

Effect of Treating Eggs with Coenzyme Q10 (CoQ10) on Growth Variables, Histomorphometry, and Antioxidant Capacity in Red Tilapia (Oreochromis aureus × Oreochromis mossambicus) Larvae


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Broodstock and Eggs Maintenance
2.2. Experimental Exposure of Fertilized Eggs to CoQ10
2.3. Growth Metrics
2.4. Histological and Morphometric Analysis
2.5. Antioxidant Potential
2.6. Statistical Analysis
3. Results
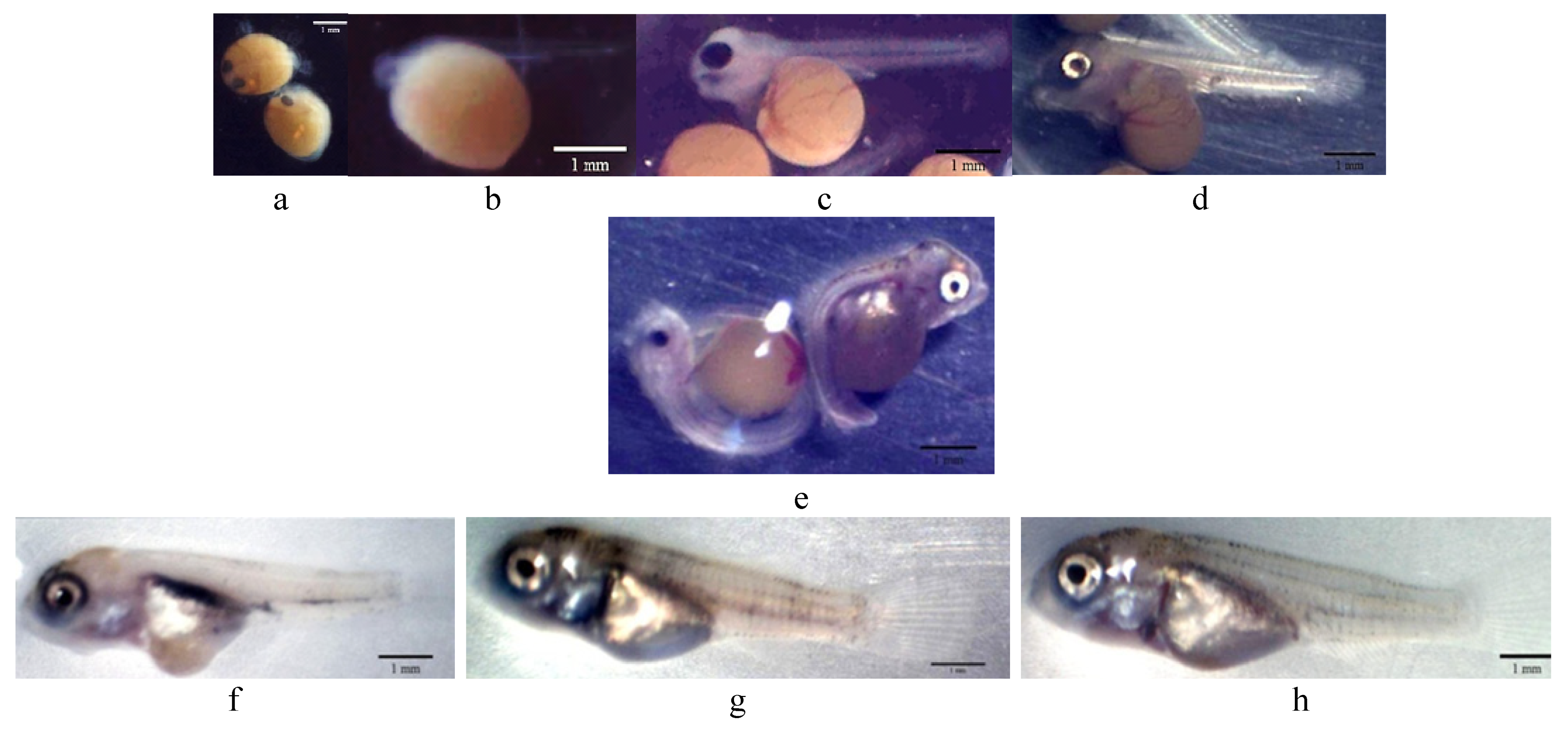
3.1. Hatchability Rate, Growth Development, and Malformation Interpretation
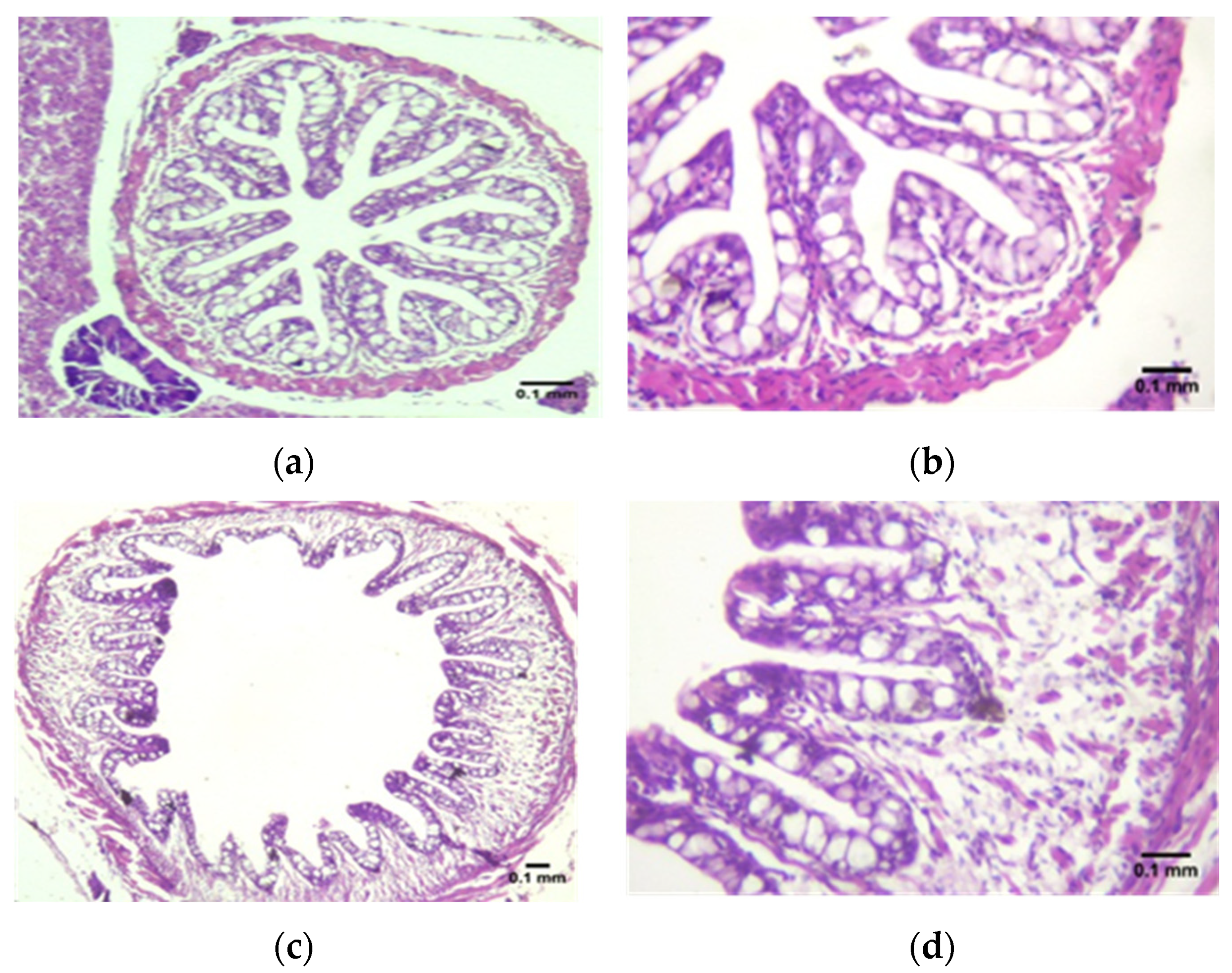
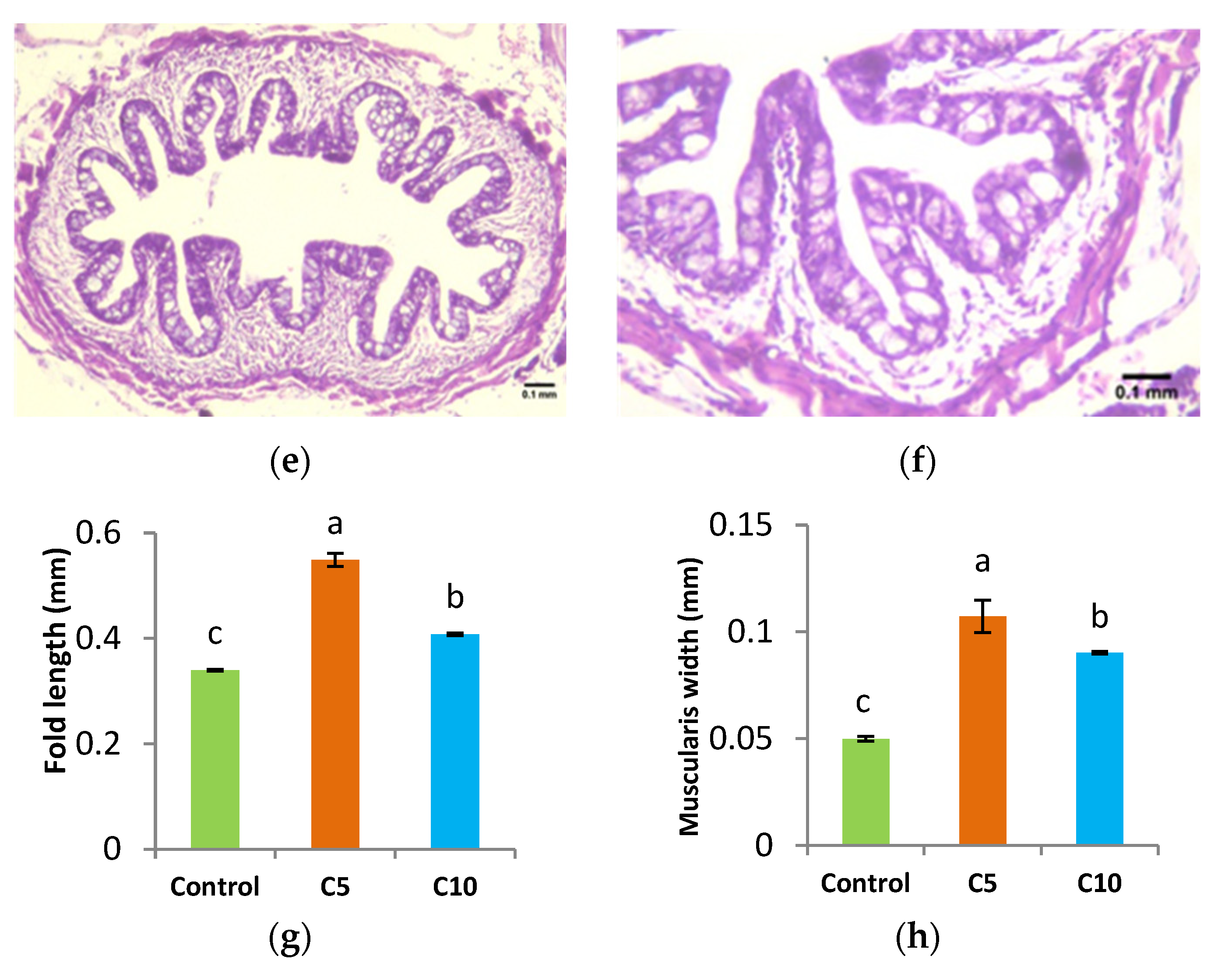
3.2. Histological Assessment
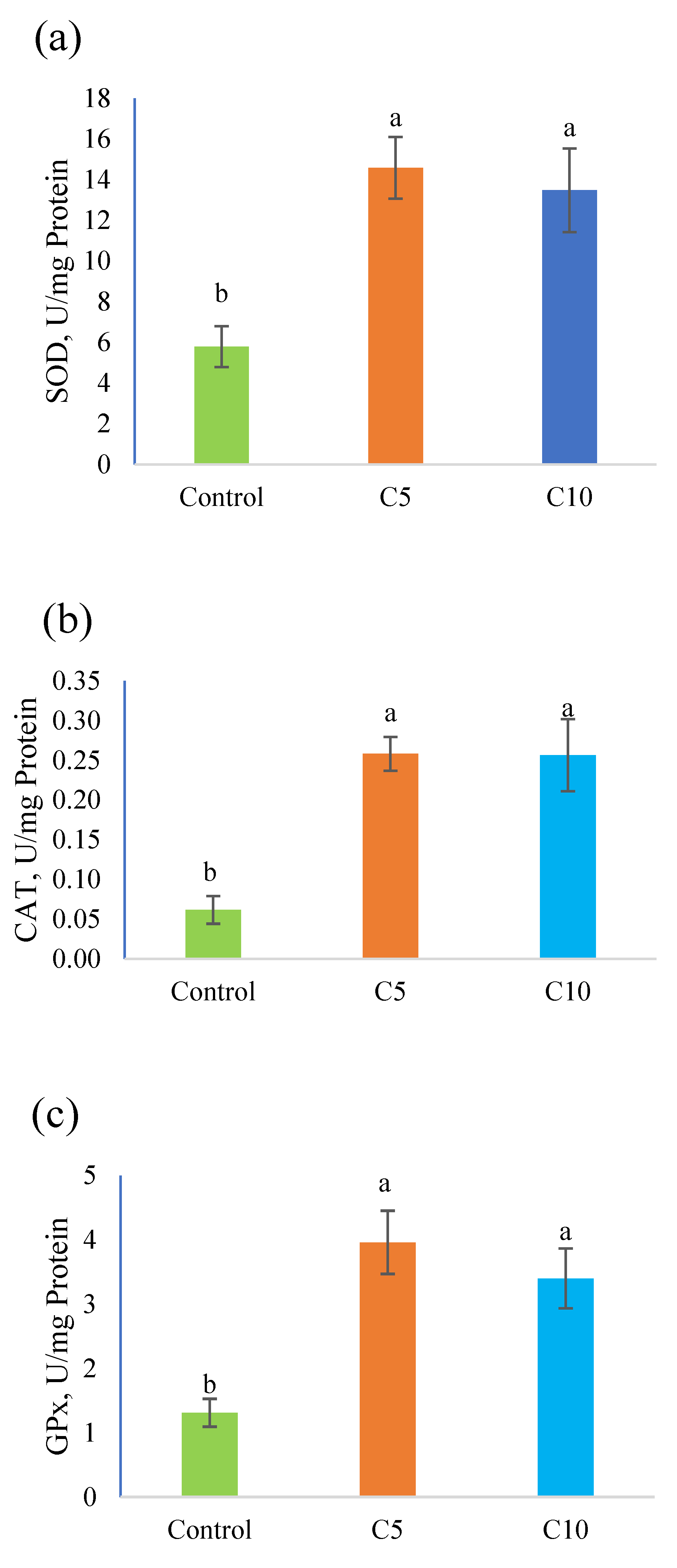
3.3. Antioxidant Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaki, M.A.A.; Alabssawy, A.N.; Nour, A.E.-A.M.; El Basuini, M.F.; Dawood, M.A.O.; Alkahtani, S.; Abdel-Daim, M.M. The impact of stocking density and dietary carbon sources on the growth, oxidative status and stress markers of Nile tilapia (Oreochromis niloticus) reared under biofloc conditions. Aquac. Rep. 2020, 16, 100282. [Google Scholar] [CrossRef]
- Cámara-Ruiz, M.; Santo, C.E.; Gessner, J.; Wuertz, S. How to improve foraging efficiency for restocking measures of juvenile Baltic sturgeon (Acipenser oxyrinchus). Aquaculture 2019, 502, 12–17. [Google Scholar] [CrossRef]
- Bentzon-Tilia, M.; Sonnenschein, E.C.; Gram, L. Monitoring and managing microbes in aquaculture—Towards a sustainable industry. Microb. Biotechnol. 2016, 9, 576–584. [Google Scholar] [CrossRef]
- Hannon, C.; Officer, R.A.; Le Dorven, J. Review of the technical challenges facing aquaculture of the European abalone Haliotis tuberculata in Ireland. Aquac. Int. 2013, 21, 243–254. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.; Chakraborty, T.; Kumar, V.; DeBoeck, G.; Mohanta, K.N. Aquaculture and stress management: A review of probiotic intervention. J. Anim. Physiol. Anim. Nutr. 2013, 97, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-B.; Li, J.-R.; Lin, J. Probiotics in aquaculture: Challenges and outlook. Aquaculture 2008, 281, 1–4. [Google Scholar] [CrossRef]
- Bobe, J. Egg quality in fish: Present and future challenges. Anim. Front. 2015, 5, 66–72. [Google Scholar] [CrossRef]
- Olafsen, J.A. Interactions between fish larvae and bacteria in marine aquaculture. Aquaculture 2001, 200, 223–247. [Google Scholar] [CrossRef]
- Lieke, T.; Steinberg, C.E.W.; Bittmann, S.; Behrens, S.; Hoseinifar, S.H.; Meinelt, T.; Knopf, K.; Kloas, W. Fulvic acid accelerates hatching and stimulates antioxidative protection and the innate immune response in zebrafish larvae. Sci. Total Environ. 2021, 796, 148780. [Google Scholar] [CrossRef]
- Teiba, I.; Okunishi, S.; Yoshikawa, T.; Ikenaga, M.; Fouad El Basuini, M.; Mae S Santander-De Leon, S.; Maeda, H. Use of Purple Non-Sulfur Photosynthetic Bacteria (Rhodobacter sphaeroides) in Promoting Ciliated Protozoa Growth. Biocontrol Sci. 2020, 25, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Vadstein, O.; Bergh, Ø.; Gatesoupe, F.-J.; Galindo-Villegas, J.; Mulero, V.; Picchietti, S.; Scapigliati, G.; Makridis, P.; Olsen, Y.; Dierckens, K.; et al. Microbiology and immunology of fish larvae. Rev. Aquac. 2013, 5, S1–S25. [Google Scholar] [CrossRef]
- McCasker, N.; Humphries, P.; Meredith, S.; Klomp, N. Contrasting Patterns of Larval Mortality in Two Sympatric Riverine Fish Species: A Test of the Critical Period Hypothesis. PLoS ONE 2014, 9, e109317. [Google Scholar]
- Bricknell, I.; Dalmo, R.A. The use of immunostimulants in fish larval aquaculture. Fish Shellfish Immunol. 2005, 19, 457–472. [Google Scholar] [CrossRef]
- Zhang, Z.; Chi, H.; Dalmo, R.A. Trained Innate Immunity of Fish Is a Viable Approach in Larval Aquaculture. Front. Immunol. 2019, 10, 42. [Google Scholar] [CrossRef]
- Oyarbide, U.; Rainieri, S.; Pardo, M.A. Zebrafish (Danio rerio) Larvae as a System to Test the Efficacy of Polysaccharides as Immunostimulants. Zebrafish 2012, 9, 74–84. [Google Scholar] [CrossRef]
- Schar, D.; Zhao, C.; Wang, Y.; Larsson, D.G.J.; Gilbert, M.; Van Boeckel, T.P. Twenty-year trends in antimicrobial resistance from aquaculture and fisheries in Asia. Nat. Commun. 2021, 12, 5384. [Google Scholar] [CrossRef]
- Gewaily, M.S.; Abdo, S.E.; Moustafa, E.M.; AbdEl-kader, M.F.; Abd El-Razek, I.M.; El-Sharnouby, M.; Alkafafy, M.; Raza, S.H.; El Basuini, M.F.; Van Doan, H.; et al. Dietary Synbiotics Can Help Relieve the Impacts of Deltamethrin Toxicity of Nile Tilapia Reared at Low Temperatures. Animals 2021, 11, 1790. [Google Scholar] [CrossRef]
- Dossou, S.; Dawood, M.A.O.; Zaineldin, A.I.; Abouelsaad, I.A.; Mzengereza, K.; Shadrack, R.S.; Zhang, Y.; El-Sharnouby, M.; Ahmed, H.A.; El Basuini, M.F. Dynamical Hybrid System for Optimizing and Controlling Efficacy of Plant-Based Protein in Aquafeeds. Complexity 2021, 2021, 9957723. [Google Scholar] [CrossRef]
- Paray, B.A.; El-Basuini, M.F.; Alagawany, M.; Albeshr, M.F.; Farah, M.A.; Dawood, M.A.O. Yucca schidigera Usage for Healthy Aquatic Animals: Potential Roles for Sustainability. Animals 2021, 11, 93. [Google Scholar] [CrossRef]
- Shukla, S.; Dubey, K.K. CoQ10 a super-vitamin: Review on application and biosynthesis. 3 Biotech 2018, 8, 249. [Google Scholar] [CrossRef]
- Klingen, A.R.; Palsdottir, H.; Hunte, C.; Ullmann, G.M. Redox-linked protonation state changes in cytochrome bc1 identified by Poisson–Boltzmann electrostatics calculations. Biochim. Biophys. Acta—Bioenerg. 2007, 1767, 204–221. [Google Scholar] [CrossRef]
- Lenaz, G.; Fato, R.; Formiggini, G.; Genova, M.L. The role of Coenzyme Q in mitochondrial electron transport. Mitochondrion 2007, 7, S8–S33. [Google Scholar] [CrossRef]
- Bentinger, M.; Brismar, K.; Dallner, G. The antioxidant role of coenzyme Q. Mitochondrion 2007, 7, S41–S50. [Google Scholar] [CrossRef]
- Linnane, A.W.; Kios, M.; Vitetta, L. Coenzyme Q10—Its role as a prooxidant in the formation of superoxide anion/hydrogen peroxide and the regulation of the metabolome. Mitochondrion 2007, 7, S51–S61. [Google Scholar] [CrossRef]
- Hernández-Camacho, J.D.; Bernier, M.; López-Lluch, G.; Navas, P. Coenzyme Q10 Supplementation in Aging and Disease. Front. Physiol. 2018, 9, 44. [Google Scholar] [CrossRef]
- El Basuini, M.F.; Teiba, I.I.; Zaki, M.A.A.; Alabssawy, A.N.; El-Hais, A.M.; Gabr, A.A.; Dawood, M.A.O.; Zaineldin, A.I.; Mzengereza, K.; Shadrack, R.S.; et al. Assessing the effectiveness of CoQ10 dietary supplementation on growth performance, digestive enzymes, blood health, immune response, and oxidative-related genes expression of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 98, 420–428. [Google Scholar] [CrossRef]
- El Basuini, M.F.; Shahin, S.A.; Teiba, I.I.; Zaki, M.A.A.; El-Hais, A.M.; Sewilam, H.; Almeer, R.; Abdelkhalek, N.; Dawood, M.A.O. The influence of dietary coenzyme Q10 and vitamin C on the growth rate, immunity, oxidative-related genes, and the resistance against Streptococcus agalactiae of Nile tilapia (Oreochromis niloticus). Aquaculture 2021, 531, 735862. [Google Scholar] [CrossRef]
- Desprez, D.; Briand, C.; Hoareau, M.C.; Mélard, C.; Bosc, P.; Baroiller, J.F. Study of sex ratio in progeny of a complex Oreochromis hybrid, the Florida red tilapia. Aquaculture 2006, 251, 231–237. [Google Scholar] [CrossRef]
- Muntaziana, M.P.A.; Rahim, A.A.; Harmin, S.A.; Amin, S.M.N. Effect of Broodfish Sex Ratio on Seed Production of Red Tilapia in Suspended Hapa. J. Fish. Aquat. Sci. 2011, 6, 862–866. [Google Scholar] [CrossRef]
- Pasha, R.; Moon, T.W. Coenzyme Q10 protects against statin-induced myotoxicity in zebrafish larvae (Danio rerio). Environ. Toxicol. Pharmacol. 2017, 52, 150–160. [Google Scholar] [CrossRef]
- Francis, S.; Delgoda, R.; Young, R. Effects of embryonic exposure to $α$-lipoic acid or ascorbic acid on hatching rate and development of zebrafish (Danio rerio). Aquac. Res. 2012, 43, 777–788. [Google Scholar] [CrossRef]
- Defoirdt, T.; Sorgeloos, P.; Bossier, P. Alternatives to antibiotics for the control of bacterial disease in aquaculture. Curr. Opin. Microbiol. 2011, 14, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.; Gloria, C.; Navarrete, P. Antibiotics in Aquaculture—Use, Abuse and Alternatives. In Health and Environment in Aquaculture; InTech: Vienna, Austria, 2012. [Google Scholar]
- Capkin, E.; Ozcelep, T.; Kayis, S.; Altinok, I. Antimicrobial agents, triclosan, chloroxylenol, methylisothiazolinone and borax, used in cleaning had genotoxic and histopathologic effects on rainbow trout. Chemosphere 2017, 182, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Dossou, S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Dawood, M.A.O.; El Basuini, M.F.; Olivier, A.; Zaineldin, A.I. Growth performance, blood health, antioxidant status and immune response in red sea bream (Pagrus major) fed Aspergillus oryzae fermented rapeseed meal (RM-Koji). Fish Shellfish Immunol. 2018, 75, 253–262. [Google Scholar] [CrossRef]
- Yoshida, Y.; Hayakawa, M.; Habuchi, Y.; Niki, E. Evaluation of the dietary effects of coenzyme Q in vivo by the oxidative stress marker, hydroxyoctadecadienoic acid and its stereoisomer ratio. Biochim. Biophys. Acta—Gen. Subj. 2006, 1760, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.; Buron, I.; Espinosa, J.; Castilla, J.; Villalba, J.; Torres, J. Coenzyme Q and protein/lipid oxidation in a BSE-infected transgenic mouse model. Free Radic. Biol. Med. 2007, 42, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Nepal, P.R.; Han, H.-K.; Choi, H.-K. Enhancement of solubility and dissolution of Coenzyme Q10 using solid dispersion formulation. Int. J. Pharm. 2010, 383, 147–153. [Google Scholar] [CrossRef]
- Falahatkar, B.; Dabrowski, K.; Arslan, M.; Rinchard, J. Effects of ascorbic acid enrichment by immersion of rainbow trout (Oncorhynchus mykiss, Walbaum 1792) eggs and embryos. Aquac. Res. 2006, 37, 834–841. [Google Scholar] [CrossRef]
- Henriksen, J.E.; Andersen, C.B.; Hother-Nielsen, O.; Vaag, A.; Mortensen, S.A.; Beck-Nielsen, H. Impact of ubiquinone (coenzyme Q10) treatment on glycaemic control, insulin requirement and well-being in patients with type I diabetes mellitus. Diabet. Med. 1999, 16, 312–318. [Google Scholar] [CrossRef]
- Schmelzer, C.; Lindner, I.; Rimbach, G.; Niklowitz, P.; Menke, T.; Döring, F. Functions of coenzyme Q10 in inflammation and gene expression. Biofactors 2008, 32, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Mariscal, F.M.; Yubero-Serrano, E.M.; Villalba, J.M.; Lopez-Miranda, J. Coenzyme Q 10: From bench to clinic in aging diseases, a translational review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2240–2257. [Google Scholar] [CrossRef] [PubMed]
- Hamre, K.; Krossøy, C.; Lock, E.-J.; Moren, M. Roles of lipid-soluble vitamins during ontogeny of marine fish larvae. Aquac. Res. 2010, 41, 745–750. [Google Scholar] [CrossRef]
- El Kertaoui, N.; Lund, I.; Assogba, H.; Domínguez, D.; Izquierdo, M.S.; Baekelandt, S.; Cornet, V.; Mandiki, S.N.M.; Montero, D.; Kestemont, P. Key nutritional factors and interactions during larval development of pikeperch (Sander lucioperca). Sci. Rep. 2019, 9, 7074. [Google Scholar] [CrossRef]
- Hamre, K. Metabolism, interactions, requirements and functions of vitamin E in fish. Aquac. Nutr. 2011, 17, 98–115. [Google Scholar] [CrossRef]
- El Basuini, M.F.; El-Hais, A.M.; Dawood, M.A.O.; Abou-Zeid, A.E.S.; EL-Damrawy, S.Z.; Khalafalla, M.M.E.S.; Koshio, S.; Ishikawa, M.; Dossou, S. Effect of different levels of dietary copper nanoparticles and copper sulfate on growth performance, blood biochemical profiles, antioxidant status and immune response of red sea bream (Pagrus major). Aquaculture 2016, 455, 32–40. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S. Vitamin C supplementation to optimize growth, health and stress resistance in aquatic animals. Rev. Aquac. 2018, 10, 334–350. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S. Effects of dietary inactivated Pediococcus pentosaceus on growth performance, feed utilization and blood characteristics of red sea bream, Pagrus major juvenile. Aquac. Nutr. 2016, 22, 923–932. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Magouz, F.I.; Salem, M.F.I.; Abdel-daim, H.A. Modulation of digestive enzyme activity, blood health, oxidative responses and growth-related gene expression in GIFT by heat-killed Lactobacillus plantarum ( L-137 ). Aquaculture 2019, 505, 127–136. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments. | Hatchability % | Yolk-Sac Diameter (mm) | |||
|---|---|---|---|---|---|
| 1 dph | 2 dph | ||||
| Depth (mm) | Width (mm) | Depth (mm) | Width (mm) | ||
| Control | 38.00 ± 15.88 | 1.73 ± 0.15 | 1.94 ± 0.14 a | 1.86 ± 0.08 | 1.88 ± 0.23 |
| C5 | 39.33 ± 6.11 | 1.72 ± 0.06 | 1.74 ± 0.04 b | 1.85 ± 0.13 | 1.79 ± 0.15 |
| C10 | 54.67 ± 11.55 | 1.63 ± 0.07 | 1.77 ± 0.09 ab | 1.87 ± 0.25 | 1.63 ± 0.07 |
| Treatments | SL (mm) | FTL (mm) | Final Weight 30 dph (g) | SR% | Deformed Fish% | |||
|---|---|---|---|---|---|---|---|---|
| 1 dph | 2 dph | 6 dph | 30 dph | |||||
| Control | 5.13 ± 0.15 a | 5.42 ± 0.56 | 7.22 ± 0.04 | 23.11 ± 1.84 ab | 30.00 ± 2.03 ab | 0.59 ± 0.06 a | 23.33 ± 7.02 | - |
| C5 | 4.96 ± 0.19 ab | 5.59 ± 0.24 | 7.03 ± 0.33 | 25.33 ± 1.33 a | 32.67 ± 1.33 a | 0.53 ± 0.06 ab | 22.67 ± 11.02 | - |
| C10 | 4.79 ± 0.02 b | 5.53 ± 0.09 | 7.39 ± 0.80 | 21.78 ± 1.35 b | 28.00 ± 0.88 b | 0.46 ± 0.02 b | 32.00 ± 2.00 | 2.08 ± 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mourad, M.M.; Shahin, S.A.; El-Ratel, I.T.; El Basuini, M.F. Effect of Treating Eggs with Coenzyme Q10 (CoQ10) on Growth Variables, Histomorphometry, and Antioxidant Capacity in Red Tilapia (Oreochromis aureus × Oreochromis mossambicus) Larvae. Animals 2022, 12, 2219. https://doi.org/10.3390/ani12172219
Mourad MM, Shahin SA, El-Ratel IT, El Basuini MF. Effect of Treating Eggs with Coenzyme Q10 (CoQ10) on Growth Variables, Histomorphometry, and Antioxidant Capacity in Red Tilapia (Oreochromis aureus × Oreochromis mossambicus) Larvae. Animals. 2022; 12(17):2219. https://doi.org/10.3390/ani12172219
Chicago/Turabian StyleMourad, Mona M., Shimaa A. Shahin, Ibrahim T. El-Ratel, and Mohammed F. El Basuini. 2022. "Effect of Treating Eggs with Coenzyme Q10 (CoQ10) on Growth Variables, Histomorphometry, and Antioxidant Capacity in Red Tilapia (Oreochromis aureus × Oreochromis mossambicus) Larvae" Animals 12, no. 17: 2219. https://doi.org/10.3390/ani12172219
APA StyleMourad, M. M., Shahin, S. A., El-Ratel, I. T., & El Basuini, M. F. (2022). Effect of Treating Eggs with Coenzyme Q10 (CoQ10) on Growth Variables, Histomorphometry, and Antioxidant Capacity in Red Tilapia (Oreochromis aureus × Oreochromis mossambicus) Larvae. Animals, 12(17), 2219. https://doi.org/10.3390/ani12172219