
The Effect of Environmental Enrichment on Laboratory Rare Minnows (Gobiocypris rarus): Growth, Physiology, and Behavior


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
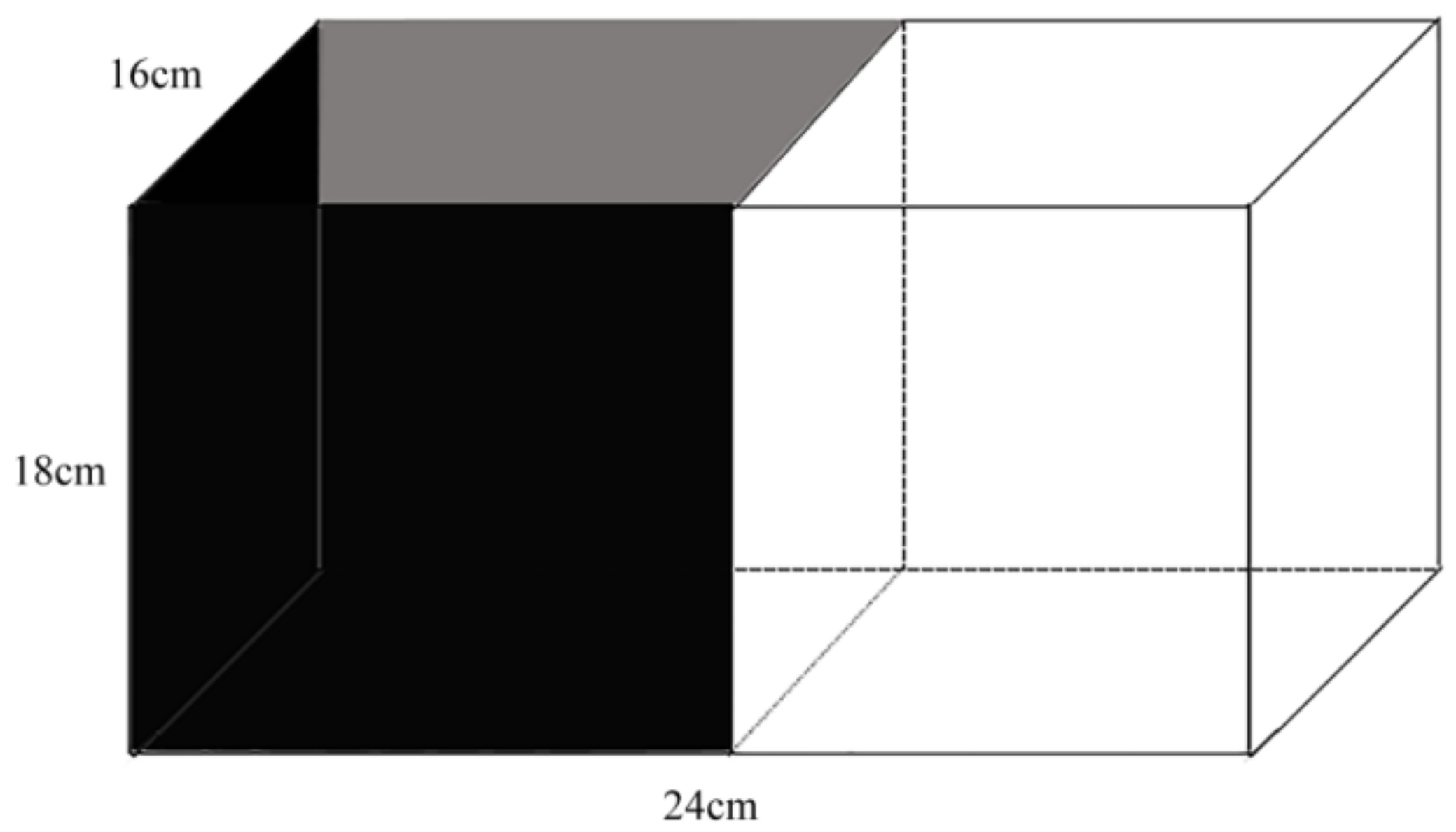
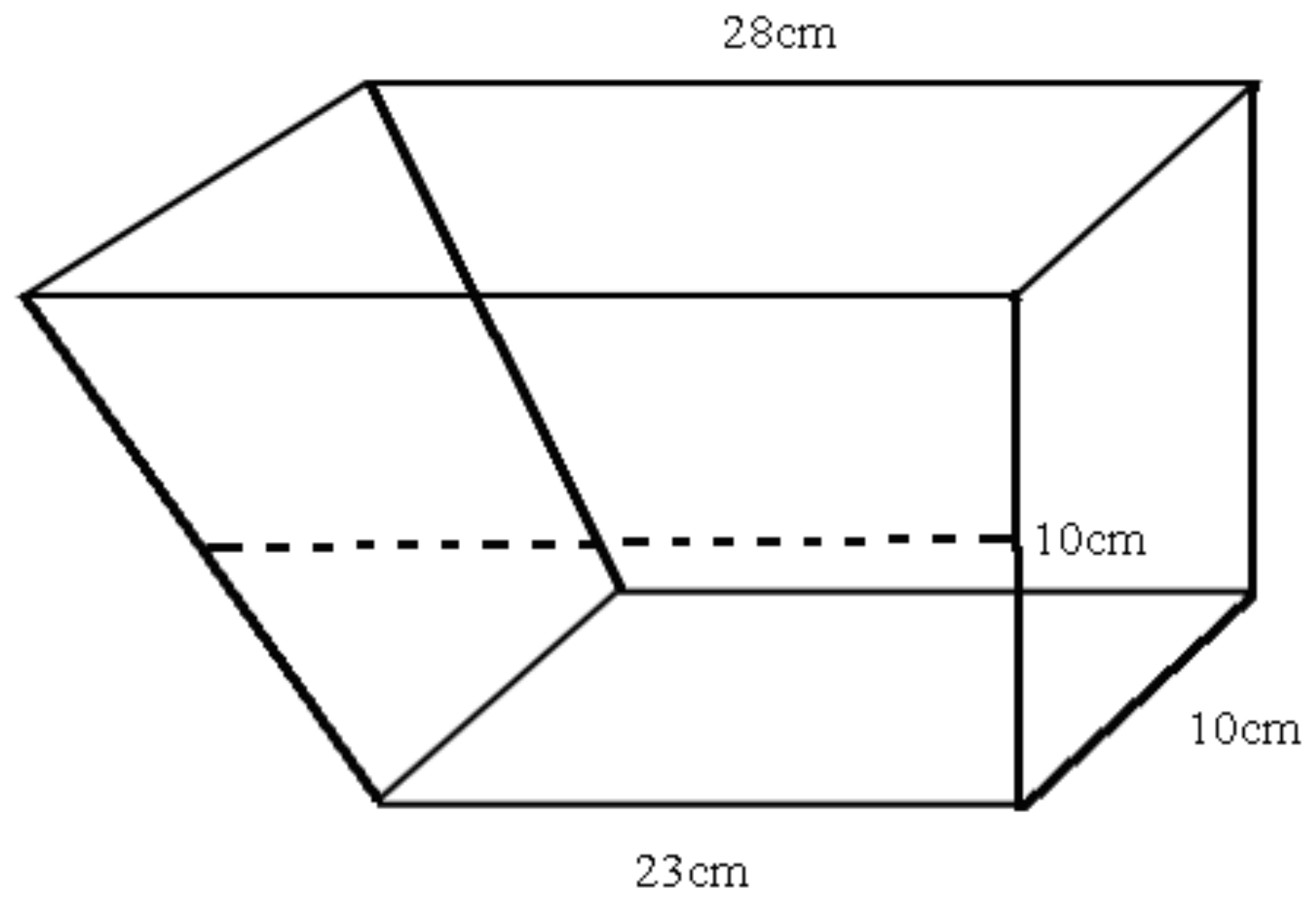
2.1. Fish Culturing and Environmental Enrichment
2.2. Sampling and Measuring of Physiological Parameters
2.3. Anxiety-like Behavioral Studies
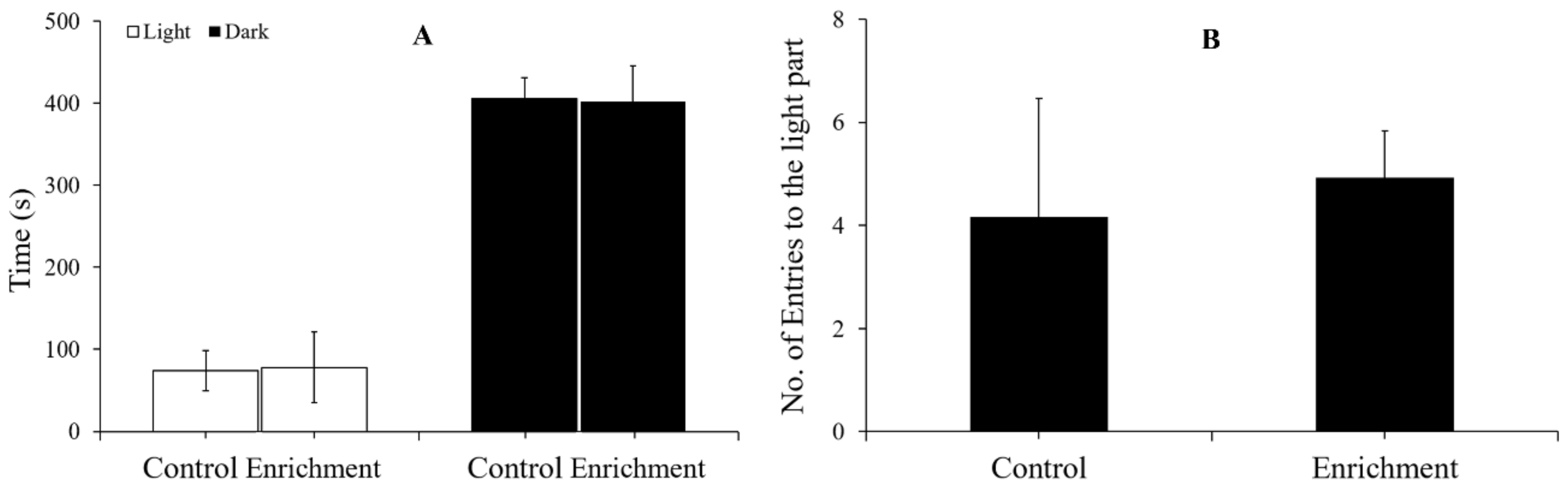
2.3.1. Light-Dark Test
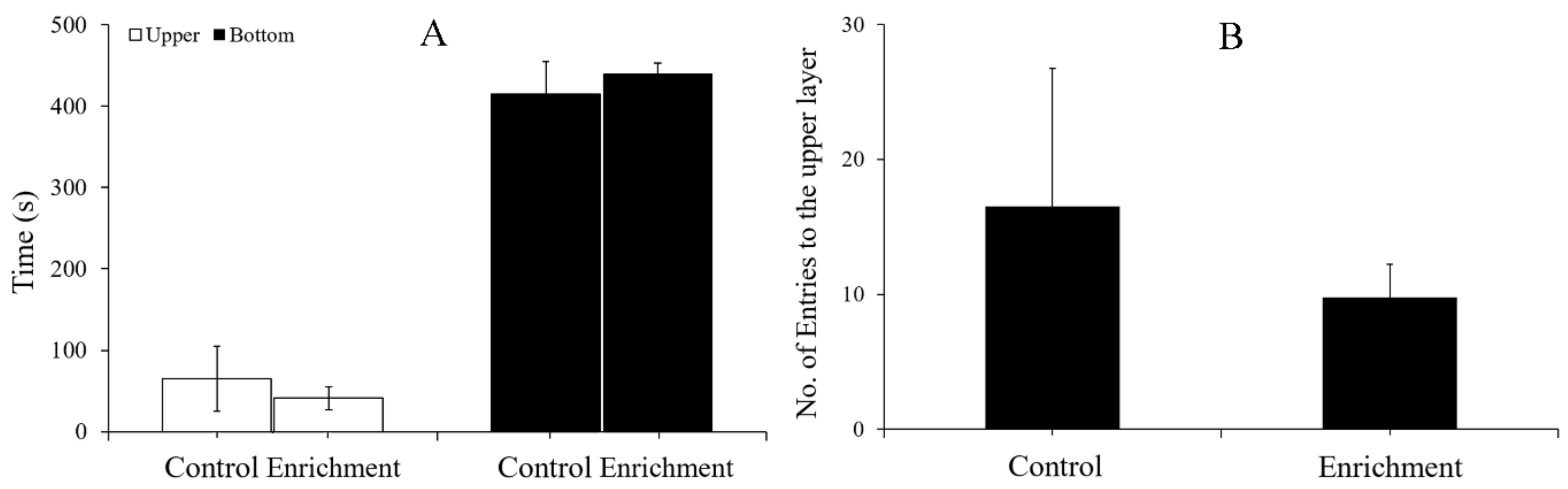
2.3.2. Novel Tank Test
2.4. Data Analysis
3. Results
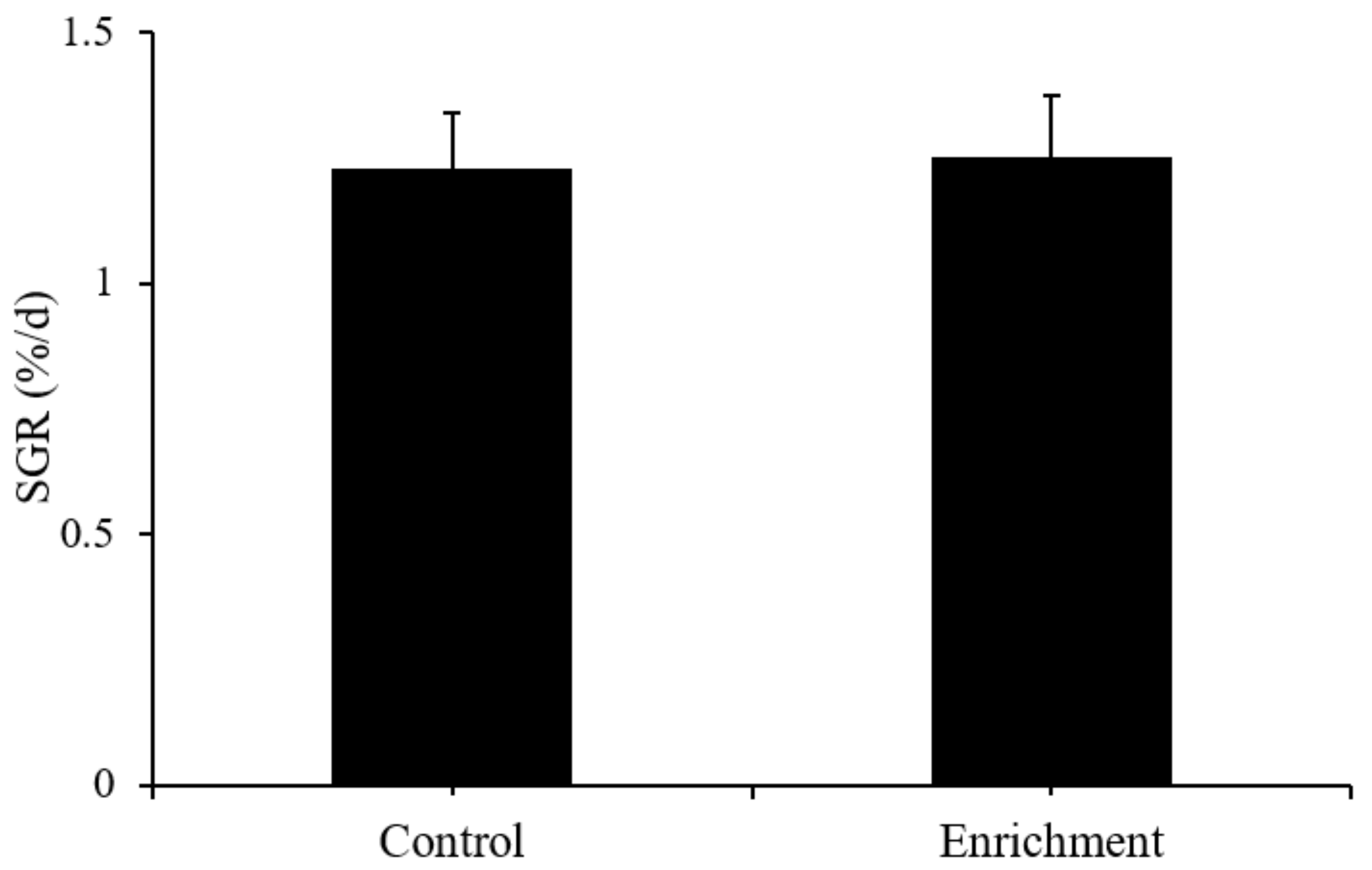
3.1. Growth
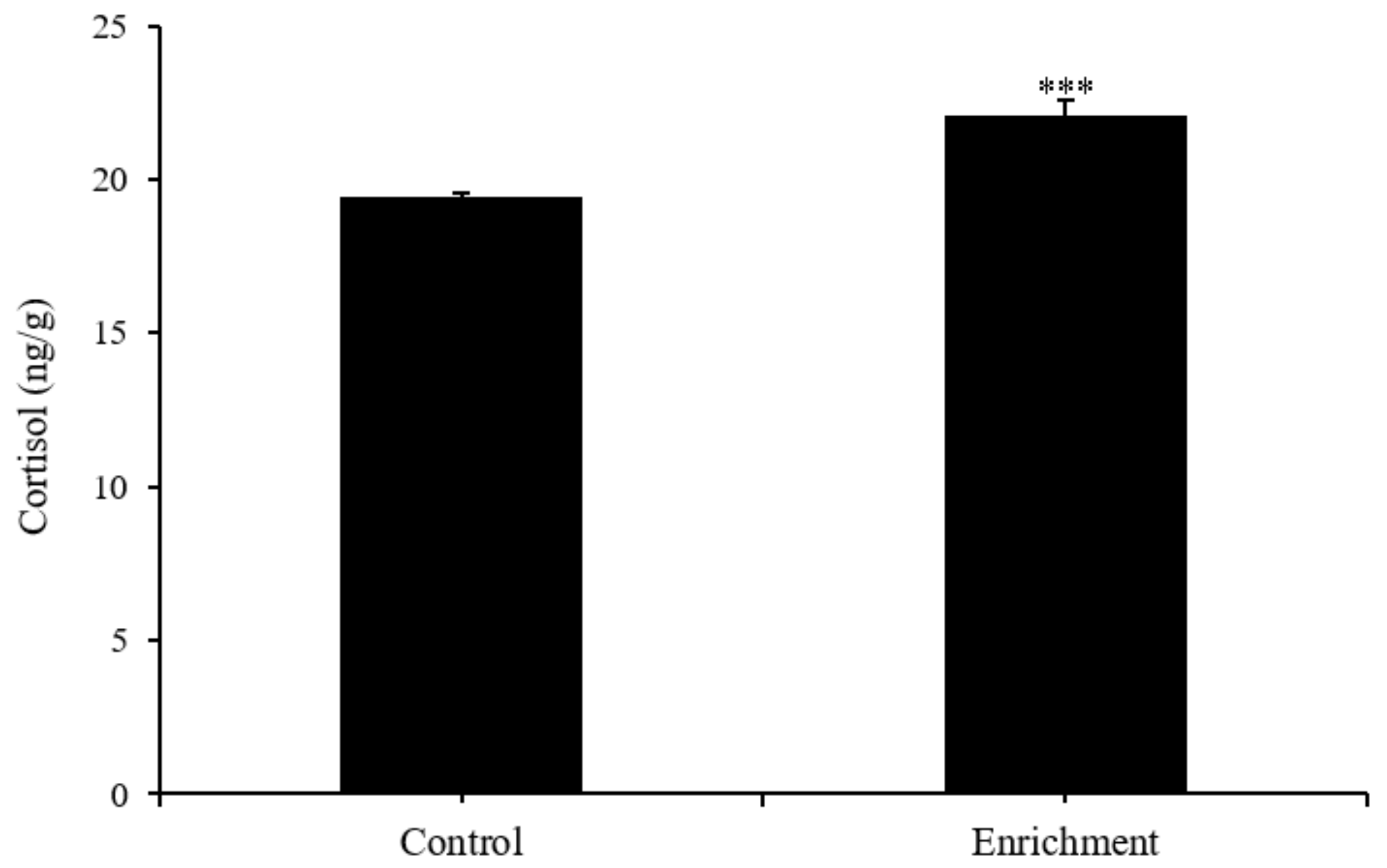
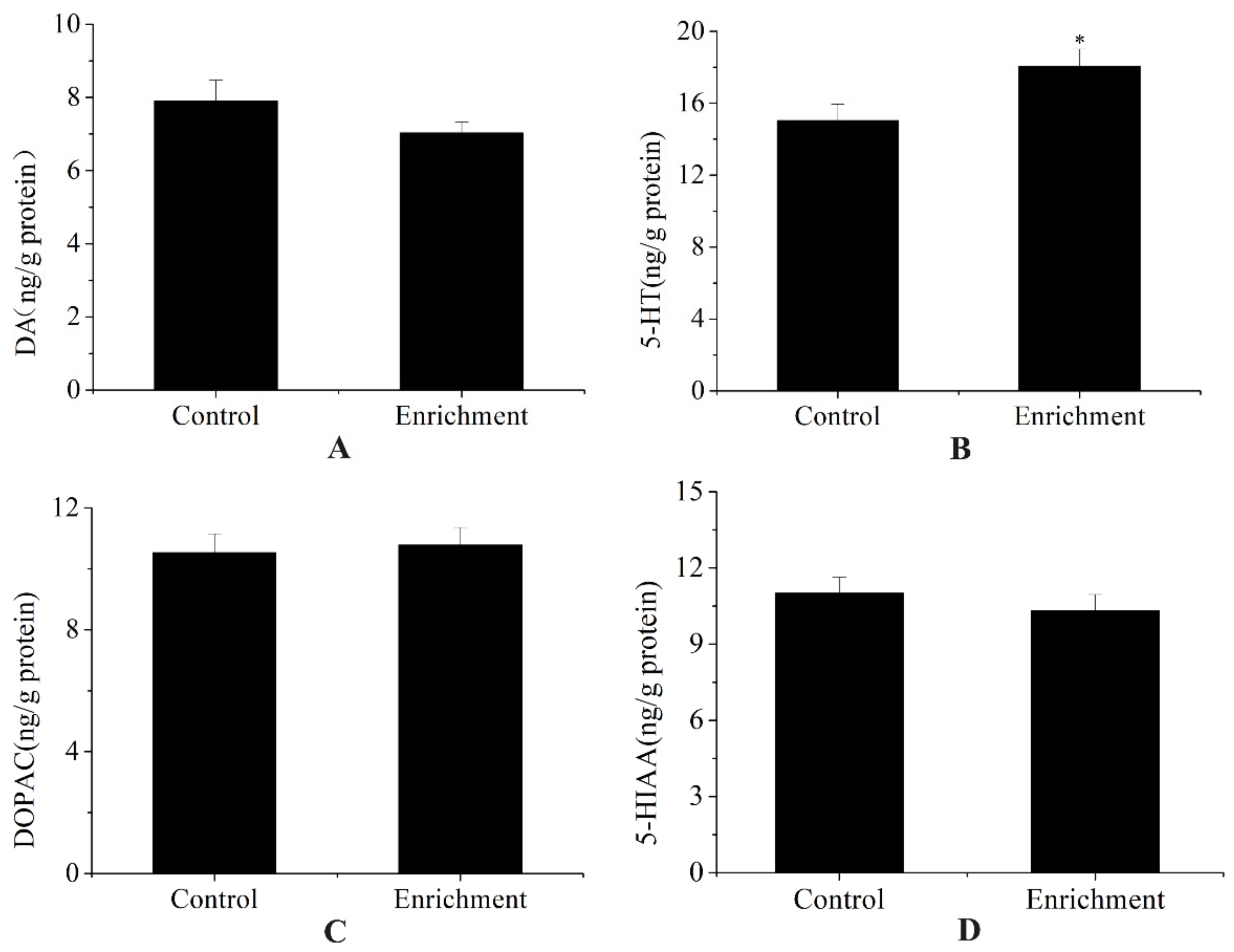
3.2. Physiological Parameters
3.3. Anxiety-like Behavior
4. Discussion
4.1. Effect of Enrichment on Growth
4.2. Effect of Enrichment on Physiological Parameters
4.3. Effect of Enrichment on Anxiety-like Behavior
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Jones, N.; Webster, M.; Salvanes, A. Physical enrichment research for captive fish: Time to focus on the DETAILS. J. Fish Biol. 2021, 99, 704–725. [Google Scholar] [CrossRef] [PubMed]
- Kientz, J.L.; Barnes, M.E. Structural complexity improves the rearing performance of rainbow trout in circular tanks. N. Am. J. Aquacult. 2016, 78, 203–207. [Google Scholar] [CrossRef]
- Kientz, J.L.; Crank, K.M.; Barnes, M.E. Enrichment of circular tanks with vertically suspended strings of colored balls improves Rainbow Trout rearing performance. N. Am. J. Aquacult. 2018, 80, 162–167. [Google Scholar] [CrossRef]
- Crank, K.M.; Kientz, J.L.; Barnes, M.E. An evaluation of vertically suspended environmental enrichment structures during Rainbow Trout rearing. N. Am. J. Aquacult. 2019, 81, 94–100. [Google Scholar] [CrossRef] [Green Version]
- White, S.C.; Krebs, E.; Huysman, N.; Voorhees, J.M.; Barnes, M.E. Use of suspended plastic conduit arrays during Brown Trout and Rainbow Trout rearing in circular tanks. N. Am. J. Aquacult. 2019, 81, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Voorhees, J.M.; Huysman, N.; Krebs, E.; Barnes, M.E. Influence of water velocity and vertically-suspended structures on rainbow trout rearing performance. Open J. Anim. Sci. 2020, 10, 152–161. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.H.; Xu, X.W.; Wang, Y.H.; Zhang, X.M. Effects of environmental enrichment on growth performance, aggressive behavior and stress-induced changes in cortisol release and neurogenesis of black rockfish Sebastes schlegelii. Aquaculture 2020, 528, 735483. [Google Scholar] [CrossRef]
- Batzina, A.; Karakatsouli, N. The presence of substrate as a means of environmental enrichment in intensively reared gilthead seabream Sparus aurata: Growth and behavioral effects. Aquaculture 2012, 370, 54–60. [Google Scholar] [CrossRef]
- Maximino, C.; de Brito, T.M.; Dias, C.A.G.D.; Gouveia, A.; Morato, S. Scototaxis as anxiety-like behavior in fish. Nat. Protoc. 2010, 5, 209–216. [Google Scholar] [CrossRef]
- Salvanes, A.G.V.; Moberg, O.; Ebbesson, L.O.E.; Nilsen, T.O.; Jensen, K.H.; Braithwaite, V.A. Environmental enrichment promotes neural plasticity and cognitive ability in fish. Proc. R. Soc. B 2013, 280, 20131331. [Google Scholar] [CrossRef] [PubMed]
- Roy, T.; Bhat, A. Learning and memory in juvenile zebrafish: What makes the difference—Population or rearing environment? Ethology 2016, 122, 308–318. [Google Scholar] [CrossRef]
- Strand, D.A.; Utne-Palm, A.C.; Jakobsen, P.J.; Braithwaite, V.A.; Jensen, K.H.; Salvanes, A.G.V. Enrichment promotes learning in fish. Mar. Ecol. Prog. Ser. 2010, 412, 273–282. [Google Scholar] [CrossRef]
- Bergendahl, I.A.; Miller, S.; Depasquale, C.; Giralico, L.; Braithwaite, V.A. Becoming a better swimmer: Structural complexity enhances agility in a captive-reared fish. J. Fish Biol. 2017, 90, 1112–1117. [Google Scholar] [CrossRef] [PubMed]
- von Krogh, K.; Sorensen, C.; Nilsson, G.E.; Overli, O. Forebrain cell proliferation, behavior, and physiology of zebrafish, Danio rerio, kept in enriched or barren environments. Physiol. Behav. 2010, 101, 32–39. [Google Scholar] [CrossRef] [PubMed]
- White, S.C.; Barnes, M.E.; Krebs, E.; Huysman, N.; Voorhees, J. Addition of vertical enrichment structures does not improve growth of three salmonid species during hatchery rearing. J. Mar. Biol. Aquacult. 2018, 4, 48–52. [Google Scholar]
- Brydges, N.M.; Braithwaite, V.A. Does environmental enrichment affect the behavior of fish commonly used in laboratory work? Appl. Anim. Behav. Sci. 2009, 118, 137–143. [Google Scholar] [CrossRef]
- Näslund, J.; Rosengren, M.; Johnsson, J.I. Fish density, but not environmental enrichment, affects the size of cerebellum in the brain of juvenile hatchery-reared Atlantic salmon. Environ. Biol. Fishes 2019, 102, 705–712. [Google Scholar] [CrossRef] [Green Version]
- Toli, E.A.; Noreikiene, K.; DeFaveri, J.; Merila, J. Environmental enrichment, sexual dimorphism, and brain size in sticklebacks. Ecol. Evol. 2017, 7, 1691–1698. [Google Scholar] [CrossRef]
- Woodward, M.A.; Winder, L.A.; Watt, P.J. Enrichment increases aggression in zebrafish. Fishes 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, L.; Owen, S.F.; Readman, G.D.; Sloman, K.A.; Wilson, R.W. Does structural enrichment for toxicology studies improve zebrafish welfare? Appl. Anim. Behav. Sci. 2012, 139, 143–150. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Bai, Q.Q.; Xu, X.W.; Guo, H.Y.; Zhang, X.M. Effects of environmental enrichment on the welfare of juvenile black rockfish Sebastes schlegelii: Growth, behavior and physiology. Aquaculture 2020, 518, 734782. [Google Scholar] [CrossRef]
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Spence, R.; Magurran, A.E.; Smith, C. Spatial cognition in zebrafish: The role of strain and rearing environment. Anim. Cogn. 2011, 4, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.; Readman, G.; Owen, S.F. Key issues concerning environmental enrichment for laboratory-held fish species. Lab. Anim. 2009, 43, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Killen, S.S.; Marras, S.; Metcalfe, N.B.; McKenzie, D.J.; Domenici, P. Environmental stressors alter relationships between physiology and behaviour. Trends Ecol. Evol. 2013, 28, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Pounder, K.C.; Mitchell, J.L.; Thomson, J.S.; Pottinger, T.G.; Buckley, J.; Sneddon, L.U. Does environmental enrichment promote recovery from stress in rainbow trout? Appl. Anim. Behav. Sci. 2016, 176, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Su, L.X.; Xu, C.S.; Cai, L.; Qiu, N.; Hou, M.M.; Wang, J.W. Susceptibility and immune responses after challenge with Flavobacterium columnare and Pseudomonas fluorescens in conventional and specific pathogen-free rare minnow (Gobiocypris rarus). Fish Shellfish Immunol. 2020, 98, 875–886. [Google Scholar] [CrossRef]
- Wang, J.J.; Li, P.; Zhang, Y.G.; Peng, Z.G. The complete mitochondrial genome of Chinese rare minnow, Gobiocypris rarus (Teleostei: Cypriniformes). Mitochondrial DNA 2011, 22, 178–180. [Google Scholar] [CrossRef]
- Hua, J.H.; Han, J.; Guo, Y.Y.; Zhou, B.S. Endocrine disruption in Chinese rare minnow (Gobiocypris rarus) after long-term exposure to low environmental concentrations of progestin megestrol Check for acetate. Ecotoxicol. Environ. Saf. 2018, 163, 289–297. [Google Scholar] [CrossRef]
- Lin, Y.S.; Wang, B.; Wang, N.H.; Ouyang, G.; Cao, H. Transcriptome analysis of rare minnow (Gobiocypris rarus) infected by the grass carp reovirus. Fish Shellfish Immunol. 2019, 89, 337–344. [Google Scholar] [CrossRef]
- Hong, X.S.; Zha, J.M. Fish behavior: A promising model for aquatic toxicology research. Sci. Total Environ. 2019, 686, 311–321. [Google Scholar] [CrossRef]
- Zhang, J.L.; Zhang, C.N.; Sun, P.; Shao, X. Tributyltin affects shoaling and anxiety behavior in female rare minnow (Gobiocypris rarus). Aquat. Toxicol. 2016, 178, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Qiu, N.; Xu, C.S.; Wang, X.Z.; Hou, M.M.; Xia, Z.J.; Wang, J.W. Chemicals Weaken Shoal Preference in the Rare Minnow Gobiocypris rarus. Environ. Toxicol. Chem. 2020, 39, 2018–2027. [Google Scholar] [CrossRef]
- Qiu, N. Effect of Chemicals Exposure on Behaviors of Rare Minnow. Ph.D. Thesis, University of Chinese Academy of Sciences, Beijing, China, 2020. [Google Scholar]
- Song, C.; Liu, B.P.; Zhang, Y.P.; Peng, Z.L.; Wang, J.J.; Collier, A.D.; Echevarria, D.J.; Savelieva, K.V.; Lawrence, R.F. Modeling consequences of prolonged strong unpredictable stress in zebrafish: Complex effects on behavior and physiology. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 81, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Collymore, C.; Tolwani, R.J.; Rasmussen, S. The Behavioral Effects of Single Housing and Environmental Enrichment on Adult Zebrafish (Danio rerio). J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 280–285. [Google Scholar] [PubMed]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamilselvan, P.; Sloman, K.A. Developmental social experience of parents affects behaviour of offspring in zebrafish. Anim. Behav. 2017, 133, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Huntingford, F.A.; Adams, C.; Braithwaite, V.A.; Kadri, S.; Pottinger, T.G.; Sandoe, P.; Turnbull, J.F. Current issues in fish welfare. J. Fish Biol. 2006, 68, 332–372. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.L.; Luo, S.; Xie, S.Q.; Wang, J.W. Growth of rare minnows (Gobiocypris rarus) fed different amounts of dietary protein and lipids. Lab Anim. 2016, 45, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.L.; Xiong, X.Q.; Xie, S.Q.; Wang, J.W. Dietary lipid and gross energy affect protein utilization in the rare minnow Gobiocypris rarus. Chin. J. Oceanol. Limnol. 2016, 34, 740–748. [Google Scholar] [CrossRef]
- Wu, B.L.; Luo, S.; Wang, J.W. Effects of temperature and feeding frequency on ingestion and growth for rare minnow. Physiol. Behav. 2015, 140, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Su, L.X.; Luo, S.; Qiu, N.; Xu, C.S.; Hou, M.M.; Xiong, X.Q.; Wang, J.W. Comparative allometric growth of rare minnow (Gobiocypris rarus) in two culture environments. Hydrobiologia 2020, 847, 2083–2095. [Google Scholar] [CrossRef]
- Luo, S.; Wu, B.L.; Xiong, X.Q.; Wang, J.W. Effects of Total Hardness and Calcium: Magnesium Ratio of Water during Early Stages of Rare Minnows (Gobiocypris rarus). Comp. Med. 2016, 66, 181–187. [Google Scholar] [PubMed]
- Selye, H. Stress and the general adaptation syndrome. Br. Med. J. 1950, 1, 1383–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, B.A.; Iwama, G.K. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroid. Annual. Rev. Fish Dis. 1991, 1, 3–26. [Google Scholar] [CrossRef]
- Oliveira, R.; Galhardo, L. Psychological stress and welfare in fish. Annu. Rev. Biomed. Sci. 2009, 11, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fisher. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Sørensen, C.; Johansen, I.B.; Øverli, Ø. Neural plasticity and stress coping in teleost fishes. Gen. Comp. Endocrinol. 2013, 181, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Batzina, A.; Dalla, C.; Papadopoulou-Daifoti, Z.; Karakatsouli, N. Effects of environmental enrichment on growth, aggressive behaviour and brain monoamines of gilthead seabream Sparus aurata reared under different social conditions. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 169, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Batzina, A.; Dalla, C.; Tsopelakos, A.; Papadopoulou-Daifoti, Z.; Karakatsouli, N. Environmental enrichment induces changes in brain monoamine levels in gilthead seabream Sparus aurata. Physiol. Behav. 2014, 130, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Batzina, A.; Kalogiannis, D.; Dalla, C.; Papadopoulou-Daifoti, Z.; Chadio, S.; Karakatsouli, N. Blue substrate modifies the time course of stress response in gilthead seabream Sparus aurata. Aquaculture 2014, 420, 247–253. [Google Scholar] [CrossRef]
- Rosengren, M.; Kvingedal, E.; Naslund, J.; Johnsson, J.I.; Sundell, K. Born to be wild: Effects of rearing density and environmental enrichment on stress, welfare, and smolt migration in hatchery-reared Atlantic salmon. Can. J. Fish. Aquat. Sci. 2016, 74, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Cogliati, K.M.; Herron, C.L.; Noakes, D.L.G.; Schreck, C.B. Reduced stress response in juvenile Chinook Salmon reared with structure. Aquaculture 2019, 504, 96–101. [Google Scholar] [CrossRef]
- Naslund, J.; Rosengren, M.; Del Villar, D.; Gansel, L.; Norrgard, J.R.; Persson, L.; Winkowski, J.J.; Kvingedal, E. Hatchery tank enrichment affects cortisol levels and shelter-seeking in Atlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci. 2013, 70, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Keck, V.A.; Edgerton, D.S.; Hajizadeh, S.; Swift, L.L.; Dupont, W.D.; Lawrence, C.; Boyd, K.L. Effects of habitat complexity on pair-housed zebrafish. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 378–383. [Google Scholar]
- Dickens, M.J.; Romero, L.M.A. Consensus endocrine profile for chronically stressed wild animals does not exist. Gen. Comp. Endocrinol. 2013, 191, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F. Hedonic valence, dopamine and motivation. Mol. Psychiatry 1996, 1, 186–189. [Google Scholar] [PubMed]
- Phillips, A.G.; Vacca, G.; Ahn, S.A. Top-down perspective on dopamine, motivation and memory. Pharmacol. Biochem. Behav. 2008, 90, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Beninger, R.J. The role of dopamine in locomotor activity and learning. Brain Res. Rev. 1983, 6, 173–196. [Google Scholar] [CrossRef] [Green Version]
- Salamone, J.D. Complex motor and sensorimotor functions of striatal and accumbens dopamine: Involvement in instrumental behavior processes. Psychopharmacology 1992, 107, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Saudou, F.; Amara, D.A.; Dierich, A.; Lemeur, M.; Ramboz, S.; Segu, L.; Buhot, M.C.; Hen, R. Enhanced aggressive-behavior in mice lacking 5-HT1B receptor. Science 1994, 265, 1875–1878. [Google Scholar] [CrossRef] [PubMed]
- Mann, J.J. Role of the serotonergic system in the pathogenesis of major depression and suicidal behavior. Neuropsychopharmacology 1999, 21, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Pang, T.Y.C.; Du, X.; Zajac, M.S.; Howard, M.L.; Hannan, A.J. Altered serotonin receptor expression is associated with depression-related behavior in the R6/1 transgenic mouse model of Huntington’s disease. Hum. Mol. Genet. 2009, 18, 753–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, N.E.; Markou, A. Animal models and treatments for addiction and depression co-morbidity. Neurotoxic. Res. 2007, 11, 1–32. [Google Scholar] [CrossRef]
- Mann, J.J. Violence and aggression. In Psychopharmacology: The Fourth Generation of Progress; Bloom, F.E., Kupfer, D.J., Eds.; Raven Press: New York, NY, USA, 1995; pp. 1919–1928. [Google Scholar]
- Benaroya-Milshtein, N.; Hollander, N.; Apter, A.; Kukulansky, T.; Raz, N.; Wilf, A.; Yaniv, I.; Pick, C.G. Environmental enrichment in mice decreases anxiety, attenuates stress responses and enhances natural killer cell activity. Eur. J. Neurosci. 2004, 20, 1341–1347. [Google Scholar] [CrossRef]
- Gortz, N.; Lewejohann, L.; Tomm, M.; Ambree, O.; Keyvani, K.; Paulus, W.; Sachser, N. Effects of environmental enrichment on exploration, anxiety, and memory in female TgCRND8 Alzheimer mice. Behav. Brain Res. 2008, 191, 43–48. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SGR (%/d) | Cortisol (ng/g) | DA (ng/g Protein) | DOPAC (ng/g Protein) | 5-HT (ng/g Protein) | 5-HIAA (ng/g Protein) | |
|---|---|---|---|---|---|---|
| Control | 1.13 ± 0.11 | 19.46 ± 0.11 | 7.9 ± 0.57 | 10.53 ± 0.62 | 15.03 ± 0.93 | 11.01 ± 0.62 |
| Enrichment | 1.25 ± 0.12 | 22.07 ± 0.5 | 7.03 ± 0.3 | 10.78 ± 0.58 | 18.06 ± 0.96 | 10.32 ± 0.64 |
| Light-Dark Test | Novel Tank Test | |||||
|---|---|---|---|---|---|---|
| Light Time (s) | Dark Time (s) | No. of Entries | Upper Time (s) | Bottom Time (s) | No. of Entries | |
| Control | 74.17 ± 24.82 | 405.83 ± 24.82 | 4.17 ± 2.29 | 65.17 ± 40.43 | 414.83 ± 40.43 | 16.5 ± 10.23 |
| Enrichment | 77.92 ± 43.21 | 402.08 ± 43.21 | 4.92 ± 0.92 | 41.08 ± 14.41 | 438.92 ± 14.41 | 9.8 ± 2.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Hou, M.; Su, L.; Qiu, N.; Yu, F.; Zou, X.; Wang, C.; Wang, J.; He, Y. The Effect of Environmental Enrichment on Laboratory Rare Minnows (Gobiocypris rarus): Growth, Physiology, and Behavior. Animals 2022, 12, 514. https://doi.org/10.3390/ani12040514
Xu C, Hou M, Su L, Qiu N, Yu F, Zou X, Wang C, Wang J, He Y. The Effect of Environmental Enrichment on Laboratory Rare Minnows (Gobiocypris rarus): Growth, Physiology, and Behavior. Animals. 2022; 12(4):514. https://doi.org/10.3390/ani12040514
Chicago/Turabian StyleXu, Chunsen, Miaomiao Hou, Liangxia Su, Ning Qiu, Fandong Yu, Xinhua Zou, Chunling Wang, Jianwei Wang, and Yongfeng He. 2022. "The Effect of Environmental Enrichment on Laboratory Rare Minnows (Gobiocypris rarus): Growth, Physiology, and Behavior" Animals 12, no. 4: 514. https://doi.org/10.3390/ani12040514
APA StyleXu, C., Hou, M., Su, L., Qiu, N., Yu, F., Zou, X., Wang, C., Wang, J., & He, Y. (2022). The Effect of Environmental Enrichment on Laboratory Rare Minnows (Gobiocypris rarus): Growth, Physiology, and Behavior. Animals, 12(4), 514. https://doi.org/10.3390/ani12040514