
Aquaporins and Animal Gamete Cryopreservation: Advances and Future Challenges

 ,
,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Cryopreservation Techniques of Gametes and Embryos
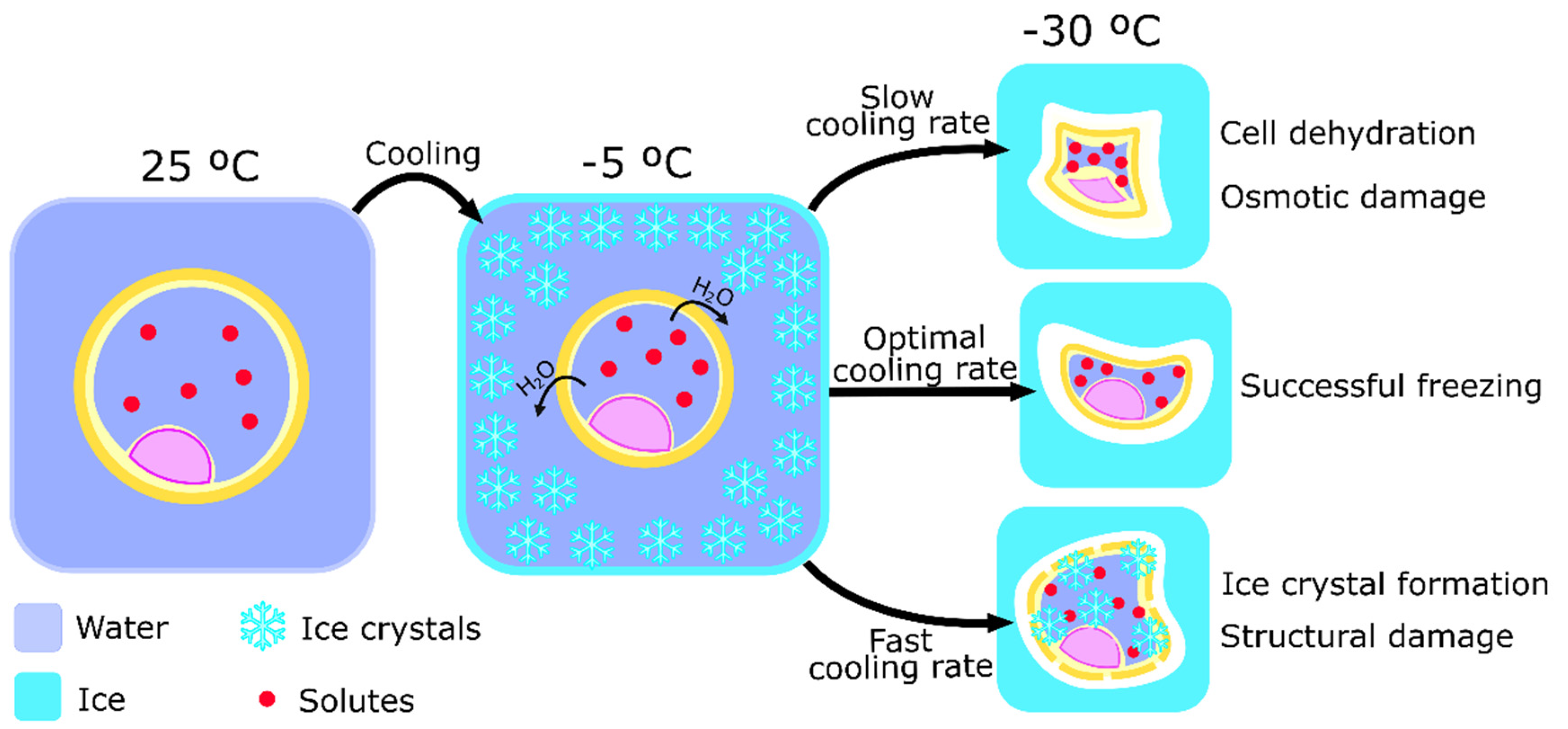
2.1. Cryoinjury
2.2. Cryoprotective Agents (CPAs)
2.3. Controlled-Rate Slow Freezing
2.4. Vitrification
2.5. Controlled-Rated Slow Freezing Versus Vitrification: Advantages and Drawbacks
3. Aquaporins (AQPs) and Cryotolerance: Expression and Function in Spermatozoa, Oocyte, and Embryos
3.1. AQPs Expression and Function in Spermatozoa
AQPs expression and Cryotolerance in Spermatozoa
3.2. AQPs Expression and Function in Oocytes and Embryos
AQPs Expression and Cryotolerance in Oocytes and Embryos
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ugur, M.R.; Saber Abdelrahman, A.; Evans, H.C.; Gilmore, A.A.; Hitit, M.; Arifiantini, R.I.; Purwantara, B.; Kaya, A.; Memili, E. Advances in Cryopreservation of Bull Sperm. Front. Vet. Sci. 2019, 6, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Santo, M.; Tarozzi, N.; Nadalini, M.; Borini, A. Human Sperm Cryopreservation: Update on Techniques, Effect on DNA Integrity, and Implications for ART. Adv. Urol. 2012, 2012, 854837. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.L.; Bilodeau, J.F.; Cormier, N. Semen cryopreservation in domestic animals: A damaging and capacitating phenomenon. J. Androl. 2000, 21, 1–7. [Google Scholar]
- Spallanzani, L.; Bonnet, C. Opuscoli di Fisica Animale e Vegetabile; Presso La Societa’ Tipografica: Modena, Italy, 1776. [Google Scholar]
- Polge, C.; Smith, A.U.; Parkes, A.S. Revival of spermatozoa after vitrification and dehydration at low temperatures. Nature 1949, 164, 666. [Google Scholar] [CrossRef] [PubMed]
- Polge, C.; Rowson, L.E.A. Fertilizing capacity of bull spermatozoa after freezing at −79 °C. Nature 1952, 169, 626–627. [Google Scholar] [CrossRef]
- Bunge, R.G.; Sherman, J.K. Fertilizing capacity of frozen human spermatozoa. Nature 1953, 172, 767–768. [Google Scholar] [CrossRef] [PubMed]
- Sherman, J.K. Synopsis of the Use of Frozen Human Semen Since 1964: State of the Art of Human Semen Banking. Fertil. Steril. 1973, 24, 397–412. [Google Scholar] [CrossRef]
- Perloff, W.H.; Steinberger, E. In Vivo Survival of Spermatozoa in Cervical Mucus. Am. J. Obstet. Gynecol. 1964, 88, 439–442. [Google Scholar] [CrossRef]
- Andrabi, S.M. Factors affecting the quality of cryopreserved buffalo (Bubalus bubalis) bull spermatozoa. Reprod. Domest. Anim. 2009, 44, 552–569. [Google Scholar] [CrossRef]
- Rasul, Z.; Ahmad, N.; Anzar, M. Changes in motion characteristics, plasma membrane integrity, and acrosome morphology during cryopreservation of buffalo spermatozoa. J. Androl. 2001, 22, 278–283. [Google Scholar]
- Paynter, S.J. A rational approach to oocyte cryopreservation. Reprod. Biomed. Online 2005, 10, 578–586. [Google Scholar] [CrossRef]
- Massip, A. Cryopreservation of embryos of farm animals. Reprod. Domest. Anim. 2001, 36, 49–55. [Google Scholar] [CrossRef]
- Jang, T.H.; Park, S.C.; Yang, J.H.; Kim, J.Y.; Seok, J.H.; Park, U.S.; Choi, C.W.; Lee, S.R.; Han, J. Cryopreservation and its clinical applications. Integr. Med. Res. 2017, 6, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.B.; de la Llave, E.; Welke, K.; Scherlis, D.A.; Molinero, V. Freezing, melting and structure of ice in a hydrophilic nanopore. Phys. Chem. Chem. Phys. 2010, 12, 4124–4134. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.B.; Molinero, V. Structural transformation in supercooled water controls the crystallization rate of ice. Nature 2011, 479, 506–508. [Google Scholar] [CrossRef] [PubMed]
- Estudillo, E.; Jimenez, A.; Bustamante-Nieves, P.E.; Palacios-Reyes, C.; Velasco, I.; Lopez-Ornelas, A. Cryopreservation of Gametes and Embryos and Their Molecular Changes. Int. J. Mol. Sci. 2021, 22, 10864. [Google Scholar] [CrossRef] [PubMed]
- McGann, L.E.; Yang, H.Y.; Walterson, M. Manifestations of cell damage after freezing and thawing. Cryobiology 1988, 25, 178–185. [Google Scholar] [CrossRef]
- Liu, X.; Xu, Y.; Liu, F.; Pan, Y.; Miao, L.; Zhu, Q.; Tan, S. The Feasibility of Antioxidants Avoiding Oxidative Damages from Reactive Oxygen Species in Cryopreservation. Front. Chem. 2021, 9, 648684. [Google Scholar] [CrossRef]
- Meryman, H.T. Cryoprotective agents. Cryobiology 1971, 8, 173–183. [Google Scholar] [CrossRef]
- Meryman, H.T.; Williams, R.J.; Douglas, M.S.J. Freezing injury from “solution effects” and its prevention by natural or artificial cryoprotection. Cryobiology 1977, 14, 287–302. [Google Scholar] [CrossRef]
- Lovelock, J.E. The protective action of neutral solutes against haemolysis by freezing and thawing. Biochem. J. 1954, 56, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Tharasanit, T.; Thuwanut, P. Oocyte Cryopreservation in Domestic Animals and Humans: Principles, Techniques and Updated Outcomes. Animals 2021, 11, 2949. [Google Scholar] [CrossRef] [PubMed]
- Len, J.S.; Koh, W.S.D.; Tan, S.X. The roles of reactive oxygen species and antioxidants in cryopreservation. Biosci. Rep. 2019, 39, BSR20191601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadirvel, G.; Kumar, S.; Kumaresan, A. Lipid peroxidation, mitochondrial membrane potential and DNA integrity of spermatozoa in relation to intracellular reactive oxygen species in liquid and frozen-thawed buffalo semen. Anim. Reprod. Sci. 2009, 114, 125–134. [Google Scholar] [CrossRef]
- Banday, M.N.; Lone, F.A.; Rasool, F.; Rashid, M.; Shikari, A. Use of antioxidants reduce lipid peroxidation and improve quality of crossbred rAm. sperm during its cryopreservation. Cryobiology 2017, 74, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Mostek, A.; Dietrich, M.A.; Słowińska, M.; Ciereszko, A. Cryopreservation of bull semen is associated with carbonylation of sperm proteins. Theriogenology 2017, 92, 95–102. [Google Scholar] [CrossRef]
- Ladeira, C.; Koppen, G.; Scavone, F.; Giovannelli, L. The comet assay for human biomonitoring: Effect of cryopreservation on DNA damage in different blood cell preparations. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2019, 843, 11–17. [Google Scholar] [CrossRef]
- Karow, A.M., Jr. Cryoprotectants—A new class of drugs. J. Pharm. Pharmacol. 1969, 21, 209–223. [Google Scholar] [CrossRef]
- Weng, L.; Chen, C.; Zuo, J.; Li, W. Molecular dynamics study of effects of temperature and concentration on hydrogen-bond abilities of ethylene glycol and glycerol: Implications for cryopreservation. J. Phys. Chem. A 2011, 115, 4729–4737. [Google Scholar] [CrossRef]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A review of the actions and applications of cryoprotective solutes that modulate cell recovery from ultra-low temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef]
- Whaley, D.; Damyar, K.; Witek, R.P.; Mendoza, A.; Alexander, M.; Lakey, J.R. Cryopreservation: An Overview of Principles and Cell-Specific Considerations. Cell Transpl. 2021, 30, 963689721999617. [Google Scholar] [CrossRef] [PubMed]
- Crowe, J.H.; Crowe, L.M.; Carpenter, J.F.; Rudolph, A.S.; Wistrom, C.A.; Spargo, B.J.; Anchordoguy, T.J. Interactions of sugars with membranes. Biochim. Biophys. Acta 1988, 947, 367–384. [Google Scholar] [CrossRef]
- Crowe, J.H.; Hoekstra, F.A.; Crowe, L.M.; Anchordoguy, T.J.; Drobnis, E. Lipid phase transitions measured in intact cells with Fourier transform infrared spectroscopy. Cryobiology 1989, 26, 76–84. [Google Scholar] [CrossRef]
- Spindler, R.; Wolkers, W.F.; Glasmacher, B. Effect of Me(2)SO on membrane phase behavior and protein denaturation of human pulmonary endothelial cells studied by in situ FTIR spectroscopy. J. Biomech. Eng. 2009, 131, 074517. [Google Scholar] [CrossRef]
- Oldenhof, H.; Friedel, K.; Akhoondi, M.; Gojowsky, M.; Wolkers, W.F.; Sieme, H. Membrane phase behavior during cooling of stallion sperm and its correlation with freezability. Mol. Membr. Biol. 2012, 29, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Oldenhof, H.; Gojowsky, M.; Wang, S.; Henke, S.; Yu, C.; Rohn, K.; Wolkers, W.F.; Sieme, H. Osmotic stress and membrane phase changes during freezing of stallion sperm: Mode of action of cryoprotective agents. Biol. Reprod. 2013, 88, 68. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Hou, Y.; Zeng, S.; Li, J.; Zhu, S.; Fu, X. The Error-Prone Kinetochore-Microtubule Attachments During Meiosis I in Vitrified Oocytes. Front. Cell Dev. Biol. 2020, 8, 621. [Google Scholar] [CrossRef]
- Namgoong, S.; Kim, N.H. Meiotic spindle formation in mammalian oocytes: Implications for human infertility. Biol. Reprod. 2018, 98, 153–161. [Google Scholar] [CrossRef]
- Johnson, M.H.; Pickering, S.J. The effect of dimethylsulphoxide on the microtubular system of the mouse oocyte. Development 1987, 100, 313–324. [Google Scholar] [CrossRef]
- Van der Elst, J.; Van den Abbeel, E.; Jacobs, R.; Wisse, E.; Van Steirteghem, A. Effect of 1,2-propanediol and dimethylsulphoxide on the meiotic spindle of the mouse oocyte. Hum. Reprod. 1988, 3, 960–967. [Google Scholar] [CrossRef]
- Gook, D.A.; Osborn, S.M.; Johnston, W.I. Cryopreservation of mouse and human oocytes using 1,2-propanediol and the configuration of the meiotic spindle. Hum. Reprod. 1993, 8, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Fuller, B.J. Cryoprotectants: The essential antifreezes to protect life in the frozen state. CryoLetters 2004, 25, 375–388. [Google Scholar] [PubMed]
- Eroglu, A. Cryopreservation of mammalian oocytes by using sugars: Intra- and extracellular raffinose with small amounts of dimethylsulfoxide yields high cryosurvival, fertilization, and development rates. Cryobiology 2010, 60, S54–S59. [Google Scholar] [CrossRef] [Green Version]
- Seet, V.Y.; Al-Samerria, S.; Wong, J.; Stanger, J.; Yovich, J.L.; Almahbobi, G. Optimising vitrification of human oocytes using multiple cryoprotectants and morphological and functional assessment. Reprod. Fertil. Dev. 2013, 25, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Kohaya, N.; Fujiwara, K.; Ito, J.; Kashiwazaki, N. High developmental rates of mouse oocytes cryopreserved by an optimized vitrification protocol: The effects of cryoprotectants, calcium and cumulus cells. J. Reprod. Dev. 2011, 57, 675–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tharasanit, T.; Manee-In, S.; Buarpung, S.; Chatdarong, K.; Lohachit, C.; Techakumphu, M. Successful pregnancy following transfer of feline embryos derived from vitrified immature cat oocytes using ’stepwise’ cryoprotectant exposure technique. Theriogenology 2011, 76, 1442–1449. [Google Scholar] [CrossRef]
- Hughes, Z.E.; Mark, A.E.; Mancera, R.L. Molecular dynamics simulations of the interactions of DMSO with DPPC and DOPC phospholipid membranes. J. Phys. Chem. B 2012, 116, 11911–11923. [Google Scholar] [CrossRef]
- Rudolph, A.S.; Crowe, J.H.; Crowe, L.M. Effects of three stabilizing agents—Proline, betaine, and trehalose—on membrane phospholipids. Arch. Biochem. Biophys. 1986, 245, 134–143. [Google Scholar] [CrossRef]
- Hotamisligil, S.; Toner, M.; Powers, R.D. Changes in membrane integrity, cytoskeletal structure, and developmental potential of murine oocytes after vitrification in ethylene glycol. Biol. Reprod. 1996, 55, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Meryman, H.T.; Hornblower, M. A method for freezing and washing red blood cells using a high glycerol concentration. Transfusion 1972, 12, 145–156. [Google Scholar] [CrossRef]
- Sloviter, H.A. In-Vivo Survival of Rabbit’s Red Cells Recovered after Freezing. Lancet 1951, 257, 1350–1351. [Google Scholar] [CrossRef]
- Oda, K.; Gibbons, W.E.; Leibo, S.P. Osmotic shock of fertilized mouse ova. J. Reprod. Fertil. 1992, 95, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, P.; Kerin, J.F. Experience with the cryopreservation of human embryos using the mouse as a model to establish successful techniques. J. Vitro Fert. Embryo Transf. 1986, 3, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Pope, C.E.; Pope, V.Z.; Beck, L.R. Cryopreservation and transfer of baboon embryos. J. In Vitro Fert. Embryo Transf. 1986, 3, 33–39. [Google Scholar] [CrossRef]
- Fabbri, R.; Porcu, E.; Marsella, T.; Rocchetta, G.; Venturoli, S.; Flamigni, C. Human oocyte cryopreservation: New perspectives regarding oocyte survival. Hum. Reprod. 2001, 16, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, V.; Coticchio, G.; Distratis, V.; Di Giusto, N.; Flamigni, C.; Borini, A. Differential sucrose concentration during dehydration (0.2 mol/l) and rehydration (0.3 mol/l) increases the implantation rate of frozen human oocytes. Reprod. Biomed. Online 2007, 14, 64–71. [Google Scholar] [CrossRef]
- Coticchio, G.; De Santis, L.; Rossi, G.; Borini, A.; Albertini, D.; Scaravelli, G.; Alecci, C.; Bianchi, V.; Nottola, S.; Cecconi, S. Sucrose concentration influences the rate of human oocytes with normal spindle and chromosome configurations after slow-cooling cryopreservation. Hum. Reprod. 2006, 21, 1771–1776. [Google Scholar] [CrossRef] [Green Version]
- Parmegiani, L.; Fabbri, R.; Cognigni, G.E.; Bernardi, S.; Pocognoli, P.; Filicori, M. Blastocyst formation, pregnancy, and birth derived from human oocytes cryopreserved for 5 years. Fertil. Steril. 2008, 90, 2014.e7–2014.e10. [Google Scholar] [CrossRef]
- Whittingham, D.G. Survival of mouse embryos after freezing and thawing. Nature 1971, 233, 125–126. [Google Scholar] [CrossRef]
- Gomez, E.; Carrocera, S.; Martin, D.; Perez-Janez, J.J.; Prendes, J.; Prendes, J.M.; Vazquez, A.; Murillo, A.; Gimeno, I.; Munoz, M. Efficient one-step direct transfer to recipients of thawed bovine embryos cultured in vitro and frozen in chemically defined medium. Theriogenology 2020, 146, 39–47. [Google Scholar] [CrossRef]
- Saragusty, J.; Arav, A. Current progress in oocyte and embryo cryopreservation by slow freezing and vitrification. Reproduction 2011, 141, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Critser, J.K. Mechanisms of cryoinjury in living cells. ILAR J. 2000, 41, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahy, G.M.; MacFarlane, D.R.; Angell, C.A.; Meryman, H.T. Vitrification as an approach to cryopreservation. Cryobiology 1984, 21, 407–426. [Google Scholar] [CrossRef]
- Rall, W.F.; Wood, M.J.; Kirby, C.; Whittingham, D.G. Development of mouse embryos cryopreserved by vitrification. J. Reprod. Fertil. 1987, 80, 499–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wowk, B. Thermodynamic aspects of vitrification. Cryobiology 2010, 60, 11–22. [Google Scholar] [CrossRef]
- Ozkavukcu, S.; Erdemli, E.; Isik, A.; Oztuna, D.; Karahuseyinoglu, S. Effects of cryopreservation on sperm parameters and ultrastructural morphology of human spermatozoa. J. Assist. Reprod. Genet. 2008, 25, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.; Rasappan, P.; Shila, S.; Gunasekaran, K. A simple method of human sperm vitrification. MethodsX 2019, 6, 2198–2204. [Google Scholar] [CrossRef]
- Tao, Y.; Sanger, E.; Saewu, A.; Leveille, M.C. Human sperm vitrification: The state of the art. Reprod. Biol. Endocrinol. 2020, 18, 17. [Google Scholar] [CrossRef]
- Wurth, Y.A.; Reinders, J.M.C.; Rall, W.F.; Kruip, T.H.A.M. Developmental potential of in vitro produced bovine embryos following cryopreservation and single-embryo transfer. Theriogenology 1994, 42, 1275–1284. [Google Scholar] [CrossRef]
- Agca, Y.; Monson, R.L.; Northey, D.L.; Mazni, O.A.; Schaefer, D.M.; Rutledge, J.J. Transfer of fresh and cryopreserved IVP bovine embryos: Normal calving, birth weight and gestation lengths. Theriogenology 1998, 50, 147–162. [Google Scholar] [CrossRef]
- Sanches, B.V.; Lunardelli, P.A.; Tannura, J.H.; Cardoso, B.L.; Pereira, M.H.; Gaitkoski, D.; Basso, A.C.; Arnold, D.R.; Seneda, M.M. A new direct transfer protocol for cryopreserved IVF embryos. Theriogenology 2016, 85, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Araújo-Lemos, P.F.; Freitas Neto, L.M.; Moura, M.T.; Melo, J.V.; Lima, P.F.; Oliveira, M.A. Comparison of vitrification and conventional freezing for cryopreservation of caprine embryos. Zygote 2015, 23, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.H.; Sharma, V.; Khan, F.A.; Naykoo, N.A.; Yaqoob, S.H.; Vajta, G.; Khan, H.M.; Fazili, M.R.; Ganai, N.A.; Shah, R.A. Open pulled straw vitrification and slow freezing of sheep IVF embryos using different cryoprotectants. Reprod. Fertil. Dev. 2015, 27, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos-Neto, P.C.; Cuadro, F.; Barrera, N.; Crispo, M.; Menchaca, A. Embryo survival and birth rate after minimum volume vitrification or slow freezing of in vivo and in vitro produced ovine embryos. Cryobiology 2017, 78, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, W.K.; Roelen, B.A.; Colenbrander, B.; Stout, T.A. Cellular damage suffered by equine embryos after exposure to cryoprotectants or cryopreservation by slow-freezing or vitrification. Equine Vet. J. 2015, 47, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, D.; Panzani, D.; Rota, A.; Tesi, M.; Camillo, F.; Bollwein, H.; Herrera, C. Cryopreservation of donkey embryos: Comparison of embryo survival rate after in vitro culture between conventional freezing and vitrification. Theriogenology 2020, 154, 11–16. [Google Scholar] [CrossRef]
- Naik, B.R.; Rao, B.S.; Vagdevi, R.; Gnanprakash, M.; Amarnath, D.; Rao, V.H. Conventional slow freezing, vitrification and open pulled straw (OPS) vitrification of rabbit embryos. Anim. Reprod. Sci. 2005, 86, 329–338. [Google Scholar] [CrossRef]
- Mokrousova, V.I.; Okotrub, K.A.; Brusentsev, E.Y.; Kizilova, E.A.; Surovtsev, N.V.; Amstislavsky, S.Y. Effects of slow freezing and vitrification on embryo development in domestic cat. Reprod. Domest. Anim. 2020, 55, 1328–1336. [Google Scholar] [CrossRef]
- Uechi, H.; Tsutsumi, O.; Morita, Y.; Takai, Y.; Taketani, Y. Comparison of the effects of controlled-rate cryopreservation and vitrification on 2-cell mouse embryos and their subsequent development. Hum. Reprod. 1999, 14, 2827–2832. [Google Scholar] [CrossRef] [Green Version]
- Zander-Fox, D.; Lane, M.; Hamilton, H. Slow freezing and vitrification of mouse morula and early blastocysts. J. Assist. Reprod. Genet. 2013, 30, 1091–1098. [Google Scholar] [CrossRef] [Green Version]
- Meikle, M.N.; Schlapp, G.; Menchaca, A.; Crispo, M. Minimum volume Spatula MVD vitrification method improves embryo survival compared to traditional slow freezing, both for in vivo and in vitro produced mice embryos. Cryobiology 2018, 84, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Song, Y.; Cheng, K.M.; Silversides, F.G. Production of donor-derived offspring from cryopreserved ovarian tissue in Japanese quail (Coturnix japonica). Biol. Reprod. 2010, 83, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Marinovic, Z.; Li, Q.; Lujic, J.; Iwasaki, Y.; Csenki, Z.; Urbanyi, B.; Yoshizaki, G.; Horvath, A. Preservation of zebrafish genetic resources through testis cryopreservation and spermatogonia transplantation. Sci. Rep. 2019, 9, 13861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauvigne, F.; Boj, M.; Vilella, S.; Finn, R.N.; Cerda, J. Subcellular localization of selectively permeable aquaporins in the male germ line of a marine teleost reveals spatial redistribution in activated spermatozoa. Biol. Reprod. 2013, 89, 37. [Google Scholar] [CrossRef] [Green Version]
- Loeffler, C.A.; LØVtrup, S. Water Balance in the Salmon Egg. J. Exp. Biol. 1970, 52, 291–298. [Google Scholar] [CrossRef]
- Marques, L.S.; Fossati, A.A.N.; Rodrigues, R.B.; Da Rosa, H.T.; Izaguirry, A.P.; Ramalho, J.B.; Moreira, J.C.F.; Santos, F.W.; Zhang, T.; Streit, D.P., Jr. Slow freezing versus vitrification for the cryopreservation of zebrafish (Danio rerio) ovarian tissue. Sci. Rep. 2019, 9, 15353. [Google Scholar] [CrossRef]
- Cavusoglu, T.; Popken, J.; Guengoer, T.; Yilmaz, O.; Uyanikgil, Y.; Ates, U.; Baka, M.; Oztas, E.; Zakhartchenko, V. Ultra-Structural Alterations in In Vitro Produced Four-Cell Bovine Embryos Following Controlled Slow Freezing or Vitrification. Anat. Histol. Embryol. 2016, 45, 291–307. [Google Scholar] [CrossRef]
- Mathias, F.J.; D’Souza, F.; Uppangala, S.; Salian, S.R.; Kalthur, G.; Adiga, S.K. Ovarian tissue vitrification is more efficient than slow freezing in protecting oocyte and granulosa cell DNA integrity. Syst Biol Reprod Med 2014, 60, 317–322. [Google Scholar] [CrossRef]
- Li, L.; Zhang, X.; Zhao, L.; Xia, X.; Wang, W. Comparison of DNA apoptosis in mouse and human blastocysts after vitrification and slow freezing. Mol. Reprod. Dev. 2012, 79, 229–236. [Google Scholar] [CrossRef]
- Brair, V.L.; Maia, A.; Correia, L.F.L.; Barbosa, N.O.; Santos, J.D.R.; Brandão, F.Z.; Fonseca, J.F.; Batista, R.; Souza-Fabjan, J.M.G. Gene expression patterns of in vivo-derived sheep blastocysts is more affected by vitrification than slow freezing technique. Cryobiology 2020, 95, 110–115. [Google Scholar] [CrossRef]
- Pradiee, J.; Esteso, M.C.; Castaño, C.; Toledano-Díaz, A.; Lopez-Sebastián, A.; Guerra, R.; Santiago-Moreno, J. Conventional slow freezing cryopreserves mouflon spermatozoa better than vitrification. Andrologia 2017, 49. [Google Scholar] [CrossRef] [PubMed]
- Daramola, J.O.; Adekunle, E.O. Comparative effects of slow freezing and vitrification on cryosurvival of spermatozoa obtained from west African dwarf goat bucks. CryoLetters 2016, 37, 123–128. [Google Scholar] [PubMed]
- Salazar, J.L., Jr.; Teague, S.R.; Love, C.C.; Brinsko, S.P.; Blanchard, T.L.; Varner, D.D. Effect of cryopreservation protocol on postthaw characteristics of stallion sperm. Theriogenology 2011, 76, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Demyda-Peyrás, S.; Bottrel, M.; Acha, D.; Ortiz, I.; Hidalgo, M.; Carrasco, J.J.; Gómez-Arrones, V.; Gósalvez, J.; Dorado, J. Effect of cooling rate on sperm quality of cryopreserved Andalusian donkey spermatozoa. Anim. Reprod. Sci. 2018, 193, 201–208. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, F.M.; Ramsey, C.; Hanna, C.B.; do Valle, R.D.R.; Nichi, M.; Binelli, M.; Guimarães, M.; Hennebold, J.D. Cryopreservation and Preparation of Thawed Spermatozoa from Rhesus Macaques (Macaca mulatta) for In Vitro Fertilization. J. Am. Assoc. Lab. Anim. Sci. 2021, 60, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Mphaphathi, M.L.; Luseba, D.; Sutherland, B.; Nedambale, T.L. Comparison of slow freezing and vitrification methods for Venda cockerel’s spermatozoa. Open J. Anim. Sci. 2012, 02, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.C.; Alves, M.G.; Yeste, M.; Cho, Y.S.; Calamita, G.; Oliveira, P.F. Aquaporins and (in)fertility: More than just water transport. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2021, 1867, 166039. [Google Scholar] [CrossRef]
- Agre, P. Aquaporin water channels (Nobel Lecture). Angew. Chem. Int. Ed. Engl. 2004, 43, 4278–4290. [Google Scholar] [CrossRef]
- Sales, A.D.; Lobo, C.H.; Carvalho, A.A.; Moura, A.A.; Rodrigues, A.P. Structure, function, and localization of aquaporins: Their possible implications on gamete cryopreservation. Genet. Mol. Res. 2013, 12, 6718–6732. [Google Scholar] [CrossRef]
- Ishibashi, K.; Tanaka, Y.; Morishita, Y. The role of mammalian superaquaporins inside the cell. Biochim. Biophys. Acta 2014, 1840, 1507–1512. [Google Scholar] [CrossRef]
- Soria, L.R.; Fanelli, E.; Altamura, N.; Svelto, M.; Marinelli, R.A.; Calamita, G. Aquaporin-8-facilitated mitochondrial ammonia transport. Biochem. Biophys. Res. Commun. 2010, 393, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Carrageta, D.F.; Bernardino, R.L.; Soveral, G.; Calamita, G.; Alves, M.G.; Oliveira, P.F. Aquaporins and male (in)fertility: Expression and role throughout the male reproductive tract. Arch. Biochem. Biophys. 2020, 679, 108222. [Google Scholar] [CrossRef] [PubMed]
- Morató, R.; Prieto-Martínez, N.; Muiño, R.; Hidalgo, C.O.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Aquaporin 11 is related to cryotolerance and fertilising ability of frozen-thawed bull spermatozoa. Reprod. Fertil. Dev. 2018, 30, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Martínez, N.; Vilagran, I.; Morató, R.; Rivera Del Álamo, M.M.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Relationship of aquaporins 3 (AQP3), 7 (AQP7), and 11 (AQP11) with boar sperm resilience to withstand freeze-thawing procedures. Andrology 2017, 5, 1153–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, J.; Kawabe, M.; Ochiai, H.; Suzukamo, C.; Harada, M.; Mitsugi, Y.; Seita, Y.; Kashiwazaki, N. Expression and immunodetection of aquaporin 1 (AQP1) in canine spermatozoa. Cryobiology 2008, 57, 312–314. [Google Scholar] [CrossRef]
- Offenberg, H.; Barcroft, L.C.; Caveney, A.; Viuff, D.; Thomsen, P.D.; Watson, A.J. mRNAs encoding aquaporins are present during murine preimplantation development. Mol. Reprod. Dev. 2000, 57, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Boj, M.; Chauvigné, F.; Cerdà, J. Aquaporin biology of spermatogenesis and sperm physiology in mammals and teleosts. Biol. Bull. 2015, 229, 93–108. [Google Scholar] [CrossRef]
- Cerda, J.; Zapater, C.; Chauvigne, F.; Finn, R.N. Water homeostasis in the fish oocyte: New insights into the role and molecular regulation of a teleost-specific aquaporin. Fish Physiol. Biochem. 2013, 39, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Peng, H.; Lei, L.; Zhang, Y.; Kuang, H.; Cao, Y.; Shi, Q.X.; Ma, T.; Duan, E. Aquaporin3 is a sperm water channel essential for postcopulatory sperm osmoadaptation and migration. Cell Res. 2011, 21, 922–933. [Google Scholar] [CrossRef]
- Prieto-Martínez, N.; Morató, R.; Muiño, R.; Hidalgo, C.O.; Rodríguez-Gil, J.E.; Bonet, S.; Yeste, M. Aquaglyceroporins 3 and 7 in bull spermatozoa: Identification, localisation and their relationship with sperm cryotolerance. Reprod. Fertil. Dev. 2017, 29, 1249–1259. [Google Scholar] [CrossRef]
- Edashige, K.; Sakamoto, M.; Kasai, M. Expression of mRNAs of the aquaporin family in mouse oocytes and embryos. Cryobiology 2000, 40, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Paynter, S.J.; Cooper, A.; Gregory, L.; Fuller, B.J.; Shaw, R.W. Permeability characteristics of human oocytes in the presence of the cryoprotectant dimethylsulphoxide. Hum. Reprod. 1999, 14, 2338–2342. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.W.; Jee, B.C.; Suh, C.S.; Kim, S.H.; Choi, Y.M.; Kim, J.G.; Moon, S.Y. Effect of maturation on the expression of aquaporin 3 in mouse oocyte. Zygote 2011, 19, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.J.; Zhang, X.Y.; Ding, G.L.; Li, R.; Wang, L.; Jin, L.; Lin, X.H.; Gao, L.; Sheng, J.Z.; Huang, H.F. Aquaporin7 plays a crucial role in tolerance to hyperosmotic stress and in the survival of oocytes during cryopreservation. Sci. Rep. 2015, 5, 17741. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Tan, Y.J.; Xiong, Y.M.; Huang, Y.T.; Hu, X.L.; Lu, Y.C.; Ye, Y.H.; Wang, T.T.; Zhang, D.; Jin, F.; et al. Expression of aquaporins in human embryos and potential role of AQP3 and AQP7 in preimplantation mouse embryo development. Cell Physiol. Biochem. 2013, 31, 649–658. [Google Scholar] [CrossRef]
- Barcroft, L.C.; Offenberg, H.; Thomsen, P.; Watson, A.J. Aquaporin proteins in murine trophectoderm mediate transepithelial water movements during cavitation. Dev. Biol. 2003, 256, 342–354. [Google Scholar] [CrossRef] [Green Version]
- Hermo, L.; Schellenberg, M.; Liu, L.Y.; Dayanandan, B.; Zhang, T.; Mandato, C.A.; Smith, C.E. Membrane domain specificity in the spatial distribution of aquaporins 5, 7, 9, and 11 in efferent ducts and epididymis of rats. J. Histochem. Cytochem. 2008, 56, 1121–1135. [Google Scholar] [CrossRef] [Green Version]
- Kasimanickam, R.K.; Kasimanickam, V.R.; Arangasamy, A.; Kastelic, J.P. Associations of hypoosmotic swelling test, relative sperm volume shift, aquaporin7 mRNA abundance and bull fertility estimates. Theriogenology 2017, 89, 162–168. [Google Scholar] [CrossRef]
- Skowronski, M.T.; Lebeck, J.; Rojek, A.; Praetorius, J.; Fuchtbauer, E.M.; Frokiaer, J.; Nielsen, S. AQP7 is localized in capillaries of adipose tissue, cardiac and striated muscle: Implications in glycerol metabolism. Am. J. Physiol. Renal Physiol. 2007, 292, F956–F965. [Google Scholar] [CrossRef]
- Vicente-Carrillo, A.; Ekwall, H.; Alvarez-Rodriguez, M.; Rodriguez-Martinez, H. Membrane Stress During Thawing Elicits Redistribution of Aquaporin 7 But Not of Aquaporin 9 in Boar Spermatozoa. Reprod. Domest. Anim. 2016, 51, 665–679. [Google Scholar] [CrossRef] [Green Version]
- Bonilla-Correal, S.; Noto, F.; Garcia-Bonavila, E.; Rodriguez-Gil, J.E.; Yeste, M.; Miro, J. First evidence for the presence of aquaporins in stallion sperm. Reprod. Domest. Anim. 2017, 52 (Suppl. 4), 61–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronski, M.T.; Leska, A.; Robak, A.; Nielsen, S. Immunolocalization of aquaporin-1, -5, and -7 in the avian testis and vas deferens. J. Histochem. Cytochem. 2009, 57, 915–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, C.H.; Callies, C.; Rojek, A.; Nielsen, S.; Cooper, T.G. Aquaporin isoforms involved in physiological volume regulation of murine spermatozoa. Biol. Reprod. 2009, 80, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Santiago, C.; Yeste, M.; Rigau, T.; Rodríguez-Gil, J.; Rivera del Alamo, M. Canine sperm cells express aquaporin-8, but not aquaporin-2. Reprod. Domest. Anim. 2019, 54, 63–74. [Google Scholar] [CrossRef]
- Hamatani, T.; Carter, M.G.; Sharov, A.A.; Ko, M.S. Dynamics of global gene expression changes during mouse preimplantation development. Dev. Cell 2004, 6, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Chauvigne, F.; Boj, M.; Finn, R.N.; Cerda, J. Mitochondrial aquaporin-8-mediated hydrogen peroxide transport is essential for teleost spermatozoon motility. Sci. Rep. 2015, 5, 7789. [Google Scholar] [CrossRef] [Green Version]
- Ford, P.; Merot, J.; Jawerbaum, A.; Gimeno, M.A.; Capurro, C.; Parisi, M. Water permeability in rat oocytes at different maturity stages: Aquaporin-9 expression. J. Membr. Biol. 2000, 176, 151–158. [Google Scholar] [CrossRef]
- Zilli, L.; Schiavone, R.; Chauvigné, F.; Cerdà, J.; Storelli, C.; Vilella, S. Evidence for the involvement of aquaporins in sperm motility activation of the teleost gilthead sea bream (Sparus aurata). Biol. Reprod. 2009, 81, 880–888. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Martínez, N.; Vilagran, I.; Morató, R.; Rodríguez-Gil, J.E.; Yeste, M.; Bonet, S. Aquaporins 7 and 11 in boar spermatozoa: Detection, localisation and relationship with sperm quality. Reprod. Fertil. Dev. 2016, 28, 663–672. [Google Scholar] [CrossRef] [Green Version]
- Yeung, C.H.; Cooper, T.G. Aquaporin AQP11 in the testis: Molecular identity and association with the processing of residual cytoplasm of elongated spermatids. Reproduction 2010, 139, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Offenberg, H.; Thomsen, P.D. Functional challenge affects aquaporin mRNA abundance in mouse blastocysts. Mol. Reprod. Dev. 2005, 71, 422–430. [Google Scholar] [CrossRef]
- Thirumala, S.; Ferrer, M.S.; Al-Jarrah, A.; Eilts, B.E.; Paccamonti, D.L.; Devireddy, R.V. Cryopreservation of canine spermatozoa: Theoretical prediction of optimal cooling rates in the presence and absence of cryoprotective agents. Cryobiology 2003, 47, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Alavi, S.M.; Cosson, J. Sperm motility in fishes. (II) Effects of ions and osmolality: A review. Cell Biol. Int. 2006, 30, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sohara, E.; Ueda, O.; Tachibe, T.; Hani, T.; Jishage, K.; Rai, T.; Sasaki, S.; Uchida, S. Morphologic and functional analysis of sperm and testes in Aquaporin 7 knockout mice. Fertil. Steril. 2007, 87, 671–676. [Google Scholar] [CrossRef]
- Mann, T.; White, I.G. Metabolism of glycerol, sorbitol and related compounds by spermatozoa. Nature 1956, 178, 142–143. [Google Scholar] [CrossRef] [PubMed]
- Suzuki-Toyota, F.; Ishibashi, K.; Yuasa, S. Immunohistochemical localization of a water channel, aquaporin 7 (AQP7), in the rat testis. Cell Tissue Res. 1999, 295, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Calamita, G.; Mazzone, A.; Cho, Y.S.; Valenti, G.; Svelto, M. Expression and localization of the aquaporin-8 water channel in rat testis. Biol. Reprod. 2001, 64, 1660–1666. [Google Scholar] [CrossRef] [Green Version]
- Yeung, C.H. Aquaporins in spermatozoa and testicular germ cells: Identification and potential role. Asian J. Androl. 2010, 12, 490–499. [Google Scholar] [CrossRef] [Green Version]
- Chauvigne, F.; Ducat, C.; Ferre, A.; Hansen, T.; Carrascal, M.; Abian, J.; Finn, R.N.; Cerda, J. A multiplier peroxiporin signal transduction pathway powers piscine spermatozoa. Proc. Natl. Acad. Sci. USA 2021, 118, e2019346118. [Google Scholar] [CrossRef]
- Laforenza, U.; Pellavio, G.; Marchetti, A.L.; Omes, C.; Todaro, F.; Gastaldi, G. Aquaporin-Mediated Water and Hydrogen Peroxide Transport Is Involved in Normal Human Spermatozoa Functioning. Int. J. Mol. Sci. 2016, 18, 66. [Google Scholar] [CrossRef] [Green Version]
- Yeste, M.; Morató, R.; Rodríguez-Gil, J.E.; Bonet, S.; Prieto-Martínez, N. Aquaporins in the male reproductive tract and sperm: Functional implications and cryobiology. Reprod. Domest. Anim. 2017, 52 (Suppl. 4), 12–27. [Google Scholar] [CrossRef] [Green Version]
- Fujii, T.; Hirayama, H.; Fukuda, S.; Kageyama, S.; Naito, A.; Yoshino, H.; Moriyasu, S.; Yamazaki, T.; Sakamoto, K.; Hayakawa, H.; et al. Expression and localization of aquaporins 3 and 7 in bull spermatozoa and their relevance to sperm motility after cryopreservation. J. Reprod. Dev. 2018, 64, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Bermudez, A.; Llavanera, M.; Fernandez-Bastit, L.; Recuero, S.; Mateo-Otero, Y.; Bonet, S.; Barranco, I.; Fernandez-Fuertes, B.; Yeste, M. Aquaglyceroporins but not orthodox aquaporins are involved in the cryotolerance of pig spermatozoa. J. Anim. Sci. Biotechnol. 2019, 10, 77. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Bermudez, A.; Noto, F.; Bonilla-Correal, S.; Garcia-Bonavila, E.; Catalan, J.; Papas, M.; Bonet, S.; Miro, J.; Yeste, M. Cryotolerance of Stallion Spermatozoa Relies on Aquaglyceroporins rather than Orthodox Aquaporins. Biology 2019, 8, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Bermudez, A.; Llavanera, M.; Recuero, S.; Mateo-Otero, Y.; Bonet, S.; Barranco, I.; Fernandez-Fuertes, B.; Yeste, M. Effect of AQP Inhibition on Boar Sperm Cryotolerance Depends on the Intrinsic Freezability of the Ejaculate. Int. J. Mol. Sci. 2019, 20, 6255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calamita, G.; Ferri, D.; Gena, P.; Liquori, G.E.; Cavalier, A.; Thomas, D.; Svelto, M. The inner mitochondrial membrane has aquaporin-8 water channels and is highly permeable to water. J. Biol. Chem. 2005, 280, 17149–17153. [Google Scholar] [CrossRef] [Green Version]
- Madsen, S.S.; Engelund, M.B.; Cutler, C.P. Water transport and functional dynamics of aquaporins in osmoregulatory organs of fishes. Biol. Bull 2015, 229, 70–92. [Google Scholar] [CrossRef]
- Cerda, J.; Chauvigne, F.; Finn, R.N. The Physiological Role and Regulation of Aquaporins in Teleost Germ Cells. Adv. Exp. Med. Biol. 2017, 969, 149–171. [Google Scholar] [CrossRef]
- Kagawa, H.; Kishi, T.; Gen, K.; Kazeto, Y.; Tosaka, R.; Matsubara, H.; Matsubara, T.; Sawaguchi, S. Expression and localization of aquaporin 1b during oocyte development in the Japanese eel (Anguilla japonica). Reprod. Biol. Endocrinol. 2011, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Fabra, M.; Raldua, D.; Bozzo, M.G.; Deen, P.M.; Lubzens, E.; Cerda, J. Yolk proteolysis and aquaporin-1o play essential roles to regulate fish oocyte hydration during meiosis resumption. Dev. Biol. 2006, 295, 250–262. [Google Scholar] [CrossRef]
- Zapater, C.; Chauvigne, F.; Norberg, B.; Finn, R.N.; Cerda, J. Dual neofunctionalization of a rapidly evolving aquaporin-1 paralog resulted in constrained and relaxed traits controlling channel function during meiosis resumption in teleosts. Mol. Biol. Evol. 2011, 28, 3151–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabra, M.; Raldua, D.; Power, D.M.; Deen, P.M.; Cerda, J. Marine fish egg hydration is aquaporin-mediated. Science 2005, 307, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagawa, H.; Horiuchi, Y.; Kasuga, Y.; Kishi, T. Oocyte hydration in the Japanese eel (Anguilla japonica) during meiosis resumption and ovulation. J. Exp. Zool. A Ecol. Genet. Physiol. 2009, 311, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Tingaud-Sequeira, A.; Chauvigne, F.; Fabra, M.; Lozano, J.; Raldua, D.; Cerda, J. Structural and functional divergence of two fish aquaporin-1 water channels following teleost-specific gene duplication. BMC Evol. Biol. 2008, 8, 259. [Google Scholar] [CrossRef] [Green Version]
- Chaube, R.; Chauvigne, F.; Tingaud-Sequeira, A.; Joy, K.P.; Acharjee, A.; Singh, V.; Cerda, J. Molecular and functional characterization of catfish (Heteropneustes fossilis) aquaporin-1b: Changes in expression during ovarian development and hormone-induced follicular maturation. Gen. Comp. Endocrinol. 2011, 170, 162–171. [Google Scholar] [CrossRef]
- Singh, V.; Joy, K.P. An involvement of vasotocin in oocyte hydration in the catfish Heteropneustes fossilis: A comparison with effects of isotocin and hCG. Gen. Comp. Endocrinol. 2010, 166, 504–512. [Google Scholar] [CrossRef]
- Assou, S.; Boumela, I.; Haouzi, D.; Anahory, T.; Dechaud, H.; De Vos, J.; Hamamah, S. Dynamic changes in gene expression during human early embryo development: From fundamental aspects to clinical applications. Hum. Reprod. Update 2011, 17, 272–290. [Google Scholar] [CrossRef] [Green Version]
- Edashige, K. Permeability of the plasma membrane to water and cryoprotectants in mammalian oocytes and embryos: Its relevance to vitrification. Reprod. Med. Biol. 2017, 16, 36–39. [Google Scholar] [CrossRef]
- Edashige, K.; Tanaka, M.; Ichimaru, N.; Ota, S.; Yazawa, K.; Higashino, Y.; Sakamoto, M.; Yamaji, Y.; Kuwano, T.; Valdez, D.M., Jr.; et al. Channel-dependent permeation of water and glycerol in mouse morulae. Biol. Reprod. 2006, 74, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Edashige, K.; Ohta, S.; Tanaka, M.; Kuwano, T.; Valdez, D.M., Jr.; Hara, T.; Jin, B.; Takahashi, S.; Seki, S.; Koshimoto, C.; et al. The role of aquaporin 3 in the movement of water and cryoprotectants in mouse morulae. Biol. Reprod. 2007, 77, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Jin, B.; Higashiyama, R.; Nakata, Y.; Yonezawa, J.; Xu, S.; Miyake, M.; Takahashi, S.; Kikuchi, K.; Yazawa, K.; Mizobuchi, S.; et al. Rapid movement of water and cryoprotectants in pig expanded blastocysts via channel processes: Its relevance to their higher tolerance to cryopreservation. Biol. Reprod. 2013, 89, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, B.; Kawai, Y.; Hara, T.; Takeda, S.; Seki, S.; Nakata, Y.; Matsukawa, K.; Koshimoto, C.; Kasai, M.; Edashige, K. Pathway for the movement of water and cryoprotectants in bovine oocytes and embryos. Biol. Reprod. 2011, 85, 834–847. [Google Scholar] [CrossRef] [PubMed]
- Edashige, K.; Yamaji, Y.; Kleinhans, F.W.; Kasai, M. Artificial expression of aquaporin-3 improves the survival of mouse oocytes after cryopreservation. Biol. Reprod. 2003, 68, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morato, R.; Chauvigne, F.; Novo, S.; Bonet, S.; Cerda, J. Enhanced water and cryoprotectant permeability of porcine oocytes after artificial expression of human and zebrafish aquaporin-3 channels. Mol. Reprod. Dev. 2014, 81, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Bedford-Guaus, S.J.; Chauvigne, F.; Mejia-Ramirez, E.; Marti, M.; Ventura-Rubio, A.; Raya, A.; Cerda, J.; Veiga, A. Expression of the T85A mutant of zebrafish aquaporin 3b improves post-thaw survival of cryopreserved early mammalian embryos. Zygote 2016, 24, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Valdez, D.M., Jr.; Hara, T.; Miyamoto, A.; Seki, S.; Jin, B.; Kasai, M.; Edashige, K. Expression of aquaporin-3 improves the permeability to water and cryoprotectants of immature oocytes in the medaka (Oryzias latipes). Cryobiology 2006, 53, 160–168. [Google Scholar] [CrossRef]
- Chauvigne, F.; Lubzens, E.; Cerda, J. Design and characterization of genetically engineered zebrafish aquaporin-3 mutants highly permeable to the cryoprotectant ethylene glycol. BMC Biotechnol. 2011, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Lande, M.B.; Donovan, J.M.; Zeidel, M.L. The relationship between membrane fluidity and permeabilities to water, solutes, ammonia, and protons. J. Gen. Physiol. 1995, 106, 67–84. [Google Scholar] [CrossRef]
- Giraud, M.N.; Motta, C.; Boucher, D.; Grizard, G. Membrane fluidity predicts the outcome of cryopreservation of human spermatozoa. Hum. Reprod. 2000, 15, 2160–2164. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| AQP Isoform | Spermatozoa | |||||
|---|---|---|---|---|---|---|
| Head | Midpiece | Tail | Oocytes | Embryos | References | |
| AQP1 | Dog * | Dog * | Dog * | - | Mouse # | [106,107] |
| AQP1aa | - | - | Seabream | - | - | [85,108] |
| AQP1ab | Seabream | - | - | Teleost fishes | - | [85,108,109] |
| AQP3 | Pig Stallion * | Pig | Mouse | Mouse | Mouse | [105,110,111,112,113,114,115,116,117] |
| Mouse | Pig | |||||
| Bull | Stallion * | |||||
| Stallion* | ||||||
| AQP5 | - | - | - | - | Mouse # | [107] |
| AQP6 | - | - | - | - | Mouse # | [107] |
| AQP7 | Bull | Rat | Rat | Mouse | Mouse | [85,112,113,115,116,118,119,120,121,122,123] |
| Mouse | ||||||
| Seabream | Bull | Pig | ||||
| Bull | ||||||
| Stallion * | Stallion * | Geese | ||||
| Stallion * | ||||||
| AQP8 | Mouse * Dog * | Mouse * Dog * | Mouse * Dog * | - | Mouse | [112,117,124,125,126] |
| AQP8b | Seabream | - | - | - | - | [127] |
| AQP9 | Pig | - | - | Mouse Rat # | Mouse | [112,115,117,121,128] |
| AQP10b | Seabream | Seabream | Seabream | - | - | [129] |
| AQP11 | Pig | Pig Stallion * | Pig | - | Mouse # | [104,122,130,131,132] |
| Bull | Bull | |||||
| Stallion * | Rat | |||||
| Stallion * | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, J.C.; Carrageta, D.F.; Bernardino, R.L.; Alves, M.G.; Oliveira, P.F. Aquaporins and Animal Gamete Cryopreservation: Advances and Future Challenges. Animals 2022, 12, 359. https://doi.org/10.3390/ani12030359
Ribeiro JC, Carrageta DF, Bernardino RL, Alves MG, Oliveira PF. Aquaporins and Animal Gamete Cryopreservation: Advances and Future Challenges. Animals. 2022; 12(3):359. https://doi.org/10.3390/ani12030359
Chicago/Turabian StyleRibeiro, João C., David F. Carrageta, Raquel L. Bernardino, Marco G. Alves, and Pedro F. Oliveira. 2022. "Aquaporins and Animal Gamete Cryopreservation: Advances and Future Challenges" Animals 12, no. 3: 359. https://doi.org/10.3390/ani12030359
APA StyleRibeiro, J. C., Carrageta, D. F., Bernardino, R. L., Alves, M. G., & Oliveira, P. F. (2022). Aquaporins and Animal Gamete Cryopreservation: Advances and Future Challenges. Animals, 12(3), 359. https://doi.org/10.3390/ani12030359