
What Happens in Male Dogs after Treatment with a 4.7 mg Deslorelin Implant? I. Flare up and Downregulation
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Physical and Andrological Examination including Evaluation of Implantation Site
2.3. Ultrasound of the Prostate Gland
2.4. Semen Collection and Analysis, Urine Examination
2.5. Blood Collection, GnRH/hCG Stimulation Tests and Hormone Analysis
2.6. Hemicastration and Implant Removal
2.7. Processing of Testicular Tissues and Further Evaluation
2.8. Statistical Analyses
3. Results
3.1. Physical Examinations and Evaluation of the Treatment Site
3.2. Andrological Examination
3.3. Ultrasound of the Prostate Gland
3.4. Semen Collection and Analysis
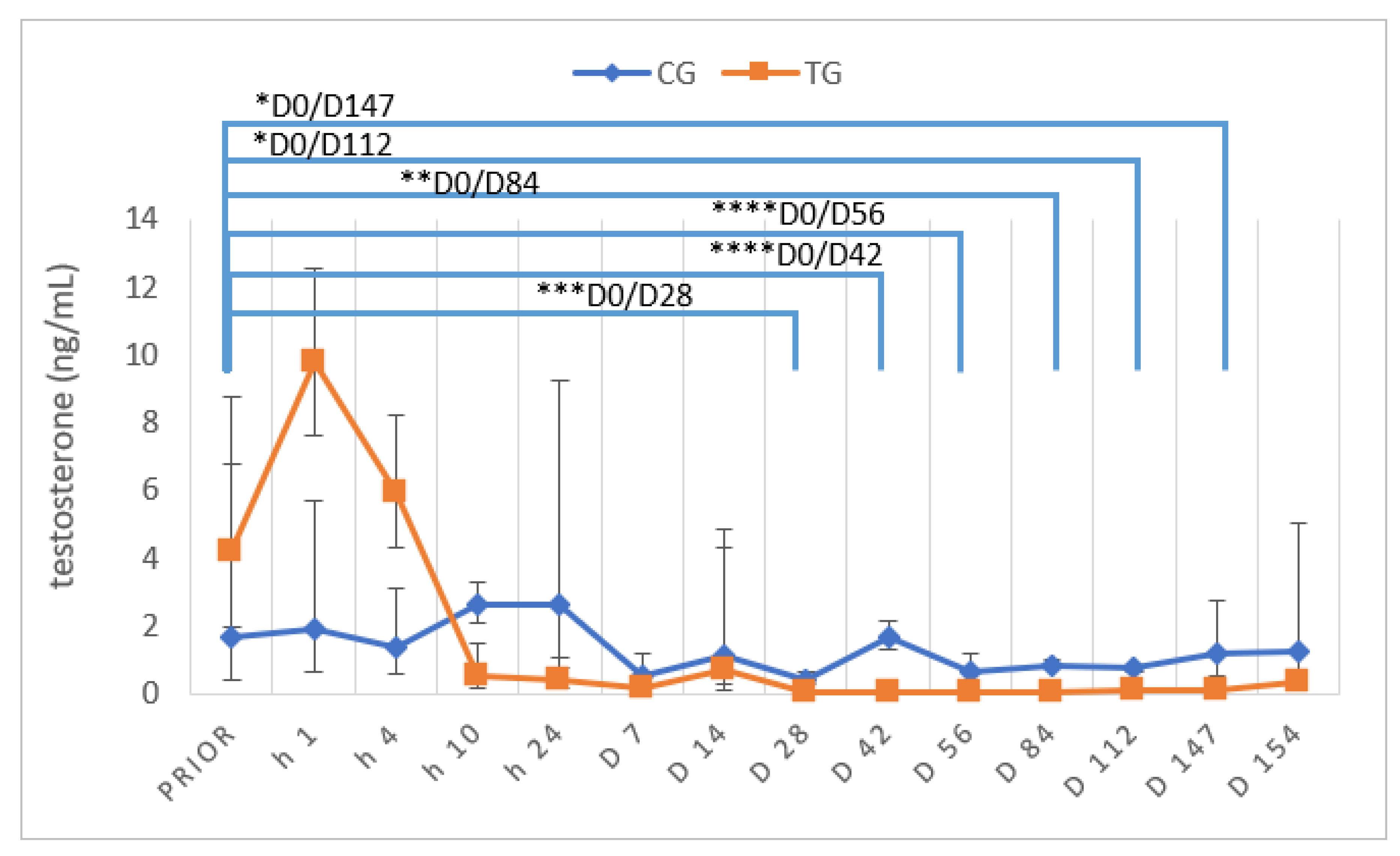
3.5. Testosterone Concentrations and Stimulation Tests
3.6. Histology of Testes
3.6.1. Vimentin Protein Expression
3.6.2. DAZL Protein Expression
4. Discussion
4.1. Evaluation of the Implantation Site and Implant Removal
4.2. Testicular and Prostatic Volume
4.3. Testosterone
4.4. Semen Evaluation
4.5. Histology and Immunhistochemistry
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arlt, S. Die hormonelle Kastration von Rüde und Hündin - ein Überblick. Veterinärspiegel 2010, 20, 155–160. [Google Scholar] [CrossRef]
- Goericke-Pesch, S. Alternativen zur chirurgischen Kastration bei Hunden, Katzen und kleinen Heimtieren. Kleintierprax 2016, 61, 657–665. [Google Scholar]
- Kutzler, M.; Wood, A. Non-surgical methods of contraception and sterilization. Theriogenology 2006, 66, 514–525. [Google Scholar] [CrossRef]
- Maenhoudt, C.; Santos, N.R.; Fontbonne, A. Suppression of fertility in adult dogs. Reprod. Domest. Anim. 2014, 49, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Palm, J.; Reichler, I.M. The use of deslorelin acetate (Suprelorin(R)) in companion animal medicine. Schweiz Arch. Tierheilkd 2012, 154, 7–12. [Google Scholar] [CrossRef]
- Riesenbeck, A.; Klein, R.; Hoffmann, B. Downregulation, eine neue, reversible Möglichkeit zur Ausschaltung der Hodenfunktion beim Rüden. Prakt. Tierarzt 2002, 83, 512–520. [Google Scholar]
- Romagnoli, S.; Siminica, A.; Sontas, B.H.; Milani, C.; Mollo, A.; Stelletta, C. Semen quality and onset of sterility following administration of a 4.7-mg deslorelin implant in adult male dogs. Reprod. Domest. Anim. 2012, 47, 389–392. [Google Scholar] [CrossRef]
- Trigg, T.E.; Wright, P.J.; Armour, A.F.; Williamson, P.E.; Junaidi, A.; Martin, G.B.; Doyle, A.G.; Walsh, J. Use of a GnRH analogue implant to produce reversible long-term suppression of reproductive function in male and female domestic dogs. J. Reprod. Fertil. Suppl. 2001, 57, 255–261. [Google Scholar]
- Stempel, S.; Goericke-Pesch, S. GnRH agonist implants in small animal practice - what do we know 13 years following EU registration? Tierarztl. Prax. Ausg. K Kleintiere Heimtiere 2020, 48, 420–432. [Google Scholar]
- Ludwig, C.; Desmoulins, P.O.; Driancourt, M.A.; Goericke-Pesch, S.; Hoffmann, B. Reversible downregulation of endocrine and germinative testicular function (hormonal castration) in the dog with the GnRH-agonist azagly-nafarelin as a removable implant “Gonazon”; a preclinical trial. Theriogenology 2009, 71, 1037–1045. [Google Scholar] [CrossRef]
- Animal Welfare Act. Available online: https://www.regjeringen.no/en/dokumenter/animal-welfare-act/id571188/ (accessed on 6 September 2022).
- Tierschutzgesetz. Available online: https://www.gesetze-im-internet.de/tierschg/BJNR012770972.html (accessed on 6 September 2022).
- Downes, M.J.; Devitt, C.; Downes, M.T.; More, S.J. Neutering of cats and dogs in Ireland; pet owner self-reported perceptions of enabling and disabling factors in the decision to neuter. PeerJ 2015, 3, e1196. [Google Scholar] [CrossRef] [PubMed]
- Root Kustritz, M.V. Effects of surgical sterilization on canine and feline health and on society. Reprod. Domest. Anim. 2012, 47, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Soto, F.R.; Ferreira, F.; Pinheiro, S.R.; Nogari, F.; Risseto, M.R.; de Souza, O.; Amaku, M. Adoption of shelter dogs in a Brazilian community: Assessing the caretaker profile. J. Appl. Anim. Welf. Sci. 2005, 8, 105–116. [Google Scholar] [CrossRef]
- Root Kustritz, M.V. Population Control in Small Animals. Vet. Clin. North Am. Small Anim. Pract. 2018, 48, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Ru, G.; Terracini, B.; Glickman, L.T. Host Related Risk Factors for Canine Osteosarcoma. Vet. J. 1998, 156, 31–39. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Goldschmidt, M.; Shofer, F.; Goldkamp, C.; Ferracone, J. Immunohistochemical characterization of canine prostatic carcinoma and correlation with castration status and castration time. Vet. Comp. Oncol. 2003, 1, 48–56. [Google Scholar] [CrossRef]
- van Hagen, M.A.; Ducro, B.J.; van den Broek, J.; Knol, B.W. Incidence, risk factors, and heritability estimates of hind limb lameness caused by hip dysplasia in a birth cohort of boxers. Am. J. Vet. Res. 2005, 66, 307–312. [Google Scholar] [CrossRef]
- Bryan, J.N.; Keeler, M.R.; Henry, C.J.; Bryan, M.E.; Hahn, A.W.; Caldwell, C.W. A population study of neutering status as a risk factor for canine prostate cancer. Prostate 2007, 67, 1174–1181. [Google Scholar] [CrossRef]
- Teske, E.; Naan, E.C.; van Dijk, E.M.; Van Garderen, E.; Schalken, J.A. Canine prostate carcinoma: Epidemiological evidence of an increased risk in castrated dogs. Mol. Cell. Endocrinol. 2002, 197, 251–255. [Google Scholar] [CrossRef]
- Junaidi, A.; Williamson, P.E.; Cummins, J.M.; Martin, G.B.; Blackberry, M.A.; Trigg, T.E. Use of a new drug delivery formulation of the gonadotrophin-releasing hormone analogue Deslorelin for reversible long-term contraception in male dogs. Reprod. Fertil. Dev. 2003, 15, 317–322. [Google Scholar] [CrossRef]
- Junaidi, A.; Williamson, P.E.; Martin, G.B.; Stanton, P.G.; Blackberry, M.A.; Cummins, J.M.; Trigg, T.E. Pituitary and testicular endocrine responses to exogenous gonadotrophin-releasing hormone (GnRH) and luteinising hormone in male dogs treated with GnRH agonist implants. Reprod. Fertil. Dev. 2007, 19, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Junaidi, A.; Williamson, P.E.; Trigg, T.E.; Cummins, J.M.; Martin, G.B. Morphological study of the effects of the GnRH superagonist deslorelin on the canine testis and prostate gland. Reprod. Domest. Anim. 2009, 44, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Goericke-Pesch, S.; Ludwig, C.; Hoffmann, B. Development of semen quality following reversible downregulation of testicular function in male dogs with a GnRH agonist implant. Reprod. Domest. Anim. 2012, 47, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Goericke-Pesch, S.; Spang, A.; Schulz, M.; Ozalp, G.; Bergmann, M.; Ludwig, C.; Hoffmann, B. Recrudescence of spermatogenesis in the dog following downregulation using a slow release GnRH agonist implant. Reprod. Domest. Anim. 2009, 44, 302–308. [Google Scholar] [CrossRef]
- Vickery, B.H.; McRae, G.I.; Goodpasture, J.C.; Sanders, L.M. Use of potent LHRH analogues for chronic contraception and pregnancy termination in dogs. J. Reprod. Fertil. Suppl. 1989, 39, 175–187. [Google Scholar]
- Vickery, B.H.; McRae, G.I.; Bonasch, H. Effect of chronic administration of a highly potent LHRH agonist on prostate size and secretory function in geriatric dogs. Prostate 1982, 3, 123–130. [Google Scholar] [CrossRef]
- Goericke-Pesch, S.; Wilhelm, E.; Hoffmann, B. Hormonelle Downregulation der Hodenfunktion bei Rüde und Kater; eine retrospektive Studie. Prakt. Tierarzt. 2010, 91, 563–570. [Google Scholar]
- Polisca, A.; Orlandi, R.; Troisi, A.; Brecchia, G.; Zerani, M.; Boiti, C.; Zelli, R. Clinical efficacy of the GnRH agonist (deslorelin) in dogs affected by benign prostatic hyperplasia and evaluation of prostatic blood flow by Doppler ultrasound. Reprod. Domest. Anim. 2013, 48, 673–680. [Google Scholar] [CrossRef]
- Junaidi, A.; Williamson, P.E.; Martin, G.B.; Blackberry, M.A.; Cummins, J.M.; Trigg, T.E. Dose-response studies for pituitary and testicular function in male dogs treated with the GnRH superagonist, deslorelin. Reprod. Domest. Anim. 2009, 44, 725–734. [Google Scholar] [CrossRef]
- Nizanski, W.; Ochota, M.; Fontaine, C.; Pasikowska, J. Comparison of Clinical Effectiveness of Deslorelin Acetate and Osaterone Acetate in Dogs with Benign Prostatic Hyperplasia. Animals 2020, 10, 1936. [Google Scholar] [CrossRef]
- Nizanski, W.; Ochota, M.; Fontaine, C.; Pasikowska, J. B-Mode and Doppler Ultrasonographic Findings of Prostate Gland and Testes in Dogs Receiving Deslorelin Acetate or Osaterone Acetate. Animals 2020, 10, 2379. [Google Scholar] [CrossRef] [PubMed]
- Gouletsou, P.G.; Galatos, A.D.; Leontides, L.S. Comparison between ultrasonographic and caliper measurements of testicular volume in the dog. Anim. Reprod. Sci. 2008, 108, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Günzel-Apel, A.R.; Bostedt, H. Reproduktionsmedizin und Neonatologie von Hund und Katze, 1st ed.; Schattauer GmbH: Stuttgart, Germany, 2016; pp. 3–738. [Google Scholar]
- Ruel, Y.; Barthez, P.Y.; Mailles, A.; Begon, D. Ultrasonographic evaluation of the prostate in healthy intact dogs. Vet. Radiol. Ultrasound 1998, 39, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Werhahn Beining, F.; Urhausen, C.; Wolf, K.; Schmicke, M.; Rohn, K.; Schuler, G.; Gunzel-Apel, A.R. Rhodesian Ridgebacks have an increased risk to develop benign prostatic hyperplasia. Reprod. Domest. Anim. 2020, 55, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Moxon, R.; Bright, L.; Pritchard, B.; Bowen, I.M.; de Souza, M.B.; da Silva, L.D.; England, G.C. Digital image analysis of testicular and prostatic ultrasonographic echogencity and heterogeneity in dogs and the relation to semen quality. Anim. Reprod. Sci. 2015, 160, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Mantziaras, G.; Alonge, S.; Faustini, M.; Luvoni, G.C. Assessment of the age for a preventive ultrasonographic examination of the prostate in the dog. Theriogenology 2017, 100, 114–119. [Google Scholar] [CrossRef]
- Pesch, S.; Schuler, G.; Wilhelm, E.; Hoffmann, B. Samengewinnung, -konservierung und künstliche Besamung beim Hund. Tierärztl. Praxis 2007, 35, 81–90. [Google Scholar]
- Goericke-Pesch, S.; Failing, K. Retrospective analysis of canine semen evaluations with special emphasis on the use of the hypoosmotic swelling (HOS) test and acrosomal evaluation using Spermac((R)). Reprod. Domest. Anim. 2013, 48, 213–217. [Google Scholar] [CrossRef]
- Günzel-Apel, A.R.; Terhaer, P.; Waberski, D. Hodendimensionen und Ejakulatbeschaffenheit fertiler Rüden unterschiedlicher Körpergewichte. Kleintierpraxis 1994, 39, 483–486. [Google Scholar]
- Hoffmann, B.; Landeck, A. Testicular endocrine function, seasonality and semen quality of the stallion. Animal Reproduction Science 1999, 57, 89–98. [Google Scholar] [CrossRef]
- Röcken, F.E.; Nothelfer, H.B.; Hoffmann, B. Testosteronkonzentrationen im peripheren Plasma sowie morphologische Hodenbefunde von Rüden mit einer Perinealhernie. Kleintierpraxis 1995, 40, 261–267. [Google Scholar]
- Goericke-Pesch, S. Kryptorchismus bei Hund und Katze. Kleintierprax 2010, 55, 255–261. [Google Scholar]
- Körber, H.; Goericke-Pesch, S. Expression of PTGS2, PGFS and PTGFR during downregulation and restart of spermatogenesis following GnRH agonist treatment in the dog. Cell Tissue Res. 2019, 375, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Körber, H.; Goericke-Pesch, S. Expression of prostaglandin (PG) D synthase lipocalin and hematopoietic type and PG D receptor during restart of spermatogenesis following downregulation using a slow release GnRH agonist implant in the dog. Cell Tissue Res. 2019, 378, 359–370. [Google Scholar] [CrossRef]
- Goericke-Pesch, S.; Gentil, M.; Spang, A.; Kowalewski, M.P.; Failing, K.; Hoffmann, B. Status of the down-regulated canine testis using two different GNRH agonist implants in comparison with the juvenile testis. Reproduction 2013, 146, 517–526. [Google Scholar] [CrossRef]
- Ferre-Dolcet, L.; Carniello, L.; Ferro, S.; Cattai, A.; Romagnoli, S.; Mollo, A. Interval between Removal of a 4.7 mg Deslorelin Implant after a 3-, 6-, and 9-Month Treatment and Restoration of Testicular Function in Tomcats. Animals 2020, 10, 1559. [Google Scholar] [CrossRef]
- Goericke-Pesch, S.; Georgiev, P.; Antonov, A.; Albouy, M.; Wehrend, A. Clinical efficacy of a GnRH-agonist implant containing 4.7 mg deslorelin, Suprelorin, regarding suppression of reproductive function in tomcats. Theriogenology 2011, 75, 803–810. [Google Scholar] [CrossRef]
- Goericke-Pesch, S.; Georgiev, P.; Fasulkov, I.; Vodenicharov, A.; Wehrend, A. Basal testosterone concentrations after the application of a slow-release GnRH agonist implant are associated with a loss of response to buserelin, a short-term GnRH agonist, in the tom cat. Theriogenology 2013, 80, 65–69. [Google Scholar] [CrossRef]
- Novotny, R.; Vitasek, R.; Bartoskova, A.; Cizek, P.; Prinosilova, P.; Novakova, K. Azoospermia with variable testicular histology after 7 months of treatment with a deslorelin implant in toms. Theriogenology 2015, 83, 1188–1193. [Google Scholar] [CrossRef]
- De Palatis, L.; Moore, J.; Falvo, R.E. Plasma concentrations of testosterone and LH in the male dog. J. Reprod. Fertil. 1978, 52, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Goericke-Pesch, S.; Wilhelm, E.; Ludwig, C.; Desmoulins, P.O.; Driancourt, M.A.; Hoffmann, B. Evaluation of the clinical efficacy of Gonazon implants in the treatment of reproductive pathologies, behavioral problems, and suppression of reproductive function in the male dog. Theriogenology 2010, 73, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Trigg, T.E.; Doyle, A.G.; Walsh, J.D.; Swangchan-uthai, T. A review of advances in the use of the GnRH agonist deslorelin in control of reproduction. Theriogenology 2006, 66, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
- Balogh, O.; Somoskoi, B.; Kollar, E.; Kowalewski, M.P.; Gram, A.; Reichler, I.M.; Klein, R.; Kawate, N.; Mester, L.; Walter, B.; et al. Anti-Mullerian hormone, testosterone, and insulin-like peptide 3 as biomarkers of Sertoli and Leydig cell function during deslorelin-induced testicular downregulation in the dog. Theriogenology 2021, 175, 100–110. [Google Scholar] [CrossRef]
- Günzel-Apel, A.R.; Brinckmann, H.G.; Hoppen, H.O. Dynamik der LH- und Testosteron-Sekretion bei Beagle-Rüden verschiedener Altersgruppen. Reprod. Domest. Anim. 1990, 25, 78–86. [Google Scholar] [CrossRef]
- Sundby, A.; Ulstein, T. Plasma Concentrations of Testosterone in the Male Dog and Plasma Testosterone Profile Following Single Intramuscular Injection of HCG. Acta. Vet. Scand. 1981, 22, 409–416. [Google Scholar] [CrossRef]
- Taha, M.B.; Noakes, D.E. The effect of age and season of the year on testicular function in the dog, as determined by histological examination of the seminiferous tubules and the estimation of peripheral plasma testosterone concentrations. J. small Anim. Pract. 1982, 23, 352–357. [Google Scholar] [CrossRef]
- Rispoli, L.A.; Nett, T.M. Pituitary gonadotropin-releasing hormone (GnRH) receptor: Structure, distribution and regulation of expression. Anim. Reprod. Sci. 2005, 88, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Hazum, E.; Cuatrecasas, P.; Marian, J.; Conn, P.M. Receptor-mediated internalization of fluorescent gonadotropin-releasing hormone by pituitary gonadotropes. Proc. Natl. Acad. Sci. USA 1980, 77, 6692–6695. [Google Scholar] [CrossRef]
- Gentil, M.; Hoffmann, B.; Spang, A.; Failing, K.; Goericke-Pesch, S. Restart of steroidogenesis in dogs during recrudescence of testicular function following downregulation with a GnRH-agonist implant. Cell Tissue Res. 2012, 350, 513–523. [Google Scholar] [CrossRef]
- Novotny, R.; Cizek, P.; Vitasek, R.; Bartoskova, A.; Prinosilova, P.; Janosovska, M. Reversible suppression of sexual activity in tomcats with deslorelin implant. Theriogenology 2012, 78, 848–857. [Google Scholar] [CrossRef]
- Gregory, S.J.; Kaiser, U.B. Regulation of gonadotropins by inhibin and activin. Semin. Reprod. Med. 2004, 22, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.J.; Parker, W.G.; Larson, L.L. Duration of Estrus and Ovulation Time in Nonlactating Mares Given Human Chorionic Gonadotropin During Three Successive Estrous Periods. J. Am. Vet. Med. Assoc. 1973, 162, 895–898. [Google Scholar] [PubMed]
- Wilson, C.G.; Downie, C.R.; Hughes, J.P.; Roser, J.F. Effects of repeated hCG injections on reproductive efficiency in mares. J. Equine Vet. Sci. 1990, 10, 301–308. [Google Scholar] [CrossRef]
- Giordano, J.O.; Wiltbank, M.C.; Fricke, P.M. Humoral immune response in lactating dairy cows after repeated exposure to human chorionic gonadotropin. Theriogenology 2012, 78, 218–224. [Google Scholar] [CrossRef]
- Sundby, A.; Torjesen, P.A. Plasma levels of testosterone in bulls. Response to repeated HCG injections. Acta. Endocrinol. 1978, 88, 787–792. [Google Scholar] [CrossRef]
- Swanson, W.F.; Roth, T.L.; Graham, K.; Horohov, D.W.; Godke, R.A. Kinetics of the humoral immune response to multiple treatments with exogenous gonadotropins and relation to ovarian responsiveness in domestic cats. Am. J. Vet. Res. 1996, 57, 302–307. [Google Scholar]
- Swanson, W.F.; Horohov, D.W.; Godke, R.A. Production of exogenous gonadotrophin-neutralizing immunoglobulins in cats after repeated eCG-hCG treatment and relevance for assisted reproduction in felids. J. Reprod. Fertil. 1995, 105, 35–41. [Google Scholar] [CrossRef]
- Al-Kafawi, A.A.; Hopwood, M.L.; Pineda, M.H.; Faulkner, L.C. Immunization of dogs against human chorionic gonadotropin. Am. J. Vet. Res. 1974, 35, 261–264. [Google Scholar]
- Beach, F.A. Coital Behavior in Dogs. VI. Long-term Effects of Castration Upon Mating in the Male. J. Comp. Physiol. Psychol. 1970, 70, 1–32. [Google Scholar] [CrossRef]
- Pieri, N.; Souza, A.F.; Mancanares, A.; Roballo, K.; Casals, J.B.; Ambrosio, C.E.; Martins, D.S. Immunolocalization of proteins in the spermatogenesis process of canine. Reprod. Domest. Anim. 2017, 52, 170–176. [Google Scholar] [CrossRef] [Green Version]











 spermatogonia;
spermatogonia;  spermatocytes;
spermatocytes;  round spermatids;
round spermatids;  elongated spermatids;
elongated spermatids;  Sertoli cells; white arrow, Leydig cells.
spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells.
Sertoli cells; white arrow, Leydig cells.
spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells. spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells.
spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells.
spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells.
spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells. spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells.
spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells.
spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells.
spermatogonia; spermatocytes; round spermatids; elongated spermatids; Sertoli cells; white arrow, Leydig cells.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Testosterone (ng/mL) | Most Developed Germ Cell: | Level of Arrest of Spermatogenesis (Most Developed) | Semen Findings |
|---|---|---|---|---|
| Control group: | ||||
| Piet Knopf Merlin | 0.25 1.91 3.78 | Elongated spermatids | Full/Normal spermatogenesis | Normospermia |
| Treated group: | ||||
| Ole | 0.10 | Spermatogonia | Arrest at level of spermatogonia | Aspermia (no ejaculate) |
| Strolch | 0.10 | Spermatogonia | ||
| Fiete | 0.16 | Spermatogonia | ||
| Spencer | 0.13 | Spermatogonia | ||
| Murphy | 0.27 | Spermatogonia | ||
| Toni | 1.76 | Spermatocytes | Arrest at level of spermatocytes | Azoospermia (VOL: 0.1 mL) |
| Lui | 3.16 | Elongating spermatids | Full spermatogenesis | Pathospermia (VOL: 0.1 mL PM: 20% MAS: 97% *) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stempel, S.; Körber, H.; Reifarth, L.; Schuler, G.; Goericke-Pesch, S. What Happens in Male Dogs after Treatment with a 4.7 mg Deslorelin Implant? I. Flare up and Downregulation. Animals 2022, 12, 2379. https://doi.org/10.3390/ani12182379
Stempel S, Körber H, Reifarth L, Schuler G, Goericke-Pesch S. What Happens in Male Dogs after Treatment with a 4.7 mg Deslorelin Implant? I. Flare up and Downregulation. Animals. 2022; 12(18):2379. https://doi.org/10.3390/ani12182379
Chicago/Turabian StyleStempel, Sabrina, Hanna Körber, Larena Reifarth, Gerhard Schuler, and Sandra Goericke-Pesch. 2022. "What Happens in Male Dogs after Treatment with a 4.7 mg Deslorelin Implant? I. Flare up and Downregulation" Animals 12, no. 18: 2379. https://doi.org/10.3390/ani12182379
APA StyleStempel, S., Körber, H., Reifarth, L., Schuler, G., & Goericke-Pesch, S. (2022). What Happens in Male Dogs after Treatment with a 4.7 mg Deslorelin Implant? I. Flare up and Downregulation. Animals, 12(18), 2379. https://doi.org/10.3390/ani12182379