, M. Artificial insemination with seminal plasma improves the reproductive performance of frozen-thawed boar epididymal spermatozoa. J. Androl. 2012, 33, 990–998. [Google Scholar] [CrossRef]
- Polge, C.; Smith, A.U.; Parkes, A.S. Revival of spermatozoa after vitrification and dehydration at low temperatures. Nature 1949, 164, 666. [Google Scholar] [CrossRef]
- Kim, S.; Lee, Y.J.; Ji, D.B.; Kim, Y.J. Evaluation of different cryoprotectants (CPAs) in boar semen cryopreservation. J. Vet. Med. Sci. 2011, 73, 961–963. [Google Scholar] [CrossRef] [Green Version]
- Athurupana, R.; Takahashi, D.; Ioki, S.; Funahashi, H. Trehalose in glycerol-free freezing extender enhances post-thaw survival of boar spermatozoa. J. Reprod. Dev. 2015, 61, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Martinez, H.; Wallgren, M. Advances in boar semen cryopreservation. Vet. Med. Int. 2010, 2010, 396181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorontsov, D.A.; Sazaki, G.; Hyon, S.H.; Matsumura, K.; Furukawa, Y. Antifreeze effect of carboxylated epsilon-poly-L-lysine on the growth kinetics of ice crystals. J. Phys. Chem. B 2014, 118, 10240–10249. [Google Scholar] [CrossRef] [PubMed]
- Rajan, R.; Hayashi, F.; Nagashima, T.; Matsumura, K. Toward a Molecular Understanding of the Mechanism of Cryopreservation by Polyampholytes: Cell Membrane Interactions and Hydrophobicity. Biomacromolecules 2016, 17, 1882–1893. [Google Scholar] [CrossRef]
- Jain, M.; Rajan, R.; Hyon, S.H.; Matsumura, K. Hydrogelation of dextran-based polyampholytes with cryoprotective properties via click chemistry. Biomater. Sci. 2014, 2, 308–317. [Google Scholar] [CrossRef]
- Suarez, S.S.; Pacey, A.A. Sperm transport in the female reproductive tract. Hum. Reprod. Update 2006, 12, 23–37. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, T.; Imamura, S.; Tokumaru, M.; Ando, T.; Gen, Y.; Hyon, S.H.; Kubota, C. Cryoprotective effect of antifreeze polyamino-acid (Carboxylated Poly-l-Lysine) on bovine sperm: A technical note. Cryobiology 2018, 82, 159–162. [Google Scholar] [CrossRef]
- Amaral, A.; Ramalho-Santos, J.; St John, J.C. The expression of polymerase gamma and mitochondrial transcription factor A and the regulation of mitochondrial DNA content in mature human sperm. Hum. Reprod. 2007, 22, 1585–1596. [Google Scholar] [CrossRef]
- St John, J.C.; Jokhi, R.P.; Barratt, C.L. The impact of mitochondrial genetics on male infertility. Int. J. Androl. 2005, 28, 65–73. [Google Scholar] [CrossRef]
- Amaral, A.; Lourenco, B.; Marques, M.; Ramalho-Santos, J. Mitochondria functionality and sperm quality. Reproduction 2013, 146, R163–R174. [Google Scholar] [CrossRef] [Green Version]
- Ramio-Lluch, L.; Fernandez-Novell, J.M.; Pena, A.; Colas, C.; Cebrian-Perez, J.A.; Muino-Blanco, T.; Ramirez, A.; Concha, II; Rigau, T.; Rodriguez-Gil, J.E. ‘In vitro’ capacitation and acrosome reaction are concomitant with specific changes in mitochondrial activity in boar sperm: Evidence for a nucleated mitochondrial activation and for the existence of a capacitation-sensitive subpopulational structure. Reprod. Domest. Anim. 2011, 46, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Umehara, T.; Okazaki, T.; Goto, M.; Fujita, Y.; Hoque, S.A.M.; Kawai, T.; Zeng, W.; Shimada, M. Gene Expression and Protein Synthesis in Mitochondria Enhance the Duration of High-Speed Linear Motility in Boar Sperm. Front. Physiol. 2019, 10, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Fan, X.; Lv, Y.; Lin, Y.; Wu, D.; Zeng, W. Glutamine protects rabbit spermatozoa against oxidative stress via glutathione synthesis during cryopreservation. Reprod. Fertil. Dev. 2017, 29, 2183–2194. [Google Scholar] [CrossRef] [PubMed]
- Storey, B.T. Mammalian sperm metabolism: Oxygen and sugar, friend and foe. Int. J. Dev. Biol. 2008, 52, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Zeng, Y.; Zeng, W. Cysteine improves boar sperm quality via glutathione biosynthesis during the liquid storage. Anim. Biosci. 2022, 35, 166–176. [Google Scholar] [CrossRef]
- Bansal, A.K.; Bilaspuri, G.S. Impacts of oxidative stress and antioxidants on semen functions. Vet. Med. Int. 2010, 2010, 686137. [Google Scholar] [CrossRef] [Green Version]
- Trzcinska, M.; Bryla, M. Apoptotic-like changes of boar spermatozoa in freezing media supplemented with different antioxidants. Pol. J. Vet. Sci. 2015, 18, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Ofosu, J.; Qazi, I.H.; Fang, Y.; Zhou, G.B. Use of melatonin in sperm cryopreservation of farm animals: A brief review. Anim. Reprod. Sci. 2021, 233, 106850. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sperm Parameters | 0% | 0.125% | 0.25% | 0.5% | 1% |
|---|---|---|---|---|---|
| Total motility (%) | 36.2 ± 3.6 b | 34.4 ± 4.0 b | 58.2 ± 5.0 a | 53.2 ± 2.1 a | 41.5 ± 5.3 ab |
| Progressive motility (%) | 17.7 ± 1.2 c | 19.2 ± 2.7 c | 43.0 ± 4.3 a | 32.8 ± 3.2 b | 23.3 ± 3.5 c |
| VCL (μm/s) | 117.4 ± 9.1 b | 115.4 ± 4.6 b | 144.4 ± 4.4 a | 139.3± 3.0 a | 125.3 ± 8.0 b |
| VSL (μm/s) | 32.6 ± 3.3 b | 28.5 ± 1.8 b | 48.7± 3.0 a | 50.1 ± 2.3 a | 37.9 ± 5.1 b |
| VAP (μm/s) | 61.2 ± 4.5 b | 64.3 ± 3.8 b | 75.1 ± 2.1 a | 74.0 ± 0.8 a | 67.8 ± 4.6 ab |
| BCF (Hz) | 24.7 ± 1.2 | 24.5 ± 1.4 | 26.4 ± 0.9 | 24.9 ± 0.6 | 24.2 ± 1.2 |
| ALH (μm) | 8.7 ± 0.6 | 8.9 ± 0.3 | 8.3 ± 0.4 | 8.2 ± 0.3 | 8.4 ± 0.3 |
| STR (%) | 47.5 ± 2.0 b | 44.8 ± 1.3 b | 63.6 ± 3.3 a | 66.4 ± 2.6 a | 54.2 ± 4.2 ab |
| LIN (%) | 26.1 ± 1.0 b | 25.1 ± 1.0 b | 33.8 ± 2.1 a | 36.1 ± 1.9 a | 29.8 ± 2.1 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Li, Y.; Zhu, Z. Carboxylated ε-Poly-L-Lysine Supplementation of the Freezing Extender Improves the Post-Thawing Boar Sperm Quality. Animals 2022, 12, 1726. https://doi.org/10.3390/ani12131726
Zhang W, Li Y, Zhu Z. Carboxylated ε-Poly-L-Lysine Supplementation of the Freezing Extender Improves the Post-Thawing Boar Sperm Quality. Animals. 2022; 12(13):1726. https://doi.org/10.3390/ani12131726
Chicago/Turabian StyleZhang, Weijing, Yajing Li, and Zhendong Zhu. 2022. "Carboxylated ε-Poly-L-Lysine Supplementation of the Freezing Extender Improves the Post-Thawing Boar Sperm Quality" Animals 12, no. 13: 1726. https://doi.org/10.3390/ani12131726
APA StyleZhang, W., Li, Y., & Zhu, Z. (2022). Carboxylated ε-Poly-L-Lysine Supplementation of the Freezing Extender Improves the Post-Thawing Boar Sperm Quality. Animals, 12(13), 1726. https://doi.org/10.3390/ani12131726