
Role of Guinea Pigs (Cavia porcellus) Raised as Livestock in Ecuadorian Andes as Reservoirs of Zoonotic Yeasts
 ,
,  ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
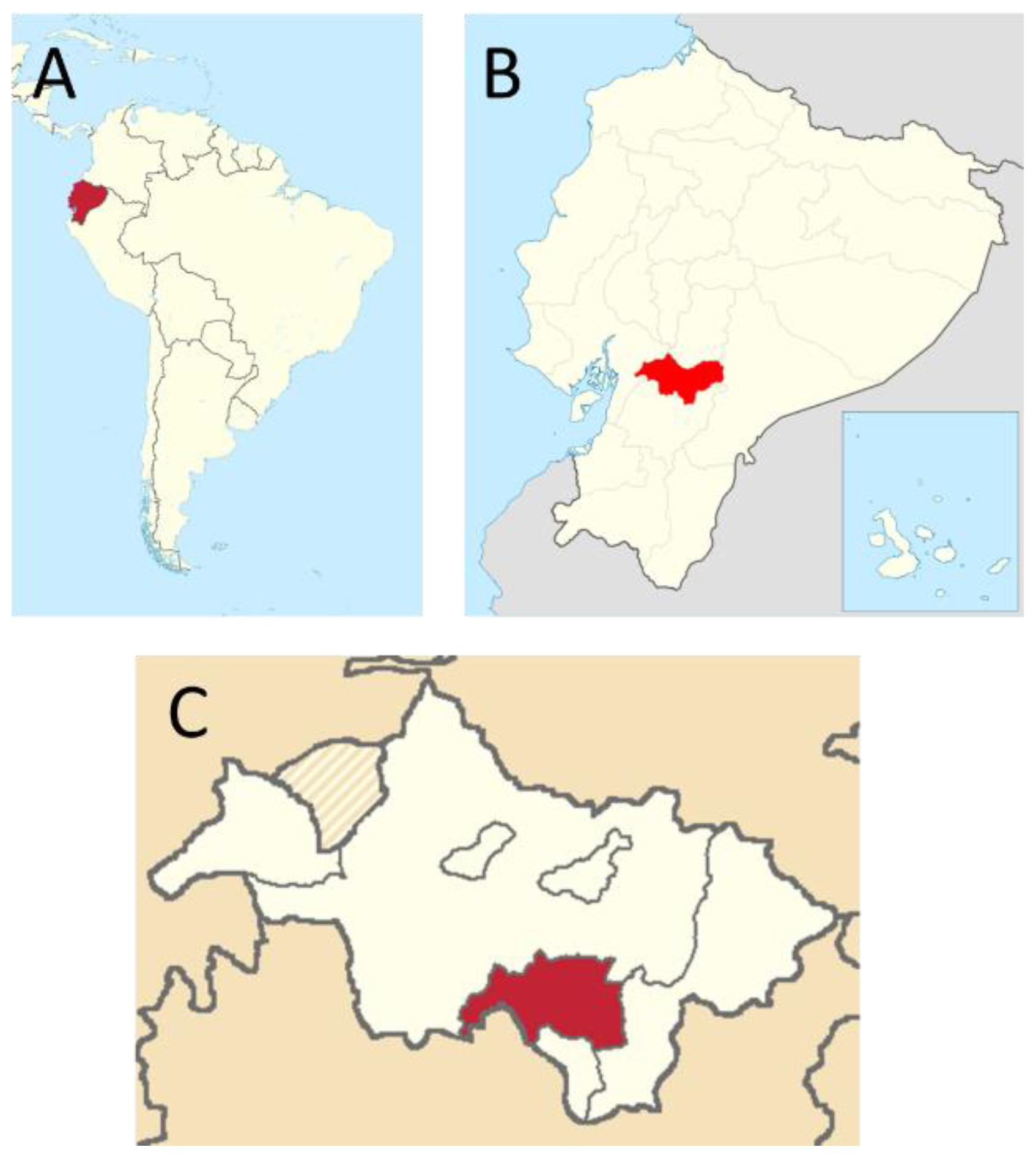
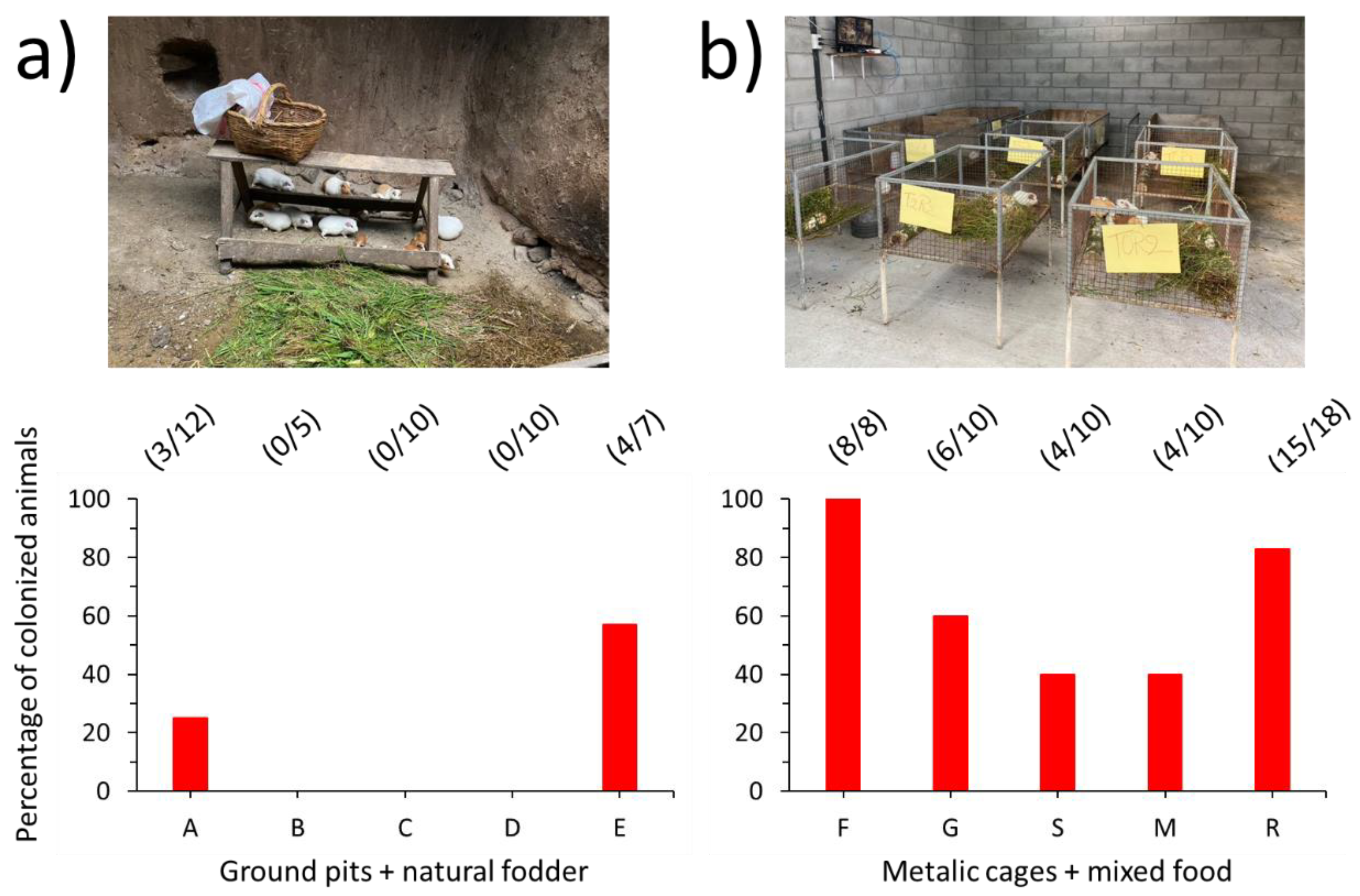
2.1. Sampling Area and Animals
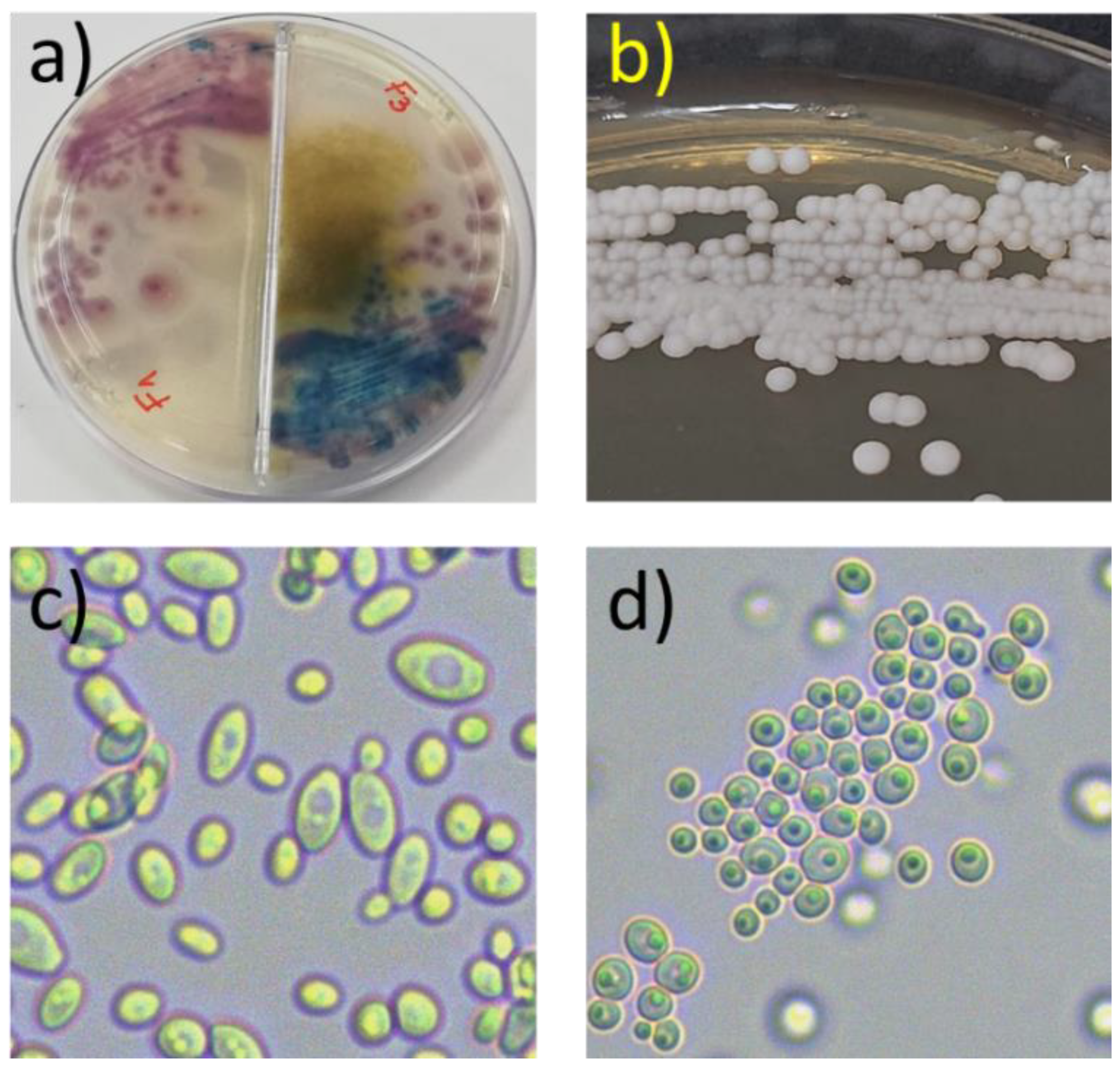
2.2. Sample Collection and Isolation Procedures
2.3. Antifungal Susceptibility Tests
2.4. DNA Extraction and Molecular Identification of the Isolates
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Avilés, D.; Martínez, A.; Landi, V.; Delgado, J. The guinea pig (Cavia porcellus): An Andean resource of interest as an agricultural food source. Anim. Genet. Res. 2014, 55, 87–91. [Google Scholar] [CrossRef]
- Vasco, K.; Graham, J.P.; Trueba, G. Detection of zoonotic enteropathogens in children and domestic animals in a semirural community in Ecuador. Appl. Environ. Microbiol. 2016, 82, 4218–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Zaldívar, L.C. Producción de Cuyes (Cavia porcellus) en los Países Andinos. Food and Agriculture Organization Document. Available online: https://www.fao.org/3/V6200T/v6200T05.htm (accessed on 2 October 2022).
- Graham, J.P.; Vasco, K.; Trueba, G. Hyperendemic Campylobacter jejuni in guinea pigs (Cavia porcellus) raised for food in a semi-rural community of Quito, Ecuador. Environ. Microbiol. Rep. 2016, 8, 382–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amman, B.R.; Pavlin, B.I.; Albarino, C.G.; Comer, J.A.; Erickson, B.R.; Oliver, J.B.; Sealy, T.K.; Vincent, M.J.; Nichol, S.T.; Paddock, C.D.; et al. Pet rodents and fatal lymphocytic choriomeningitis in transplant patients. Emerg. Infect. Dis. 2007, 13, 719–725. [Google Scholar] [CrossRef]
- Lutz-Wohlgroth, L.; Becker, A.; Brugnera, E.; Huat, Z.L.; Zimmermann, D.; Grimm, F.; Haessig, M.; Greub, G.; Kaps, S.; Spiess, B.; et al. Chlamydiales in guinea-pigs and their zoonotic potential. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2006, 53, 185–193. [Google Scholar] [CrossRef]
- Fredriksson-Ahomaa, M. Yersinia enterocolitica and Yersinia pseudotuberculosis. In Foodborne Diseases; Simjee, S., Ed.; Humana Press: New York, NY, USA, 2007; pp. 79–113. [Google Scholar]
- Gruszynski, K.; Young, A.; Levine, S.J.; Garvin, J.P.; Brown, S.; Turner, L.; Fritzinger, A.; Gertz, R.E., Jr.; Murphy, J.M.; Vogt, M.; et al. Streptococcus equi subsp zooepidemicus infections associated with guinea pigs. Emerg. Infect. Dis. 2015, 21, 156–158. [Google Scholar] [CrossRef]
- Walther, B.; Wieler, L.H.; Vincze, S.; Antão, E.M.; Brandenburg, A.; Stamm, I.; Kopp, P.A.; Kohn, B.; Semmler, T.; Lübke-Becker, A. MRSA variantin companion animals. Emerg. Infect. Dis. 2012, 18, 2017–2020. [Google Scholar] [CrossRef]
- González, L.C.; Esteban, J.G.; Bargues, M.D.; Valero, M.A.; Ortiz, P.; Naquira, C.; Mas-Coma, S. Hyper-endemic human fascioliasis in Andean valleys: An altitudinal transect analysis in children of Cajamarca province, Peru. Acta Trop. 2011, 120, 119–129. [Google Scholar] [CrossRef]
- Gabastou, J.M.; Proano, J.; Vimos, A.; Jaramillo, G.; Hayes, E.; Gage, K.; Chu, M.; Guarner, J.; Zaki, S.; Bowers, J.; et al. An outbreak of plague including cases with probable pneumonic infection, Ecuador, 1998. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 387–391. [Google Scholar] [CrossRef]
- Levy, M.Z.; Bowman, N.M.; Kawai, V.; Waller, L.A.; Cornejo del Carpio, J.G.; Cordova Benzaquen, E.; Gilman, R.H.; Bern, C. Periurban Trypanosoma cruzi-infected Triatoma infestans, Arequipa, Peru. Emerg. Infect. Dis. 2006, 12, 1345–1352. [Google Scholar] [CrossRef]
- Zambrano-Mila, M.; Rodriguez, A.S.; Rivera-Olivero, I.A.; Salas-Rueda, M.; Caceres-Orellana, M.V.; de Waard, J.H.; Garcia-Bereguiain, M.A. Methicillin resistant Staphylococcus aureus carriage among guinea pigs raised as livestock in Ecuador. One Health 2020, 9, 100118. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Grado, V.H.; Mubareka, S.; Krammer, F.; Cárdenas, W.B.; Palese, P. Influenza virus infection in Guinea pigs raised as livestock, Ecuador. Emerg. Infect. Dis. 2012, 18, 1135–1137. [Google Scholar] [CrossRef] [Green Version]
- Miceli, M.H.; Díaz, J.A.; Lee, S.A. Emerging opportunistic yeast infections. Lancet Infect. Dis. 2011, 1, 142–151. [Google Scholar] [CrossRef]
- WHO. Managing Zoonotic Public Health Risks at the Human Animal–Ecosystem Interface; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- Seyedmousavi, S.; Guillot, J.; Tolooe, A.; Verweij, P.E.; de Hoog, G.S. Neglected fungal zoonoses: Hidden threats to man and animals. Clin. Microbiol. Infect. 2015, 21, 416–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otte, J.; Pica-Ciamarra, U. Emerging infectious zoonotic diseases: The neglected role of food animals. One Health 2021, 13, 100323. [Google Scholar] [CrossRef] [PubMed]
- Brilhante, R.S.; Bittencourt, P.V.; Castelo-Branco, D.; de Oliveira, J.S.; Alencar, L.P.; Cordeiro, R.; Pinheiro, M.; Nogueira-Filho, E.F.; Pereira-Neto, W.; Sidrim, J.J.; et al. Trends in antifungal susceptibility and virulence of Candida spp. from the nasolacrimal duct of horses. Med. Mycol. 2016, 54, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glushakova, A.M.; Rodionova, E.N.; Kachalkin, A.V. Yeasts in feces of pigeons (Columba Livia) in the city of Moscow. Curr. Microbiol. 2021, 78, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Rosario Medina, I.; Román Fuentes, L.; Batista Arteaga, M.; Real Valcárcel, F.; Acosta Arbelo, F.; Padilla Del Castillo, D.; Déniz Suárez, S.; Ferrer Quintana, O.; Vega Gutiérrez, B.; Silva Sergent, F.; et al. Pigeons and their droppings as reservoirs of Candida and other zoonotic yeasts. Rev. Iberoam. Micol. 2017, 34, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Giacobbe, D.R.; Maraolo, A.E.; Simeon, V.; Magnè, F.; Pace, M.C.; Gentile, I.; Chiodini, P.; Viscoli, C.; Sanguinetti, M.; Mikulska, M.; et al. Changes in the relative prevalence of candidaemia due to non-albicans Candida species in adult in-patients: A systematic review, meta-analysis and meta-regression. Mycoses 2020, 63, 334–342. [Google Scholar] [CrossRef]
- Papon, N.; Savini, V.; Lanoue, A.; Simkin, A.J.; Crèche, J.; Giglioli-Guivarc’h, N.; Clastre, M.; Courdavault, V.; Sibirny, A.A. Candida guilliermondii: Biotechnological applications, perspectives for biological control, emerging clinical importance and recent advances in genetics. Curr. Genet. 2013, 59, 73–90. [Google Scholar] [CrossRef]
- Ley Orgánica de Sanidad Agropecuaria. Asamblea Nacional, República del Ecuador. Available online: https://www.gob.ec/sites/default/files/regulations/2018-09/Documento_Ley%20Org%C3%A1nica%20de%20Sanidad%20Agropecuaria.pdf (accessed on 11 November 2022).
- Clinical and Laboratory Standards Institute (CLSI). Method for Antifungal Disk Diffusion Susceptibility Testing of Yeasts, 3rd ed.; CLSI Guideline M44; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Almeida da Silva, G.; Bernardi, T.L.; Carvalho Schaker, P.D.; Menegotto, M.; Valente, P. Rapid yeast DNA extraction by boiling and freeze-thawing without using chemical reagents and DNA purification. Braz. Arch. Biol. Technol. 2012, 55, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes: Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols a Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Opulente, D.A.; Langdon, Q.K.; Buh, K.V.; Haase, M.A.B.; Sylvester, K.; Moriarty, R.V.; Jarzyna, M.; Considine, S.L.; Schneider, R.M.; Hittinger, C.T. Pathogenic budding yeasts isolated outside of clinical settings. FEMS Yeast Res. 2019, 19, foz032. [Google Scholar] [CrossRef] [PubMed]
- Deorukhkar, S.C.; Saini, S.; Mathew, S. Non-albicans Candida infection: An emerging threat. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 615958. [Google Scholar] [CrossRef] [Green Version]
- Farmakiotis, D.; Kontoyiannis, D.P. Epidemiology of antifungal resistance in human pathogenic yeasts: Current viewpoint and practical recommendations for management. Int. J. Antimicrob. Agents 2017, 50, 318–324. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Rodloff, A.; Fu, W.; Ling, T.A. Global Antifungal Surveillance Group. Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2007: A 10.5-year analysis of susceptibilities of Candida Species to fluconazole and voriconazole as determined by CLSI standardized disk diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377. [Google Scholar] [CrossRef] [Green Version]
- Botelho, N.S.; Paula, S.B.; Panagio, L.A.; Pinge-Filho, P.; Yamauchi, L.M.; Yamada-Ogatta, S.F. Candida species isolated from urban bats of Londrina-Paraná, Brazil and their potential virulence. Zoonoses Public Health 2012, 59, 16–22. [Google Scholar] [CrossRef]
- Mancianti, F.; Nardoni, S.; Ceccherelli, R. Occurrence of yeasts in psittacines droppings from captive birds in Italy. Mycopathologia 2002, 153, 121–124. [Google Scholar] [CrossRef]
- Zhang, L.; Xiao, M.; Arastehfar, A.; Ilkit, M.; Zou, J.; Deng, Y.; Xu, Y.; Liao, W.; Zhao, J.; Fang, W.; et al. Investigation of the emerging nosocomial Wickerhamomyces anomalus infections at a Chinese tertiary teaching hospital and a systemic review: Clinical manifestations, risk factors, treatment, outcomes, and anti-fungal susceptibility. Front. Microbiol. 2021, 12, 744502. [Google Scholar] [CrossRef]
- Lin, H.C.; Lin, H.Y.; Su, B.H.; Ho, M.W.; Ho, C.M.; Lee, C.Y.; Lin, M.H.; Hsieh, H.Y.; Lin, H.C.; Li, T.C.; et al. Reporting an outbreak of Candida pelliculosa fungemia in a neonatal intensive care unit. J. Microbiol. Immunol. Infect. 2013, 46, 456–462. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Castanheira, M.; Messer, S.A.; Jones, R.N. In vitro antifungal susceptibilities of isolates of Candida spp. and Aspergillus spp. From China to nine systemically active antifungal agents: Data from the SENTRY antifungal surveillance program, 2010 through 2012. Mycoses 2015, 58, 209–214. [Google Scholar] [CrossRef]
- Joo, H.S.; Ndegwa, P.M.; Shoda, M.; Phae, C.G. Bioremediation of oil-contaminated soil using Candida catenulata and food waste. Environ. Pollut. 2008, 156, 891–896. [Google Scholar] [CrossRef]
- Babaei, F.; Habibi, A. Fast biodegradation of diesel hydrocarbons at high concentration by the sophorolipid-producing yeast Candida catenulata KP324968. J. Mol. Microbiol. Biotechnol. 2018, 28, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Cafarchia, C.; Iatta, R.; Danesi, P.; Camarda, A.; Capelli, G.; Otranto, D. Yeasts isolated from cloacal swabs, feces, and eggs of laying hens. Med. Mycol. 2019, 57, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Subramanya, S.H.; Sharan, N.K.; Baral, B.P.; Hamal, D.; Nayak, N.; Prakash, P.Y.; Sathian, B.; Bairy, I.; Gokhale, S. Diversity, in-vitro virulence traits and antifungal susceptibility pattern of gastrointestinal yeast flora of healthy poultry, Gallus domesticus. BMC Microbiol. 2017, 17, 113. [Google Scholar] [CrossRef] [Green Version]
- Boden, S.; Morio, F.; Zhou, M.; Gerrits van den Ende, B.; Hagen, F. De novo Nanopore genome sequencing of the clinical Diutina catenulata type-strain CBS565. Mycopathologia 2022, 187, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Rhimi, W.; Aneke, C.I.; Annoscia, G.; Camarda, A.; Mosca, A.; Cantacessi, C.; Otranto, D.; Cafarchia, C. Virulence and in vitro antifungal susceptibility of Candida albicans and Candida catenulata from laying hens. Int. Microbiol. 2021, 24, 57–63. [Google Scholar] [CrossRef]
- GunTang, W.; Kamonvoradej, N.; Chomchat, C.; Suriyakan, S.; Sanit, S.; Wongwigkarn, J.; Lamlertthon, S. Prevalence and virulence factors of Candida spp. associated with blow flies. Asian Pac. J. Trop. Biomed. 2017, 7, 428–431. [Google Scholar] [CrossRef]
- Radosavljevic, M.; Koenig, H.; Letscher-Bru, V.; Waller, J.; Maloisel, F.; Lioure, B.; Herbrecht, R. Candida catenulata fungemia in a cancer Patient. J. Clin. Microbiol. 1999, 37, 475–477. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.V.; Choy, M.S.; McCoy, D.; Fernandez, N.; Suh, J.S. Candida catenulata candidaemia and possible endocarditis in a cirrhotic patient successfully de-escalated to oral fluconazole. J. Clin. Pharm. Ther. 2018, 43, 910–913. [Google Scholar] [CrossRef]
- Chen, X.-F.; Zhang, W.; Fan, X.; Hou, X.; Liu, X.-Y.; Huang, J.-J.; Kang, W.; Zhang, G.; Zhang, H.; Yang, W.-H.; et al. Antifungal susceptibility profiles and resistance mechanisms of clinical Diutina catenulata isolates with high MIC values. Front. Cell. Infect. Microbiol. 2021, 11, 739496. [Google Scholar] [CrossRef]
- Lamoth, F.; Lewis, R.E.; Kontoyiannis, D.P. Role and interpretation of antifungal susceptibility testing for the management of invasive fungal infections. J. Fungi 2021, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.Y.; Liang, H.Y.; Jia, J.H. Taxonomic relationships among the taxa in the Candida guilliermondii complex, as revealed by comparative electrophoretic karyotyping. Int. J. Syst. Evol. Microbiol. 2000, 50, 417–422. [Google Scholar] [CrossRef]
- Vaughan-Martini, A.; Kurtzman, C.P.; Meyer, S.A.; O’Neill, E.B. Two new species in the Pichia guilliermondii clade: Pichia caribbica sp. nov., the ascosporic state of Candida fermentati, and Candida carpophila comb. nov. FEMS Yeast Res. 2005, 5, 463–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marco, L.; Epis, S.; Capone, A.; Martin, E.; Bozic, J.; Crotti, E.; Ricci, I.; Sassera, D. The genomes of four Meyerozyma caribbica isolates and novel insights into the Meyerozyma guilliermondii species complex. G3 2018, 8, 755–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, A.; Trilles, L.; Alves, G.M.; Figueiredo-Carvalho, M.; Brito-Santos, F.; Coelho, R.A.; Martins, I.S.; Almeida-Paes, R. A case-series of bloodstream infections caused by the Meyerozyma guilliermondii species complex at a reference center of oncology in Brazil. Med. Mycol. 2021, 4, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Brilhante, R.; Castelo-Branco, D.; Soares, G.; Astete-Medrano, D.J.; Monteiro, A.J.; Cordeiro, R.A.; Sidrim, J.; Rocha, M. Characterization of the gastrointestinal yeast microbiota of cockatiels (Nymphicus hollandicus): A potential hazard to human health. J. Med. Microbiol. 2010, 59, 718–723. [Google Scholar] [CrossRef] [Green Version]
- Brito, E.H.; Fontenelle, R.O.; Brilhante, R.S.; Cordeiro, R.A.; Monteiro, A.J.; Sidrim, J.J.; Rocha, M.F. The anatomical distribution and antimicrobial susceptibility of yeast species isolated from healthy dogs. Vet. J. 2009, 182, 320–326. [Google Scholar] [CrossRef]
- Bliss, J.M. Candida parapsilosis: An emerging pathogen developing its own identity. Virulence 2015, 6, 109–111. [Google Scholar] [CrossRef] [Green Version]
- Wirth, F.; Goldani, L.Z. Epidemiology of Rhodotorula: An emerging pathogen. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 465717. [Google Scholar] [CrossRef]
- Ioannou, P.; Vamvoukaki, R.; Samonis, G. Rhodotorula species infections in humans: A systematic review. Mycoses 2019, 62, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.T.; Kirkeby, L.P.; Poulsen, K.; Reinholdt, J.; Kilian, M. Resident aerobic microbiota of the adult human nasal cavity. APMIS 2000, 108, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Noverr, M.C.; Huffnagle, G.B. Does the microbiota regulate immune outside the gut? Trends Microbiol. 2004, 12, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Schenck, L.P.; McGrath, J.; Lamarche, D.; Stämpfli, M.R.; Bowdish, D.; Surette, M.G. Nasal tissue extraction is essential for characterization of the murine upper respiratory tract microbiota. mSphere 2020, 5, e00562-20. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; van der Heijden, M. Soil microbiomes and one health. Nat. Rev. Microbiol. 2022; 1–15, epub ahead of print. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Isolate ID | GenBank Accession Number | Closest Phylogenetic Neighbor | % Identity | % Query | Sequence Length |
|---|---|---|---|---|---|
| E4.1 | ON706026 | Diutina catenulata CBS 565 | 99 | 100 | 390 |
| E6.2 | ON706027 | Diutina catenulata CBS 565 | 99 | 99 | 397 |
| E6.3 | ON706028 | Cryptococcus aspenensis DS712 | 99.8 | 86 | 512 |
| E7 | ON706029 | Wickerhamomyces anomalus L428/15 | 100 | 100 | 590 |
| F2.1 | ON706030 | Diutina catenulata CBS 565 | 99.5 | 96 | 405 |
| F2.2 | ON706031 | Wickerhamomyces anomalus L428/15 | 100 | 100 | 591 |
| F2.3 | ON706032 | Pichia kluyveri E225 | 99.8 | 97 | 434 |
| F2.4 | ON706033 | Candida parapsilosis LMICRO180 | 100 | 100 | 500 |
| F4.1 | ON706034 | Wickerhamomyces anomalus CBS 113 | 100 | 100 | 590 |
| F4.2 | ON706035 | Diutina catenulata CBS 565 | 99 | 100 | 398 |
| F5.3 | ON706036 | Galactomyces geotrichum SPPRISTMF1 | 98 | 100 | 350 |
| F6 | ON706037 | Candida parapsilosis IFM 63564 | 99.8 | 100 | 490 |
| F9.1 | ON706038 | Wickerhamomyces anomalus CBS:261 | 100 | 100 | 592 |
| G2.1 | ON706039 | Wickerhamomyces anomalus CBS 5759 | 99.8 | 100 | 590 |
| G3.1 | ON706040 | Wickerhamomyces anomalus T12 | 100 | 100 | 422 |
| G5.1 | ON706041 | (Candida) railenensis R97308 | 99.8 | 100 | 600 |
| G6.2 | ON706042 | Wickerhamomyces anomalus 3Y66 | 100 | 100 | 580 |
| G6.3 | ON706043 | (Candida) railenensis R97308 | 99.8 | 100 | 600 |
| G7.2 | ON706044 | Wickerhamomyces anomalus SSL32 | 100 | 100 | 420 |
| G8.1 | ON706045 | Wickerhamomyces anomalus SSL32 | 100 | 100 | 422 |
| G8.2 | ON706046 | Pichia fermentans strain E224 | 100 | 100 | 424 |
| G9.1 | ON706047 | Wickerhamomyces anomalus TTG-100 | 100 | 100 | 425 |
| G9.2 | ON706048 | (Candida) railenensis R97308 | 99.8 | 100 | 602 |
| G9.4 | ON706049 | Pichia kluyveri E225 | 99.8 | 100 | 422 |
| MC.2 | ON706050 | Meyerozyma caribbica E12 + 4 | 99 | 98 | 590 |
| MC.3 | ON706051 | Meyerozyma (Candida) carpophila FF3 | 98.8 | 99 | 590 |
| MC.10 | ON706052 | Meyerozyma caribbica CBS 9966 | 99 | 99 | 595 |
| PCE.3 | ON706054 | Rhodotorula glutinis H1 | 99.5 | 97 | 592 |
| PCA8 | ON706053 | Diutina catenulata CBS 565 | 99.5 | 100 | 372 |
| SP.8 | ON706055 | Meyerozyma caribbica CBS 9966 | 99.8 | 99 | 582 |
| Isolate ID | Molecular Identification | Antifungals | |||
|---|---|---|---|---|---|
| FLU-25 * | ITC-10 | VO-1 | CAS-5 | ||
| ATCC90028 | Candida albicans | 28 | 12 | 29 | 20 |
| E4.1 | Diutina catenulata | 19 | 12 | 21 | 11 |
| E7 | Wickerhamomyces anomalus | 26 | NGI | 18 | 24 |
| F2.1 | Diutina catenulata | 17 | 17 | 31 | NGI |
| F2.2 | Wickerhamomyces anomalus | 26 | NGI | 22 | 23 |
| F2.3 | Pichia kluyveri | NGI | NGI | NGI | 23 |
| F4.1 | Wickerhamomyces anomalus | 22 | 10 | 17 | 22 |
| F4.2 | Diutina catenulata | 15 | 15 | 11 | 27 |
| G2.1 | Wickerhamomyces anomalus | 28 | 12 | 22 | 23 |
| G3.1 | Wickerhamomyces anomalus | 32 | NGI | 21 | 24 |
| G6.2 | Wickerhamomyces anomalus | 26 | NGI | 14 | 25 |
| G7.2 | Wickerhamomyces anomalus | 31 | NGI | 20 | 22 |
| G8.1 | Wickerhamomyces anomalus | 27 | 12 | 30 | 23 |
| G8.2 | Pichia fermentans | NGI | NGI | NGI | 23 |
| G9.1 | Wickerhamomyces anomalus | 29 | NGI | 20 | 22 |
| G9.2 | (Candida) railenensis | 15 | 7 | 10 | 30 |
| G9.4 | Pichia kluyveri | NGI | NGI | NGI | 21 |
| MC10 | Meyerozyma caribbica | 27 | NGI | 39 | NGI |
| MC2 | Meyerozyma caribbica | 30 | NGI | 31 | NGI |
| MC3 | Meyerozyma carpophila | 24 | NGI | 31 | NGI |
| PCA.8 | Diutina catenulata | 16 | 11 | 22 | 10 |
| SP8 | Meyerozyma caribbica | 29 | NGI | 30 | 12 |
| PCE3 | Rhodotorula glutinis | NGI | NGI | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buela, L.; Cuenca, M.; Sarmiento, J.; Peláez, D.; Mendoza, A.Y.; Cabrera, E.J.; Yarzábal, L.A. Role of Guinea Pigs (Cavia porcellus) Raised as Livestock in Ecuadorian Andes as Reservoirs of Zoonotic Yeasts. Animals 2022, 12, 3449. https://doi.org/10.3390/ani12243449
Buela L, Cuenca M, Sarmiento J, Peláez D, Mendoza AY, Cabrera EJ, Yarzábal LA. Role of Guinea Pigs (Cavia porcellus) Raised as Livestock in Ecuadorian Andes as Reservoirs of Zoonotic Yeasts. Animals. 2022; 12(24):3449. https://doi.org/10.3390/ani12243449
Chicago/Turabian StyleBuela, Lenys, Mercy Cuenca, Jéssica Sarmiento, Diana Peláez, Ana Yolanda Mendoza, Erika Judith Cabrera, and Luis Andrés Yarzábal. 2022. "Role of Guinea Pigs (Cavia porcellus) Raised as Livestock in Ecuadorian Andes as Reservoirs of Zoonotic Yeasts" Animals 12, no. 24: 3449. https://doi.org/10.3390/ani12243449
APA StyleBuela, L., Cuenca, M., Sarmiento, J., Peláez, D., Mendoza, A. Y., Cabrera, E. J., & Yarzábal, L. A. (2022). Role of Guinea Pigs (Cavia porcellus) Raised as Livestock in Ecuadorian Andes as Reservoirs of Zoonotic Yeasts. Animals, 12(24), 3449. https://doi.org/10.3390/ani12243449