



Glycosylation Analysis of Feline Small Intestine Following Toxoplasma gondii Infection

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Laboratory Animals
2.3. Confirmation of Infection Model Establishment
2.4. Total Protein Extraction and Digestion
2.5. Enrichment of N-glycosylated Peptides and Deglycosylation
2.6. High-Performance Liquid Chromatography Coupled to Tandem Mass Spectrometry
2.7. Enrichment of N-glycosylated Peptides and Deglycosylation
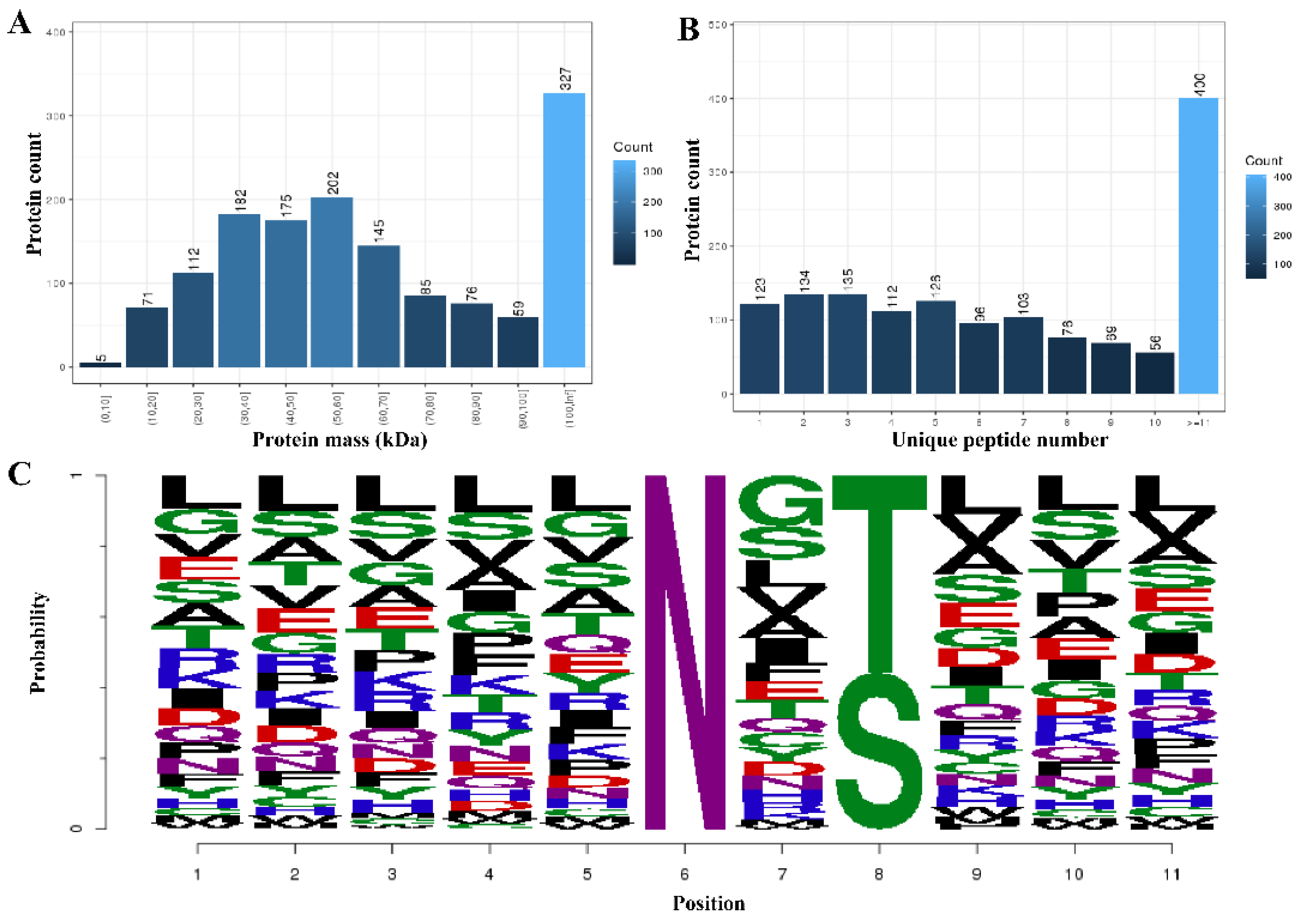
3. Results
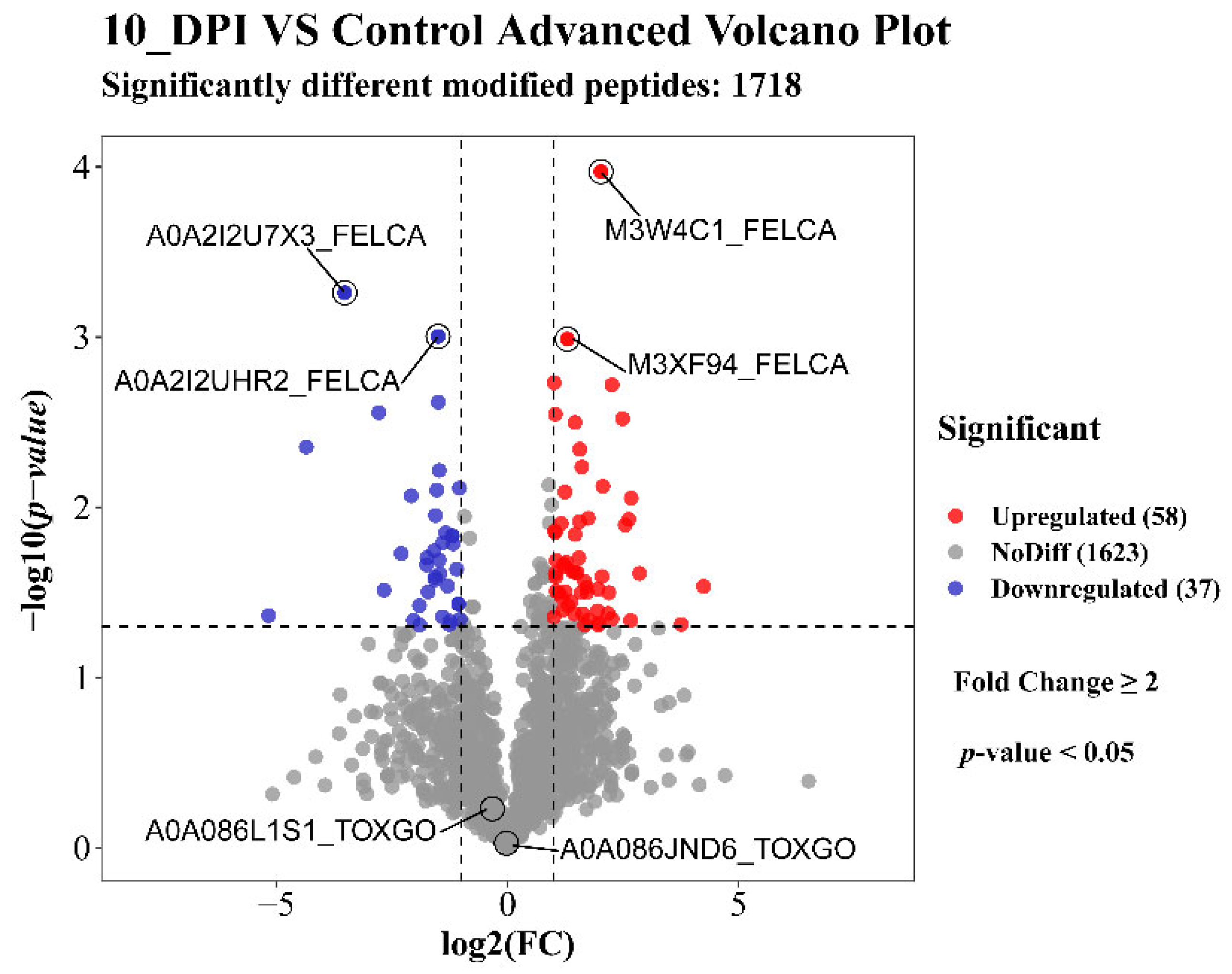
3.1. Differential Quantitative Analysis of the Modified Peptides/Proteins
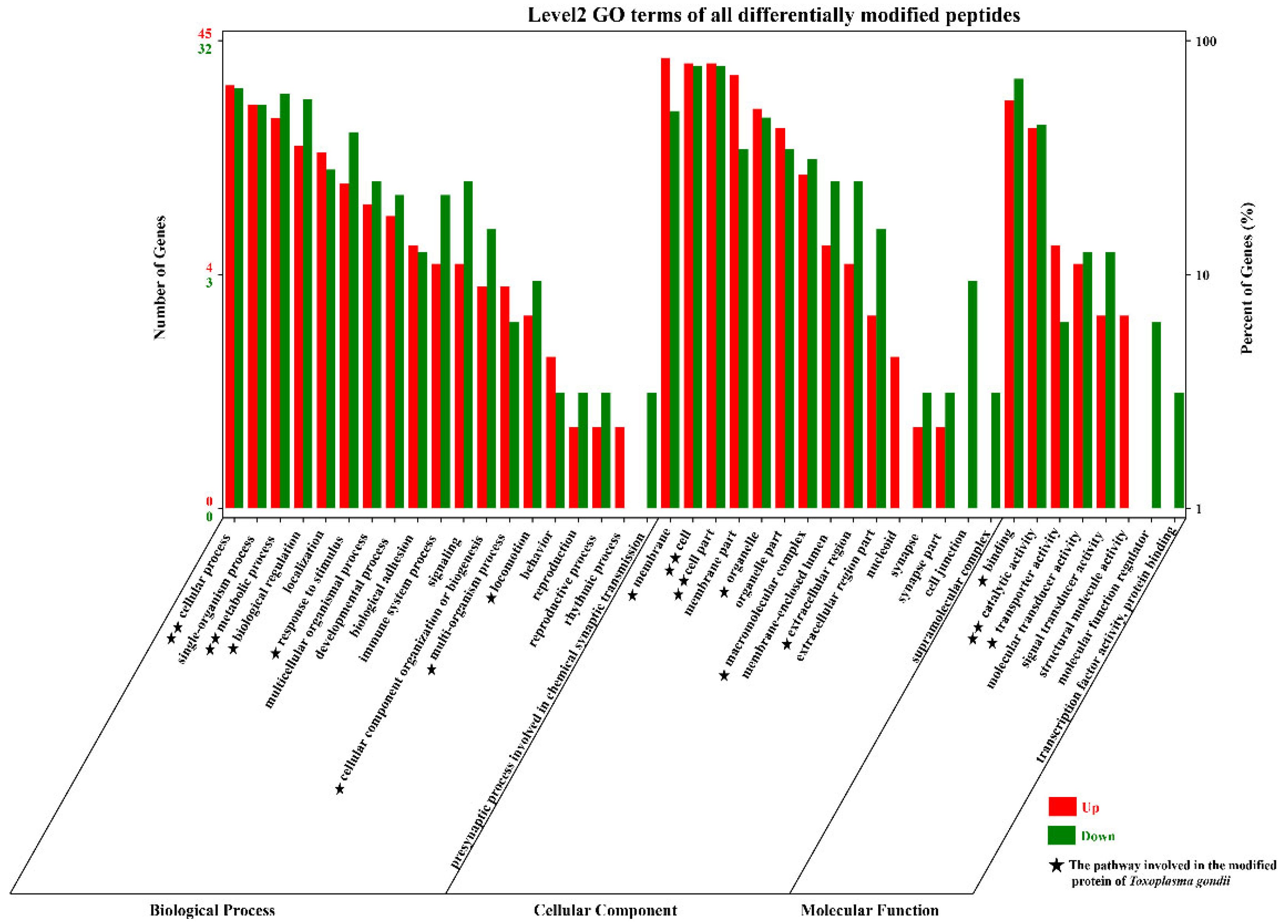
3.2. Functional Annotation and Enrichment Analysis of the Proteins with Differentially Modified Peptides
3.3. PPI Networks for Differentially Modified Peptides/Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yarovinsky, F. Innate immunity to Toxoplasma gondii infection. Nat. Rev. Immunol. 2014, 14, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.D.; Wang, S.C.; Liu, H.H.; Ma, H.Y.; Li, Z.Y.; Wei, F.; Zhu, X.Q.; Liu, Q. Prevalence and burden of Toxoplasma gondii infection in HIV-infected people: A systematic review and meta-analysis. Lancet HIV 2017, 4, e177–e188. [Google Scholar] [CrossRef]
- Moncada, P.A.; Montoya, J.G. Toxoplasmosis in the fetus and newborn: An update on prevalence, diagnosis and treatment. Expert. Rev. Anti-Infect. Ther. 2014, 10, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.C.; Goulart, C.; Hayward, J.A.; Kupz, A.; Miller, C.M.; van Dooren, G.G. Control of human toxoplasmosis. Int. J. Parasitol. 2021, 51, 95–121. [Google Scholar] [CrossRef]
- Dubey, J.P.; Cerqueira-Cezar, C.K.; Murata, F.; Kwok, O.; Yang, Y.R.; Su, C. All about toxoplasmosis in cats: The last decade. Vet. Parasitol. 2020, 283, 109145. [Google Scholar] [CrossRef]
- Di Genova, B.M.; Wilson, S.K.; Dubey, J.P.; Knoll, L.J. Intestinal delta-6-desaturase activity determines host range for Toxoplasma sexual reproduction. PLoS Biol. 2019, 17, e3000364. [Google Scholar]
- Zulpo, D.L.; Sammi, A.S.; Dos, S.J.R.; Sasse, J.P.; Martins, T.A.; Minutti, A.F.; Cardim, S.T.; de Barros, L.D.; Navarro, I.T.; Garcia, J.L. Toxoplasma gondii: A study of oocyst re-shedding in domestic cats. Vet. Parasitol. 2018, 249, 17–20. [Google Scholar] [CrossRef]
- Shapiro, K.; Bahia-Oliveira, L.; Dixon, B.; Dumetre, A.; de Wit, L.A.; VanWormer, E.; Villena, I. Environmental transmission of Toxoplasma gondii: Oocysts in water, soil and food. Food Waterborne Parasitol. 2019, 15, e00049. [Google Scholar] [CrossRef]
- Alizadeh, A.M.; Jazaeri, S.; Shemshadi, B.; Hashempour-Baltork, F.; Sarlak, Z.; Pilevar, Z.; Hosseini, H. A review on inactivation methods of Toxoplasma gondii in foods. Pathog. Glob. Health 2018, 112, 306–319. [Google Scholar] [CrossRef]
- Gomord, V.; Fitchette, A.C.; Menu-Bouaouiche, L.; Saint-Jore-Dupas, C.; Plasson, C.; Michaud, D.; Faye, L. Plant-specific glycosylation patterns in the context of therapeutic protein production. Plant Biotechnol. J. 2010, 8, 564–587. [Google Scholar] [CrossRef]
- Lauc, G.; Huffman, J.E.; Pučić, M.; Zgaga, L.; Adamczyk, B.; Mužinić, A.; Novokmet, M.; Polašek, O.; Gornik, O.; Krištić, J.; et al. Loci associated with N-glycosylation of human immunoglobulin G show pleiotropy with autoimmune diseases and haematological cancers. PLoS Genet. 2013, 9, e1003225. [Google Scholar] [CrossRef]
- Hwang, H.; Zhang, J.P.; Chung, K.A.; Leverenz, J.B.; Zabetian, C.P.; Peskind, E.R.; Jankovic, J.; Su, Z.; Hancock, A.M.; Pan, C.; et al. Glycoproteomics in neurodegenerative diseases. Mass Spectrum. Rev. 2010, 29, 79–125. [Google Scholar] [CrossRef] [Green Version]
- Zaia, J. Mass spectrometry and the emerging field of glycomics. Chem. Biol. 2008, 15, 881–892. [Google Scholar] [CrossRef] [Green Version]
- Luk, F.C.; Johnson, T.M.; Beckers, C.J. N-linked glycosylation of proteins in the protozoan parasite Toxoplasma gondii. Mol. Biochem. Parasitol. 2008, 157, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Nazarova, L.A.; Ochoa, R.J.; Jones, K.A.; Morrissette, N.S.; Prescher, J.A. Extracellular Toxoplasma gondii tachyzoites metabolize and incorporate unnatural sugars into cellular proteins. Microbes Infect. 2016, 18, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Vishwanath, P.; Cui, J.; Kelleher, D.J.; Gilmore, R.; Robbins, P.W.; Samuelson, J. The evolution of N-glycan-dependent endoplasmic reticulum quality control factors for glycoprotein folding and degradation. Proc. Natl. Acad. Sci. USA 2007, 104, 11676–11681. [Google Scholar] [CrossRef] [Green Version]
- Helenius, A.; Aebi, M. Roles of N-linked glycans in the endoplasmic reticulum. Annu. Rev. Biochem. 2004, 73, 1019–1049. [Google Scholar] [CrossRef]
- Bushkin, G.G.; Ratner, D.M.; Cui, J.; Banerjee, S.; Duraisingh, M.T.; Jennings, C.V.; Dvorin, J.D.; Gubbels, M.J.; Robertson, S.D.; Steffen, M.; et al. Suggestive evidence for darwinian selection against asparagine-linked glycans of Plasmodium falciparum and Toxoplasma gondii. Eukaryot. Cell 2010, 9, 228–241. [Google Scholar] [CrossRef] [Green Version]
- Fauquenoy, S.; Hovasse, A.; Sloves, P.J.; Morelle, W.; Alayi, T.D.; Slomianny, C.; Werkmeister, E.; Schaeffer, C.; Dorsselaer, A.V.; Tomavo, S. Unusual N-glycan structures required for trafficking Toxoplasma gondii GAP50 to the inner membrane complex regulate host cell entry through parasite motility. Mol. Cell. Proteom. 2011, 10, M111.008953. [Google Scholar]
- Gaskins, E.; Gilk, S.; DeVore, N.; Mann, T.; Ward, G.; Beckers, C. Identification of the membrane receptor of a class XIV myosin in Toxoplasma gondii. J. Cell Biol. 2004, 165, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Rani, S.; Cerqueira-Cezar, C.K.; Murata, F.; Sadler, M.; Kwok, O.; Pradhan, A.K.; Hill, D.E.; Urban, J.J.; Dubey, J.P. Toxoplasma gondii tissue cyst formation and density of tissue cysts in shoulders of pigs 7 and 14 days after feeding infected mice tissues. Vet. Parasitol. 2019, 269, 13–15. [Google Scholar] [CrossRef]
- Ma, J.; He, J.J.; Hou, J.L.; Zhou, C.X.; Zhang, F.K.; Elsheikha, H.M.; Zhu, X.Q. Metabolomic signature of mouse cerebral cortex following Toxoplasma gondii infection. Parasit. Vectors 2019, 12, 373. [Google Scholar] [CrossRef]
- Zhou, C.X.; Zhou, D.H.; Elsheikha, H.M.; Liu, G.X.; Suo, X.; Zhu, X.Q. Global metabolomic profiling of mice brains following experimental infection with the cyst-forming Toxoplasma gondii. PLoS ONE 2015, 10, e139635. [Google Scholar]
- Wu, D.; Xue, X.W.; Gao, C.C.; Liu, Y.H.; Wang, T.T.; Li, L.S.; Tong, X.H.; Li, F.; Xu, J.D. Rhubarb-Evoke mucus secretion through aggregation and degranulation of mast cell in the colon of rat: In vivo and ex vivo studies. Sci. Rep. 2019, 9, 19375. [Google Scholar] [CrossRef] [Green Version]
- Haider, S.R.; Reid, H.J.; Sharp, B.L. Tricine-SDS-PAGE. Methods Mol. Biol. 2019, 1855, 151–160. [Google Scholar]
- Zappacosta, F.; Scott, G.F.; Huddleston, M.J.; Annan, R.S. An optimized platform for hydrophilic interaction chromatography-immobilized metal affinity chromatography enables deep coverage of the rat liver phosphoproteome. J. Proteome Res. 2015, 14, 997–1009. [Google Scholar] [CrossRef]
- Cole, J.; Hanson, E.J.; James, D.C.; Dockrell, D.H.; Dickman, M.J. Comparison of data-acquisition methods for the identification and quantification of histone post-translational modifications on a Q Exactive HF hybrid quadrupole Orbitrap mass spectrometer. Rapid Commun. Mass Spectrom. 2019, 33, 897–906. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, K.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Pedrazzini, E.; Caprera, A.; Fojadelli, I.; Stella, A.; Rocchetti, A.; Bassin, B.; Martinoia, E.; Vitale, A. The Arabidopsis tonoplast is almost devoid of glycoproteins with complex N-glycans, unlike the rat lysosomal membrane. J. Exp. Bot. 2016, 67, 1769–1781. [Google Scholar] [CrossRef] [Green Version]
- Wiederschain, G.Y. Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2009. [Google Scholar]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A.V. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.H.; Jia, L.S.; Guo, H.W.; Ding, H.Y.; Yang, J.Y.; Wang, H.W. Eukaryotic elongation factor 2 is involved in the anticoccidial action of diclazuril in the second-generation merozoites of Eimeria tenella. Vet. Parasitol. 2019, 276, 108991. [Google Scholar] [CrossRef] [PubMed]
- Florimond, C.; Cordonnier, C.; Taujale, R.; van der Wel, H.; Kannan, N.; West, C.M.; Blader, I.J. A toxoplasma prolyl hydroxylase mediates oxygen stress responses by regulating translation elongation. mBio 2019, 10, e00234-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barylyuk, K.; Koreny, L.; Ke, H.; Butterworth, S.; Crook, O.M.; Lassadi, I.; Gupta, V.; Tromer, E.; Mourier, T.; Stevens, T.J.; et al. Comprehensive subcellular atlas of the Toxoplasma proteome via hyperLOPIT provides spatial context for protein functions. Cell Host Microbe 2020, 28, 752–766.e9. [Google Scholar] [CrossRef]
- Liu, Y.M.; Zou, X.Q.; Ou, M.L.; Ye, X.H.; Zhang, B.H.; Wu, T.Y.; Dong, S.R.; Chen, X.J.; Liu, H.X.; Zheng, Z.; et al. Toxoplasma gondii cathepsin C1 inhibits NF-kappaB signalling through the positive regulation of the HIF-1alpha/EPO axis. Acta Trop. 2019, 195, 35–43. [Google Scholar] [CrossRef]
- Mendonça-Natividade, F.C.; Duque, L.C.; Ricci-Azevedo, R.; Sardinha-Silva, A.; Figueiredo, P.C.; Paiva, A.A.; Alegre-Maller, A.C.P.; Nohara, L.L.; Carneiro, A.B.; Panunto-Castelo, A.; et al. Receptor heterodimerization and co-receptor engagement in TLR2 activation induced by MIC1 and MIC4 from Toxoplasma gondii. Int. J. Mol. Sci. 2019, 20, 5001. [Google Scholar] [CrossRef] [Green Version]
- Bichet, M.; Joly, C.; Henni, A.H.; Guilbert, T.; Xemard, M.; Tafani, V.; Lagal, V.; Charras, G.; Tardieux, I. The toxoplasma-host cell junction is anchored to the cell cortex to sustain parasite invasive force. BMC Biol. 2014, 12, 773. [Google Scholar] [CrossRef] [Green Version]
- Denkers, E.Y.; Bzik, D.J.; Fox, B.A.; Butcher, B.A. An inside job: Hacking into Janus kinase/signal transducer and activator of transcription signaling cascades by the intracellular protozoan Toxoplasma gondii. Infect. Immun. 2012, 80, 476–482. [Google Scholar] [CrossRef] [Green Version]
- Nast, R.; Staab, J.; Meyer, T.; Luder, C. Toxoplasma gondii stabilises tetrameric complexes of tyrosine-phosphorylated signal transducer and activator of transcription-1 and leads to its sustained and promiscuous DNA binding. Cell Microbiol. 2018, 20, e12887. [Google Scholar] [CrossRef]
- Figueras, M.J.; Martin, O.A.; Echeverria, P.C.; de Miguel, N.; Naguleswaran, A.; Sullivan, W.J.; Corvi, M.M.; Angel, S.O. Toxoplasma gondii Sis1-like J-domain protein is a cytosolic chaperone associated to HSP90/HSP70 complex. Int. J. Biol. Macromol. 2012, 50, 725–733. [Google Scholar] [CrossRef] [Green Version]
- Weiss, L.M.; Ma, Y.F.; Takvorian, P.M.; Tanowitz, H.B.; Wittner, M. Bradyzoite development in Toxoplasma gondii and the hsp70 stress response. Infect. Immun. 1998, 66, 3295–3302. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.; Monaghan, P.; Page, A.P. Peptidyl-prolyl cis-trans isomerases (immunophilins) and their roles in parasite biochemistry, host-parasite interaction and antiparasitic drug action. Int. J. Parasitol. 2006, 36, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Silverman, J.A.; Hayes, M.L.; Luft, B.J.; Joiner, K.A. Characterization of anti-Toxoplasma activity of SDZ 215–918, a cyclosporin derivative lacking immunosuppressive and peptidyl-prolyl-isomerase-inhibiting activity: Possible role of a P glycoprotein in Toxoplasma physiology. Antimicrob. Agents Chemother. 1997, 41, 1859–1866. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Zhou, D.H.; Chen, J.; Zhang, N.Z.; Wang, R.A.; Weng, Y.B.; Zhu, X.Q. Characterization of the Toxoplasma gondii hsp60 gene sequences from different hosts and geographical locations. Genet. Mol. Res. 2014, 13, 6906–6911. [Google Scholar] [CrossRef]
- Soto, A.S.; Fenoy, I.M.; Sanchez, V.R.; March, F.; Sibilia, M.D.P.; Aldirico, M.D.L.A.; Picchio, M.S.; Arcon, N.; Acosta, P.; Polack, F.P.; et al. Toxoplasma gondii serine-protease inhibitor-1: A new adjuvant candidate for asthma therapy. PLoS ONE 2017, 12, e0187002. [Google Scholar] [CrossRef] [Green Version]
- Pszenny, V.; Davis, P.H.; Zhou, X.W.; Hunter, C.A.; Carruthers, V.B.; Roos, D.S. Targeted disruption of Toxoplasma gondii serine protease inhibitor 1 increases bradyzoite cyst formation in vitro and parasite tissue burden in mice. Infect. Immun. 2012, 80, 1156–1165. [Google Scholar] [CrossRef] [Green Version]
- Dubois, D.; Fernandes, S.; Amiar, S.; Dass, S.; Katris, N.J.; Botte, C.Y.; Yamaryo-Botte, Y. Toxoplasma gondii acetyl-CoA synthetase is involved in fatty acid elongation (of long fatty acid chains) during tachyzoite life stages. J. Lipid Res. 2018, 59, 994–1004. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, S.; Docampo, M.D.; MacRae, J.I.; Ralton, J.E.; Rupasinghe, T.; McConville, M.J.; Striepen, B. The intracellular parasite Toxoplasma gondii depends on the synthesis of long-chain and very long-chain unsaturated fatty acids not supplied by the host cell. Mol. Microbiol. 2015, 97, 64–76. [Google Scholar] [CrossRef] [Green Version]
- Filipowicz, A.; Coca, M.N.; Blair, B.M.; Chang, P.Y. Acute myocarditis with cardiogenic shock and multiple organ failure, followed by bilateral panuveitis masquerading as endogenous endophthalmitis, due to Toxoplasma gondii in an immunocompetent patient. Retin. Cases Brief Rep. 2021, 15, 575–580. [Google Scholar] [CrossRef]
- Sauvant, G.; Bossart, W.; Kurrer, M.O.; Follath, F. Diagnosis and course of myocarditis: A survey in the medical clinics of Zurich university hospital 1980 to 1998. Schweiz. Med. Wochenschr. 2000, 130, 1265–1271. [Google Scholar]
- Barragan, A.; Brossier, F.; Sibley, L.D. Transepithelial migration of Toxoplasma gondii involves an interaction of intercellular adhesion molecule 1 (ICAM-1) with the parasite adhesin MIC2. Cell. Microbiol. 2005, 7, 561–568. [Google Scholar] [CrossRef]
- Bushkin, G.G.; Motari, E.; Carpentieri, A.; Dubey, J.P.; Costello, C.E.; Robbins, P.W.; Samuelson, J. Evidence for a structural role for acid-fast lipids in oocyst walls of Cryptosporidium, Toxoplasma, and Eimeria. mBio 2013, 4, e00387-13. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.M.; Berry, L.; Sullivan, W.J.; Besteiro, S. Autophagy participates in the unfolded protein response in Toxoplasma gondii. FEMS Microbiol. Lett. 2017, 364, fnx153. [Google Scholar] [CrossRef] [Green Version]
- Joyce, B.R.; Tampaki, Z.; Kim, K.; Wek, R.C.; Sullivan, W.J. The unfolded protein response in the protozoan parasite Toxoplasma gondii features translational and transcriptional control. Eukaryot. Cell 2013, 12, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Amara, N.; Foe, I.T.; Onguka, O.; Garland, M.; Bogyo, M. Synthetic fluorogenic peptides reveal dynamic substrate specificity of depalmitoylases. Cell Chem. Biol. 2019, 26, 35–47. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total_Spectra | Identified_Spectra | Proteins | Glycoproteins a | Peptides | Glycopeptides b | Glycosites |
|---|---|---|---|---|---|---|---|
| 10_DPI_1 | 287,461 | 1510 | 209 | 4 | 290 | 4 | 4 |
| 10_DPI_2 | 288,686 | 1622 | 171 | 4 | 259 | 5 | 4 |
| 10_DPI_3 | 281,373 | 1491 | 165 | 3 | 240 | 4 | 3 |
| No. | Pathway | Different Proteins with Pathway Annotation | Proteins | Pathway ID |
|---|---|---|---|---|
| 1 | Ribosome | 1 (50%) | tr|A0A086K8H6|A0A086K8H6_TOXGO | ko03010 |
| 2 | AMPK signaling pathway | 1 (50%) | tr|A0A086JND6|A0A086JND6_TOXGO | ko04152 |
| 3 | Oxytocin signaling pathway | 1 (50%) | tr|A0A086JND6|A0A086JND6_TOXGO | ko04921 |
| 4 | Amino sugar and nucleotide sugar metabolism | 1 (25%) | tr|A0A151HN22|A0A151HN22_TOXGO | ko00520 |
| 5 | Aminoacyl-tRNA biosynthesis | 1 (25%) | tr|A0A125YXZ8|A0A125YXZ8_TOXGV | ko00970 |
| 6 | Ribosome biogenesis in eukaryotes | 1 (25%) | tr|A0A086KYF0|A0A086KYF0_TOXGO | ko03008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, B.; Xie, S.; Peng, J.; Qiu, Y.; Liu, Y.; Zhu, X.; He, J.; Zhang, J. Glycosylation Analysis of Feline Small Intestine Following Toxoplasma gondii Infection. Animals 2022, 12, 2858. https://doi.org/10.3390/ani12202858
Zhai B, Xie S, Peng J, Qiu Y, Liu Y, Zhu X, He J, Zhang J. Glycosylation Analysis of Feline Small Intestine Following Toxoplasma gondii Infection. Animals. 2022; 12(20):2858. https://doi.org/10.3390/ani12202858
Chicago/Turabian StyleZhai, Bintao, Shichen Xie, Junjie Peng, Yanhua Qiu, Yang Liu, Xingquan Zhu, Junjun He, and Jiyu Zhang. 2022. "Glycosylation Analysis of Feline Small Intestine Following Toxoplasma gondii Infection" Animals 12, no. 20: 2858. https://doi.org/10.3390/ani12202858
APA StyleZhai, B., Xie, S., Peng, J., Qiu, Y., Liu, Y., Zhu, X., He, J., & Zhang, J. (2022). Glycosylation Analysis of Feline Small Intestine Following Toxoplasma gondii Infection. Animals, 12(20), 2858. https://doi.org/10.3390/ani12202858