
Effects of Stocking Rate and Environmental Enrichment on the Ontogeny of Pecking Behavior of Laying Hen Pullets Confined in Aviary Compartments during the First 4 Weeks of Life



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Farm, Animals, and Management
2.2. Study Design
2.3. Video Observation
2.4. Statistics
3. Results
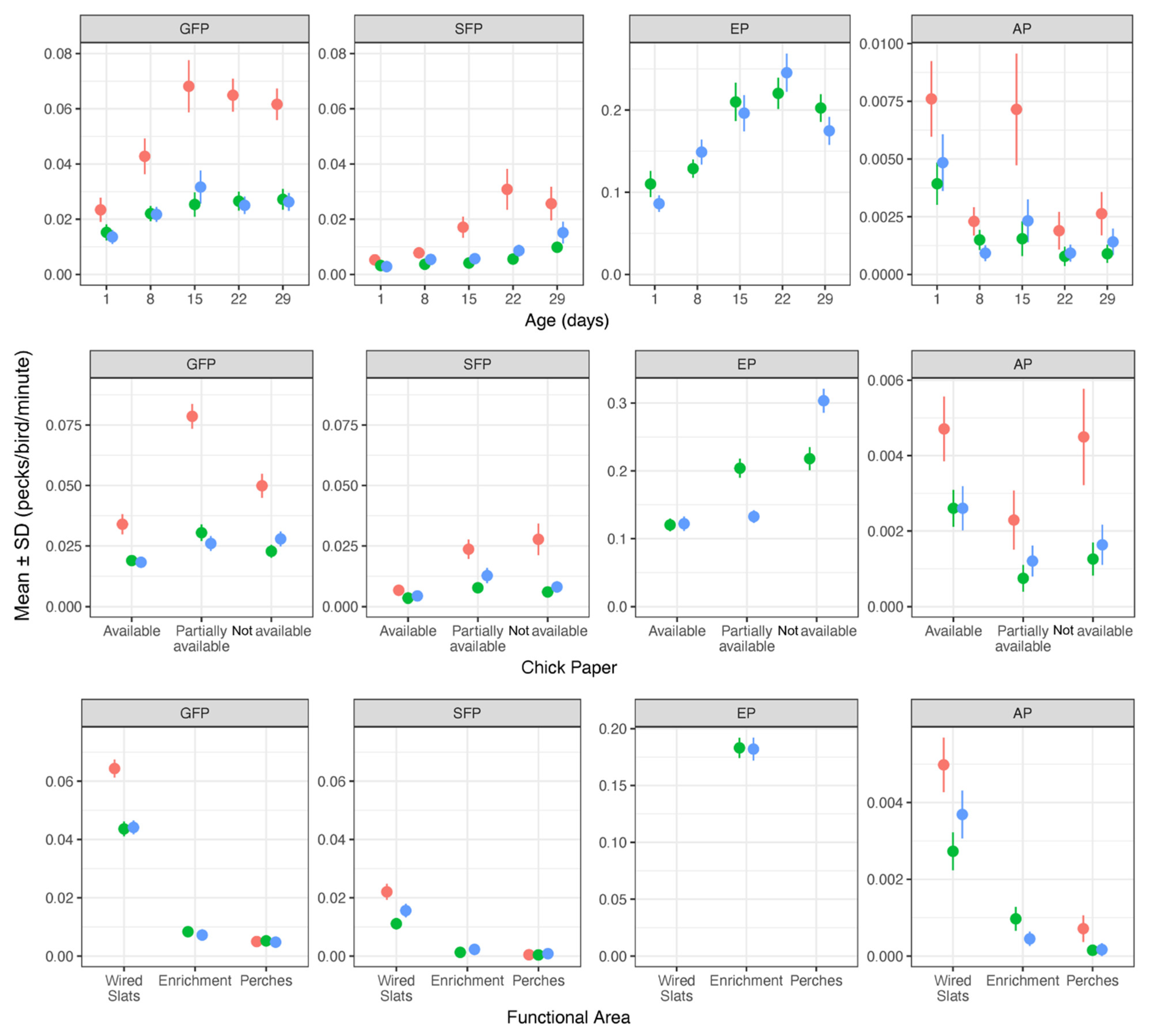
3.1. Ontogeny of the Pecking Behavior
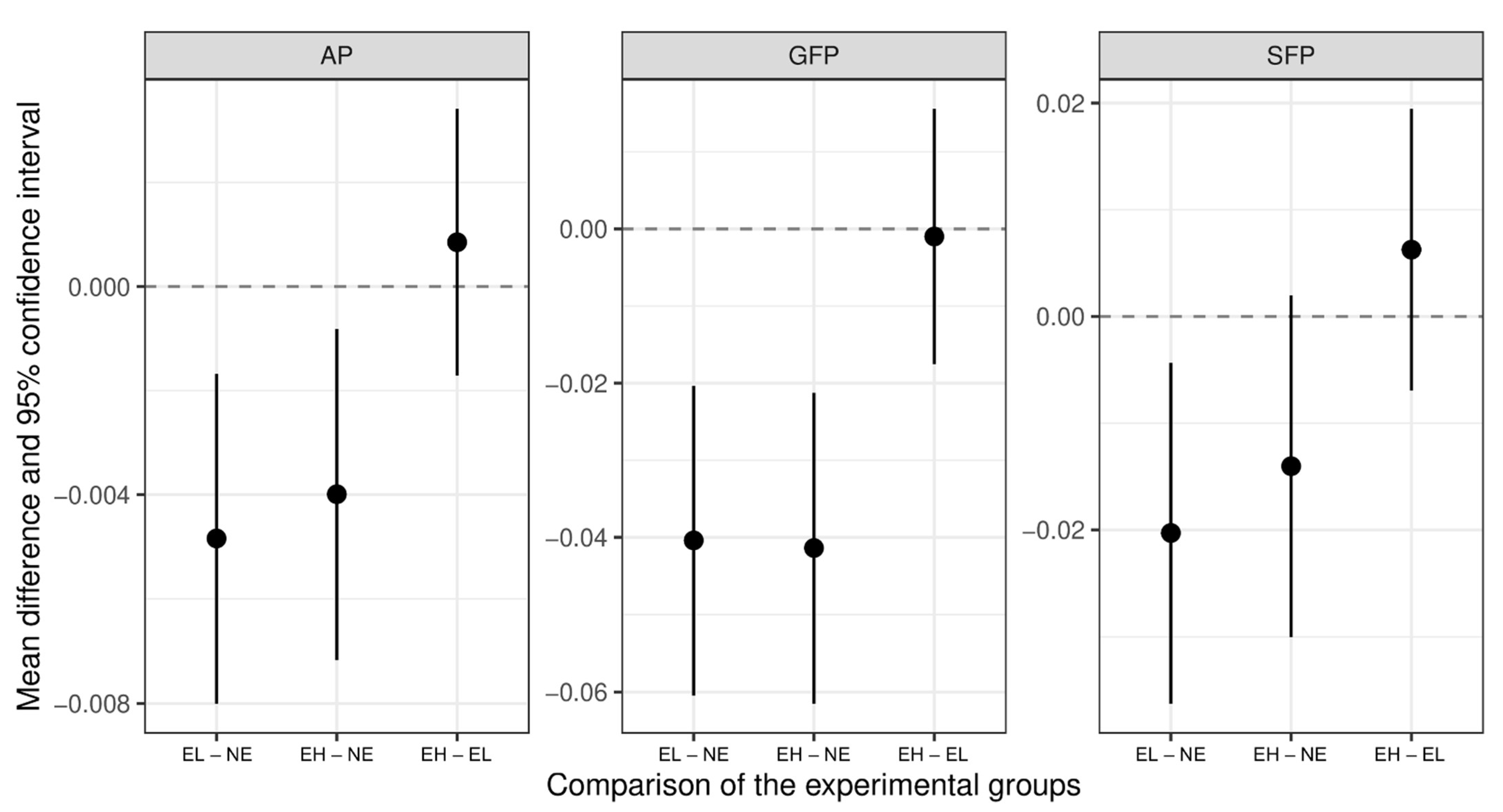
3.2. Effect of the Experimental Group on Pecking Behavior
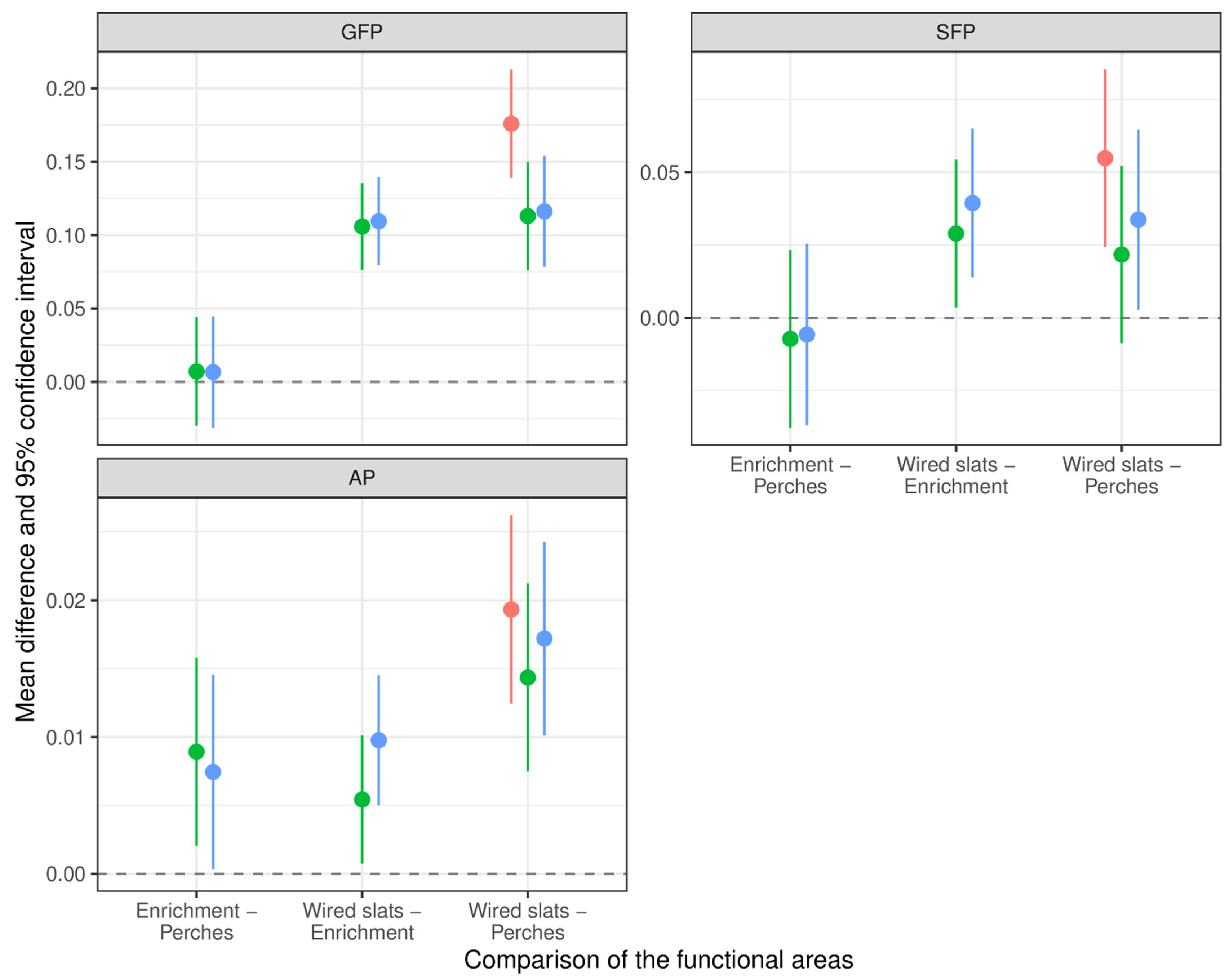
3.3. Pecking Behavior in Different Functional Areas
3.4. The Influence of Chick Paper on Pecking Behavior
3.5. Consumption of Enrichment Materials
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Hierden, Y.M.; de Boer, S.F.; Koolhaas, J.M.; Korte, S.M. The control of feather pecking by serotonin. Behav. Neurosci. 2004, 118, 575–583. [Google Scholar] [CrossRef]
- Gentle, M.J.; Hunter, L.N. Physiological and behavioural responses associated with feather removal in Gallus gallus var domesticus. Res. Vet. Sci. 1991, 50, 95–101. [Google Scholar] [CrossRef]
- Damme, K. Effect of beak-trimming and strain on performance, feather loss and nesting behaviour of different commercial white layer hybrids in floor pens. Arch. Geflügelkd. 1999, 63, 93–99. [Google Scholar]
- Savory, C.J. Feather pecking and cannibalism. World’s Poult. Sci. J. 1995, 51, 215–219. [Google Scholar] [CrossRef]
- Cloutier, S.; Newberry, R.C. A note on aggression and cannibalism in laying hens following re-housing and re-grouping. Appl. Anim. Behav. Sci. 2002, 76, 157–163. [Google Scholar] [CrossRef]
- Leonard, M.L.; Horn, A.G.; Fairfull, R. Correlates and consequences of allopecking in White Leghorn chickens. Appl. Anim. Behav. Sci. 1995, 43, 17–26. [Google Scholar] [CrossRef]
- Odén, K.; Vestergaard, K.; Algers, B. Agonistic behaviour and feather pecking in single-sexed and mixed groups of laying hens. Appl. Anim. Behav. Sci. 1999, 62, 219–231. [Google Scholar] [CrossRef]
- Iffland, H.; Schmid, M.; Preuß, S.; Bessei, W.; Tetens, J.; Bennewitz, J. Phenotypic and genomic analyses of agonistic interactions in laying hen lines divergently selected for feather pecking. Appl. Anim. Behav. Sci. 2021, 234, 105177. [Google Scholar] [CrossRef]
- Kruijt, J.P. Ontogeny of social behaviour in Burmese red junglefowl (Gallus gallus spadiceus). Behav. Suppl. 1964, 12, I–IX, 1–201. [Google Scholar]
- Riedstra, B.; Groothuis, T.G. Early feather pecking as a form of social exploration: The effect of group stability on feather pecking and tonic immobility in domestic chicks. Appl. Anim. Behav. Sci. 2002, 77, 127–138. [Google Scholar] [CrossRef]
- Huber-Eicher, B.; Wechsler, B. Feather pecking in domestic chicks: Its relation to dustbathing and foraging. Anim. Behav. 1997, 54, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, T.; van Hierden, Y.; Buitenhuis, A.; Riedstra, B.; Koene, P.; Korte, S.; van der Poel, J.; Groothuis, T.; Blokhuis, H. Feather pecking in laying hens: New insights and directions for research? Appl. Anim. Behav. Sci. 2004, 86, 291–298. [Google Scholar] [CrossRef]
- McAdie, T.; Keeling, L. The social transmission of feather pecking in laying hens: Effects of environment and age. Appl. Anim. Behav. Sci. 2002, 75, 147–159. [Google Scholar] [CrossRef]
- Guhl, A.M. Social behavior of the domestic fowl. Trans. Kans. Acad. Sci. 1968, 71, 379–384. [Google Scholar] [CrossRef] [PubMed]
- D’Eath, R.B.; Keeling, L.J. Social discrimination and aggression by laying hens in large groups: From peck orders to social tolerance. Appl. Anim. Behav. Sci. 2003, 84, 197–212. [Google Scholar] [CrossRef]
- McBRIDE, G.; FOENANDER, F. Territorial Behaviour in Flocks of Domestic Fowls. Nature 1962, 194, 102. [Google Scholar] [CrossRef]
- Pagel, M.; Dawkins, M. Peck orders and group size in laying hens: `futures contracts’ for non-aggression. Behav. Process. 1997, 40, 13–25. [Google Scholar] [CrossRef]
- Estevez, I.; Keeling, L.J.; Newberry, R.C. Decreasing aggression with increasing group size in young domestic fowl. Appl. Anim. Behav. Sci. 2003, 84, 213–218. [Google Scholar] [CrossRef]
- Ahlers, C.; Ahlers, N.; Böhmfeld, J.; Damme, K.; Gaio, C.; Hiller, P.; Kästner, B.; Keppler, C.; Menning, J.; Nette, A.; et al. Gesamtbetriebliches Haltungskonzept Junghennen. 2021. Available online: https://www.ble-medienservice.de/0341/gesamtbetriebliches-haltungskonzept-gefluegel-junghennen (accessed on 29 March 2022).
- Lohmann Breeders. Management Guide Alternative Systems: Management Recommendations for Barn, Aviary & Free Range Systems, Cuxhaven, 2021. Available online: https://lohmann-breeders.com/management-guide/alternative-housing-download/ (accessed on 13 April 2022).
- Gilani, A.-M.; Knowles, T.G.; Nicol, C.J. The effect of rearing environment on feather pecking in young and adult laying hens. Appl. Anim. Behav. Sci. 2013, 148, 54–63. [Google Scholar] [CrossRef]
- Johnsen, P.F.; Vestergaard, K.S.; Nørgaard-Nielsen, G. Influence of early rearing conditions on the development of feather pecking and cannibalism in domestic fowl. Appl. Anim. Behav. Sci. 1998, 60, 25–41. [Google Scholar] [CrossRef]
- Bestman, M.; Koene, P.; Wagenaar, J.-P. Influence of farm factors on the occurrence of feather pecking in organic reared hens and their predictability for feather pecking in the laying period. Appl. Anim. Behav. Sci. 2009, 121, 120–125. [Google Scholar] [CrossRef]
- Lambton, S.L.; Knowles, T.G.; Yorke, C.; Nicol, C.J. The risk factors affecting the development of gentle and severe feather pecking in loose housed laying hens. Appl. Anim. Behav. Sci. 2010, 123, 32–42. [Google Scholar] [CrossRef]
- Kaukonen, E.; Valros, A. Feather Pecking and Cannibalism in Non-Beak-Trimmed Laying Hen Flocks-Farmers’ Perspectives. Animals 2019, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Jongman, E.C. Rearing conditions of laying hens and welfare during the laying phase. Anim. Prod. Sci. 2021, 61, 876. [Google Scholar] [CrossRef]
- Hedlund, L.; Jensen, P. Effects of stress during commercial hatching on growth, egg production and feather pecking in laying hens. PLoS ONE 2022, 17, e0262307. [Google Scholar] [CrossRef]
- Blokhuis, H.J.; van der Haar, J.W. Effects of floor type during rearing and of beak trimming on ground pecking and feather pecking in laying hens. Appl. Anim. Behav. Sci. 1989, 22, 359–369. [Google Scholar] [CrossRef]
- Mels, C.; Niebuhr, K.; Futschik, A.; Rault, J.-L.; Waiblinger, S. Predictors for plumage damage and bloody lesions indicative of feather pecking in pullets reared in aviaries. Appl. Anim. Behav. Sci. 2022, 250, 105607. [Google Scholar] [CrossRef]
- Nicol, C.J.; Bestman, M.; Gilani, A.-M.; de Haas, E.N.; de Jong, I.C.; Lambton, S.; Wagenaar, J.P.; Weeks, C.A.; Rodenburg, T.B. The prevention and control of feather pecking: Application to commercial systems. World’s Poult. Sci. J. 2013, 69, 775–788. [Google Scholar] [CrossRef]
- Hansen, I. Behavioural expression of laying hens in aviaries and cages: Frequencies, time budgets and facility utilisation. Br. Poult. Sci. 1994, 35, 491–508. [Google Scholar] [CrossRef]
- Huber-Eicher, B.; Audigé, L. Analysis of risk factors for the occurrence of feather pecking in laying hen growers. Br. Poult. Sci. 1999, 40, 599–604. [Google Scholar] [CrossRef]
- Nicol, C.; Gregory, N.; Knowles, T.; Parkman, I.; Wilkins, L. Differential effects of increased stocking density, mediated by increased flock size, on feather pecking and aggression in laying hens. Appl. Anim. Behav. Sci. 1999, 65, 137–152. [Google Scholar] [CrossRef]
- Savory, C.J.; Mann, J.S. Feather pecking in groups of growing bantams in relation to floor litter substrate and plumage colour. Br. Poult. Sci. 1999, 40, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Zepp, M.; Louton, H.; Erhard, M.; Schmidt, P.; Helmer, F.; Schwarzer, A. The influence of stocking density and enrichment on the occurrence of feather pecking and aggressive pecking behavior in laying hen chicks. J. Vet. Behav. 2018, 24, 9–18. [Google Scholar] [CrossRef]
- Staack, M.; Gruber, B.; Keppler, C.; Zaludik, K.; Niebuhr, K.; Knierim, U. Bedeutung der Aufzucht der Legehennen für alternative Haltungsformen. Dtsch. Tierärztl. Wschr. 2007, 114, 86–90. [Google Scholar]
- Gunnarsson, S.; Keeling, L.J.; Svedberg, J. Effect of rearing factors on the prevalence of floor eggs, cloacal cannibalism and feather pecking in commercial flocks of loose housed laying hens. Br. Poult. Sci. 1999, 40, 12–18. [Google Scholar] [CrossRef]
- Keppler, C. Investigation of Important Influencing Factors Regarding Feather Pecking and Cannibalism in Non-Beak Trimmed Laying Hens Kept in Barns or Aviaries with Daylight and with Particular Consideration of the Rearing Period. Ph.D. Thesis, Kassel University, Kassel, Germany, 2008. [Google Scholar]
- Von Eugen, K.; Nordquist, R.E.; Zeinstra, E.; van der Staay, F.J. Stocking Density Affects Stress and Anxious Behavior in the Laying Hen Chick During Rearing. Animals 2019, 9, 53. [Google Scholar] [CrossRef]
- Gilani, A.-M.; Knowles, T.G.; Nicol, C.J. Factors affecting ranging behaviour in young and adult laying hens. Br. Poult. Sci. 2014, 55, 127–135. [Google Scholar] [CrossRef]
- Bilcík, B.; Keeling, L.J. Changes in feather condition in relation to feather pecking and aggressive behaviour in laying hens. Br. Poult. Sci. 1999, 40, 444–451. [Google Scholar] [CrossRef]
- Schwarzer, A.; Plattner, C.; Bergmann, S.; Rauch, E.; Erhard, M.; Reese, S.; Louton, H. Feather Pecking in Non-Beak-Trimmed and Beak-Trimmed Laying Hens on Commercial Farms with Aviaries. Animals 2021, 11, 3085. [Google Scholar] [CrossRef]
- Elger, M. Behaviour of Non-Debeaked Laying Hens on Farms with Particular Focus on Aggressive Pecking and Severe Feather Pecking in Different Functional Areas. Ph.D. Thesis, LMU Munich, Munich, Germany, 2017. [Google Scholar]
- Tahamtani, F.M.; Brantsæter, M.; Nordgreen, J.; Sandberg, E.; Hansen, T.B.; Nødtvedt, A.; Rodenburg, T.B.; Moe, R.O.; Janczak, A.M. Effects of litter provision during early rearing and environmental enrichment during the production phase on feather pecking and feather damage in laying hens. Poult. Sci. 2016, 95, 2747–2756. [Google Scholar] [CrossRef]
- Van Staaveren, N.; Ellis, J.; Baes, C.F.; Harlander-Matauschek, A. A meta-analysis on the effect of environmental enrichment on feather pecking and feather damage in laying hens. Poult. Sci. 2021, 100, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.E.; Nicol, C.J.; Weeks, C.A. The Effect of Hard Pecking Enrichment during Rear on Feather Cover, Feather Pecking Behaviour and Beak Length in Beak-Trimmed and Intact-Beak Laying Hen Pullets. Animals 2022, 12, 674. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.L.M.; Gerber, P.F.; Downing, J.A.; Lee, C. Minimal Effects of Rearing Enrichments on Pullet Behaviour and Welfare. Animals 2020, 10, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebers, C.J.; Schwarzer, A.; Erhard, M.; Schmidt, P.; Louton, H. The influence of environmental enrichment and stocking density on the plumage and health conditions of laying hen pullets. Poult. Sci. 2019, 98, 2474–2488. [Google Scholar] [CrossRef]
- Martin, P.; Bateson, P.P.G. Measuring Behaviour: An Introductory Guide, 3rd ed.; Cambridge University Press: Cambridge, UK, 2018; ISBN 9780511810893. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Krause, E.T.; Schrader, L. Suggestions to Derive Maximum Stocking Densities for Layer Pullets. Animals 2019, 9, 348. [Google Scholar] [CrossRef]
- Hansen, I.; Braastad, B.O. Effect of rearing density on pecking behaviour and plumage condition of laying hens in two types of aviary. Appl. Anim. Behav. Sci. 1994, 40, 263–272. [Google Scholar] [CrossRef]
- Huber-Eicher, B.; Wechsler, B. The effect of quality and availability of foraging materials on feather pecking in laying hen chicks. Anim. Behav. 1998, 55, 861–873. [Google Scholar] [CrossRef]
- McAdie, T.M.; Keeling, L.J.; Blokhuis, H.J.; Jones, R.B. Reduction in feather pecking and improvement of feather condition with the presentation of a string device to chickens. Appl. Anim. Behav. Sci. 2005, 93, 67–80. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| NE | EL | EH | |
|---|---|---|---|
| Group size until 10th day of life (no of pullets) | 230 | 203 | 230 |
| Group size from 11th day of life (no of pullets) | 115 | 101/102 | 115 |
| Pullets/m2 usable area until 10th day of life | 120.8 | 106.6 | 120.8 |
| Pullets/m2 usable area from 11th day of life | 60.4 | 53.6 | 60.4 |
| Enrichment (pecking stones and blocks) | No | Yes | Yes |
| Behavior | Definition |
|---|---|
| Enrichment pecking (EP) | Pecks directed against pecking blocks or pecking stones |
| Gentle feather pecking (GFP) | Gentle pecking against the plumage of other birds, without pulling or removing feathers |
| Severe feather pecking (SFP) | Forceful pecking at or plucking out feathers of other birds |
| Aggressive pecking (AP) | Forceful, upward–downward pecking directed against the head of another bird |
| EP | GFP | SFP | AP | |
|---|---|---|---|---|
| Chick Paper | Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD |
| Available | 0.3633 ± 0.1974 | 0.0675 ± 0.0848 | 0.0138 ± 0.0240 | 0.0094 ± 0.0201 |
| Partially available | 0.4980 ± 0.2688 | 0.1160 ± 0.1184 | 0.0392 ± 0.0785 | 0.0038 ± 0.0130 |
| not available | 0.7686 ± 0. 3770 | 0.0910 ± 0.0960 | 0.0338 ± 0.0772 | 0.0062 ± 0.0183 |
| Functional Area | ||||
| Wired Slats | n.a. 1 | 0.1522 ± 0.1021 | 0.0487 ± 0.0806 | 0.0114 ± 0.0226 |
| Enrichment | 0.5478 ± 0.3361 | 0.0234 ± 0.0349 | 0.0053 ± 0.0163 | 0.0021 ± 0.0092 |
| Perches | n.a. 1 | 0.0150 ± 0.0241 | 0.0017 ± 0.0063 | 0.0010 ± 0.0048 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarzer, A.; Erhard, M.; Schmidt, P.; Zismann, M.; Louton, H. Effects of Stocking Rate and Environmental Enrichment on the Ontogeny of Pecking Behavior of Laying Hen Pullets Confined in Aviary Compartments during the First 4 Weeks of Life. Animals 2022, 12, 2639. https://doi.org/10.3390/ani12192639
Schwarzer A, Erhard M, Schmidt P, Zismann M, Louton H. Effects of Stocking Rate and Environmental Enrichment on the Ontogeny of Pecking Behavior of Laying Hen Pullets Confined in Aviary Compartments during the First 4 Weeks of Life. Animals. 2022; 12(19):2639. https://doi.org/10.3390/ani12192639
Chicago/Turabian StyleSchwarzer, Angela, Michael Erhard, Paul Schmidt, Miriam Zismann, and Helen Louton. 2022. "Effects of Stocking Rate and Environmental Enrichment on the Ontogeny of Pecking Behavior of Laying Hen Pullets Confined in Aviary Compartments during the First 4 Weeks of Life" Animals 12, no. 19: 2639. https://doi.org/10.3390/ani12192639
APA StyleSchwarzer, A., Erhard, M., Schmidt, P., Zismann, M., & Louton, H. (2022). Effects of Stocking Rate and Environmental Enrichment on the Ontogeny of Pecking Behavior of Laying Hen Pullets Confined in Aviary Compartments during the First 4 Weeks of Life. Animals, 12(19), 2639. https://doi.org/10.3390/ani12192639