1. Introduction
Australian freshwater fish species are under threat of extinction due to continued habitat degradation and predation by invasive species [
1]. Small fish species, in particular, have been reported to be at a high risk of extinction within the next twenty-years without swift intervention to protect their natural habitat [
2]. However, such interventions can take time to be effective during which species remain vulnerable to further declines. The use of bio-banking is an ex situ means of securing fish genetics in the event of a sudden drop in genetic diversity or species extinction [
3,
4]. The cryopreservation of reproductive cells and tissues provides scientists an opportunity to reproduce and reintroduce individuals from a species even after they have gone extinct in the wild. While cryopreservation has been successful in preserving cells from male fish including sperm [
5] and spermatogonial cells [
6], there has been very limited success in the cryopreservation of female reproductive cells and tissues, with no reports in Australian fish species.
Fish oocytes are large and contain a high percentage of lipid and water which results in poor cryoprotectant permeability and a high risk of cryo-injury during cryopreservation [
7]. For these reasons, the recovery of viable, fertilisable oocytes post-thaw has been challenging. Fish embryos face similar limitations [
8] and even with technological advancements including the injection of cryoprotectants into the yolk [
9] and the use of gold nanoparticles and laser warming to improve thawing speed [
10,
11], post-thaw viability in fish embryos remains low. Cryopreservation has been somewhat more successful in smaller, immature oocytes [
12], however production of mature, fertilisable oocytes from immature, cryopreserved oocytes has not yet been achieved in vitro [
13]. In the absence of reliable oocyte and embryo cryopreservation methods, there are only limited opportunities to secure maternal fish genomes in biobanks. Without a robust method for the successful cryopreservation and warming of fish oocytes, the applications of sperm cryopreservation are limited to species in which mature oocytes are able to be collected from living fish for fertilisation [
5]. In the context of endangered species, this poses a significant limitation as the purpose of bio-banking is often to resurrect populations of significantly reduced or extinct species, at which point accessing spawning females may be extremely difficult or even impossible. The cryopreservation of ovarian tissue containing oogonia, the early cell stage of oogenesis, has become an increasingly popular target for the bio-banking of genetics of female fish. Cryopreservation of ovarian tissue has been reported in several species, including the common carp (
Cyprinus carpio) [
14], Piracanjunba (
Brycon orbignyanus) [
15], sea trout (
Salmo trutta) [
16], American paddlefish (
Polyodon spathula) [
17], Siberian sturgeon (
Acipenser baerii) [
18] and the endangered cyprinid honmoroko (
Gnathopogon caerulescens) [
19], representing families within the orders of Cypriniformes, Characiformes, Salmoniformes and Acipenseriformes.
Oogonia are relatively small, simple cells that are capable of differentiating into mature oocytes via oogenesis. They have the advantage of being able to be isolated from the ovaries of fish and cryopreserved at any life stage, regardless of their sexual maturity or spawning period. While oogonia are undifferentiated germ cells, they can be differentiated into viable oocytes via a surrogate species using a method commonly referred to as germ cell transplantation (GCT) or cell surrogacy [
20]. Briefly, gonial cells extracted from cryopreserved gonadal tissue of a threatened species, are injected into a sterile surrogate where they incorporate into the gonad, re-establish gametogenesis and eventually produce donor derived gametes.
Australia is home to a number of threatened freshwater fish species. Recent fish kills in the country’s largest freshwater river system, the Murray Darling Basin [
21], highlight the need for ex situ conservation methods including biobanking. To our knowledge there has been no investigation into the potential use of ovarian tissue cryopreservation in Australian fish species. In collaboration with the Australian Frozen Zoo, a Melbourne based biobanking initiative that has been operating for the past twenty-five years, we have been investigating the potential application of gonad cryopreservation in Australian fish species. We previously described successful cryopreservation of testicular tissue in the Murray River Rainbowfish (
Melanotaenia fluviatilis) [
22] from the family Melanotaeniidae (order Atheriniformes), a group of small freshwater fish species endemic to areas across Australia and surrounding regions including New Guinea. While
M. fluviatilis is listed as threatened within the state of Victoria [
23], this species also has several critically endangered and endangered relatives including the Running River Rainbowfish (
M. splendida. nov), the Slender Rainbowfish (
M. gracilis), the Lake Eacham Rainbowfish (
M. eachamensis), and the Utchee Creek Rainbowfish (
M. utcheensis) [
24].
Here we describe the successful cryopreservation of ovarian tissue in
M. fluviatilis; a protocol (data set in
Supplementary Materials) that could be applied to bio-banking programs for other species in Melanotaeniidae, and perhaps species in other families and orders of Australian fish. We have also investigated the use of flow cytometry as a method for isolating and analysing specific ovarian cell types in ovarian tissue suspensions. In the absence of successful oocyte or embryo cryopreservation protocols, the cryopreservation and storage of ovarian tissue from fish species presents an accessible target for current biobanking programs.
2. Materials and Methods
2.1. Animal Handling and Ethics Approval
Female
M. fluviatilis obtained from an aquarium supplier, Aquarium Industries (Epping, VIC, Australia), were held at 25 °C ± 1 °C on a 12:12 light-dark cycle. At the time of collection, fish weighing 1.33 g ± 0.72 g and 4.27 cm ± 0.7 cm in length (standard length) were killed by anaesthetic overdose by submersion in 175 mg/L of AQUI-S (Primo Aquaculture, Narangba, QLD, Australia) diluted in system water for 20 min. A total of thirty-eight (
n = 38) female
M. fluviatilis were used, with specific sample sizes for each experiment included below. Based on their length, the age of fish used in this study is estimated to be between one to two years old [
25]. Death was confirmed by manual destruction of the brain. Gonads weighing 0.047 g ± 0.04 g were removed and placed into chilled phosphate–buffered saline (PBS; pH 7.8). All animal handling and experimental procedures were performed in accordance with animal ethics application MMCB/2017/39 approved by the Animal Ethics Committee B at Monash Medical Centre and conducted in accordance with the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes.
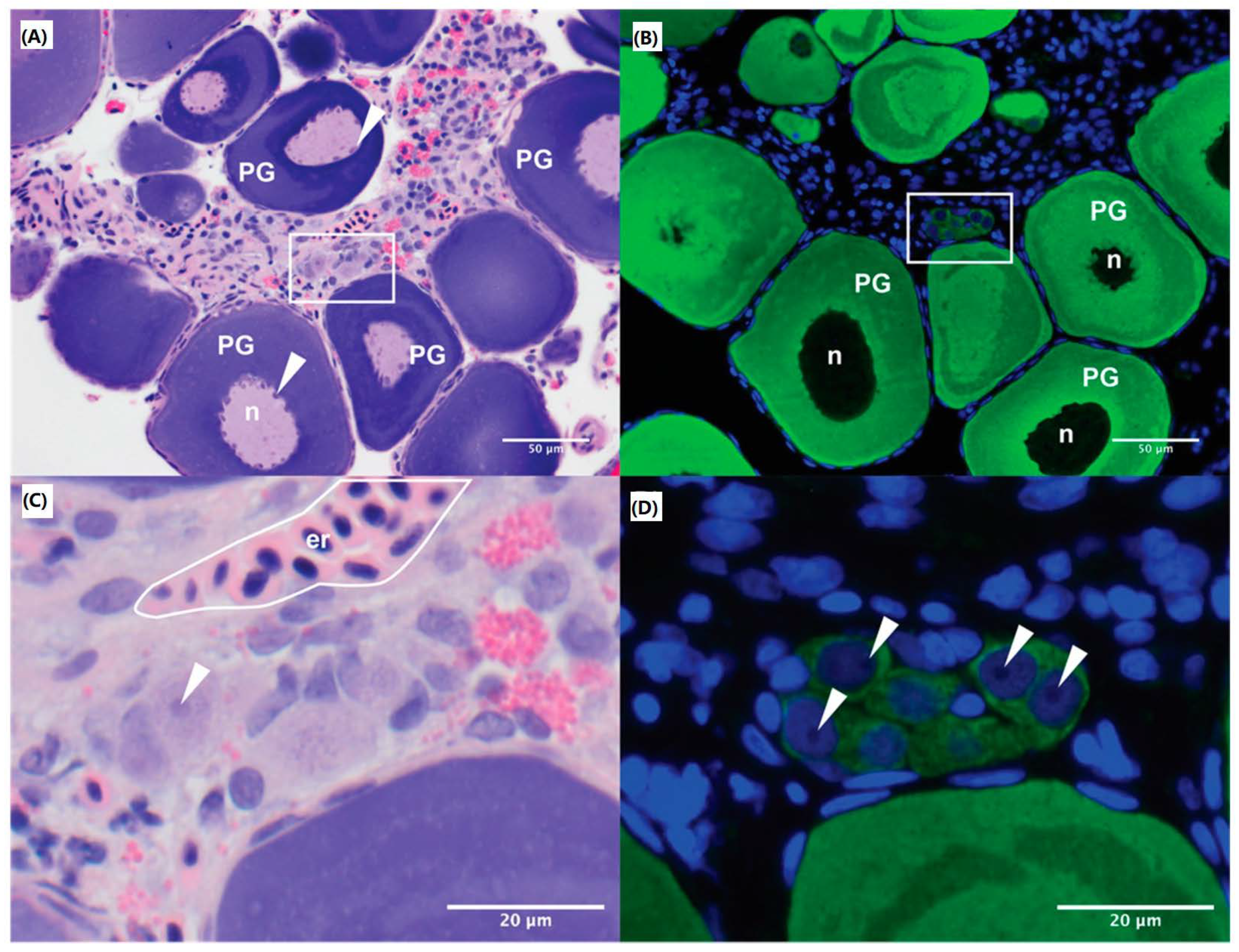
2.2. Histology & Immunohistochemistry
Whole ovaries from five (
n = 5)
M. fluviatilis were fixed in 10% neutral buffered formalin (Merck, VIC, Australia) for 48 h and processed by the Monash Histology Platform, which included standard haematoxylin and eosin staining. Unstained sections were stained for Vasa, to identify oogonia, using a zebrafish (
Danio rerio) specific anti-Vasa antibody (Sapphire Bioscience Pty. Ltd., Redfern, NSW, Australia) and counter stained with Hoechst (ThermoFisher Scientific, Waltham, MA, USA) as described in Rivers et al., 2020a. Briefly, de-paraffinised sections were rehydrated before antigen retrieval in 10 mM citrate buffer (pH 6). Sections were blocked with CAS Block (Invitrogen, Waltham, MA, USA) for one hour followed by incubation with anti-Vasa antibody (1:200) in diluent, 5% bovine serum albumin (BSA) in PBS, at 4 °C overnight. Sections were incubated with secondary antibody, Alexa Fluor 488-conjugated goat anti-rabbit IgG (1:500), and Hoechst nuclear counterstain (1:1000) in diluent before cover-slipping for imaging. Images were captured using the EVOS FL Auto 2 Imaging system (ThermoFisher Scientific) and Olympus BX43 Upright Microscope with an X-Cite Series 120 Q laser (Lumen Dynamics, Mississauga, ON, Canada). Sizes of oogonia were measured using cellSens Standard imaging software (Software version: 1.16, build 15404, Olympus, Tokyo, Japan) and images were analysed in FIJI [
26]. An estimation of the proportion of oogonia present in the ovary in comparison to total ovarian cell number was determined by counting the number of oogonia, based on size and Vasa-positive staining, and the total number of cells across five randomly sampled fields taken at 20× magnification with at least 250 cells present per field.
2.3. Cryopreservation and Thawing
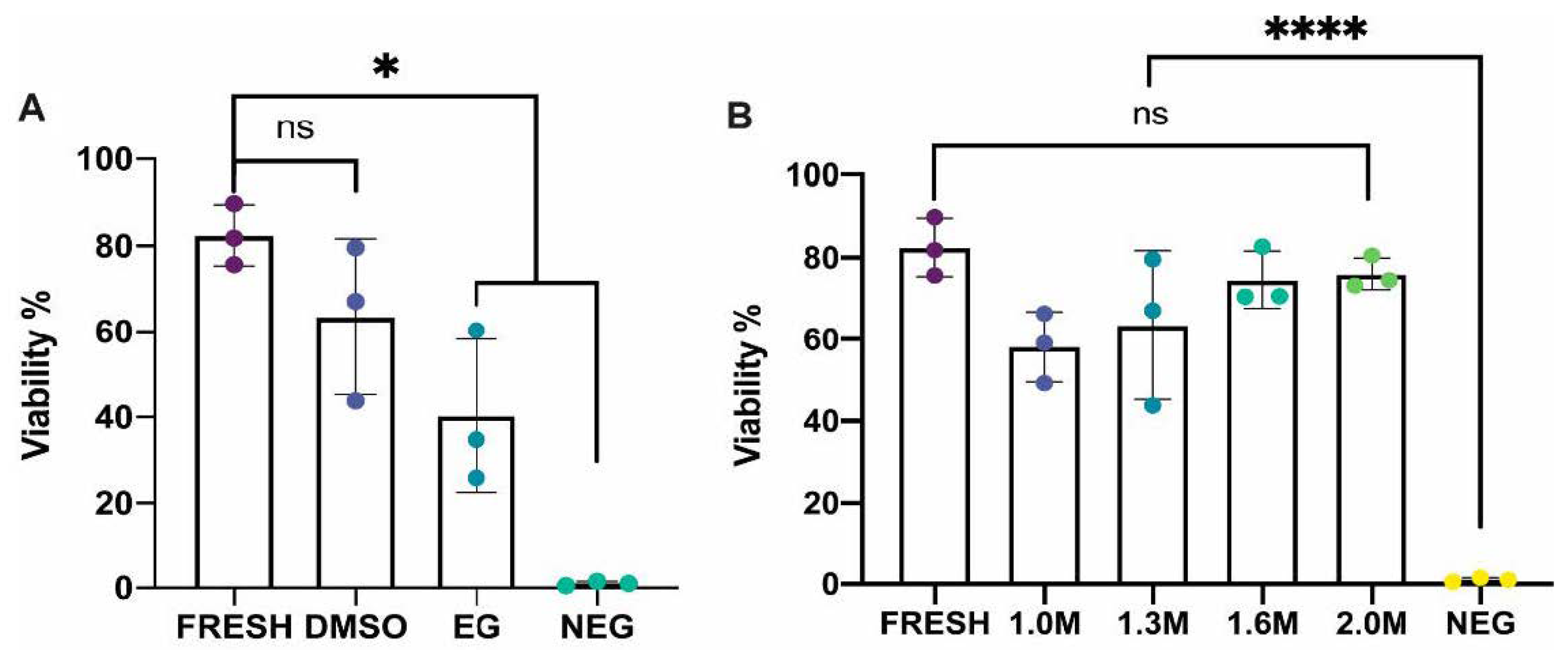
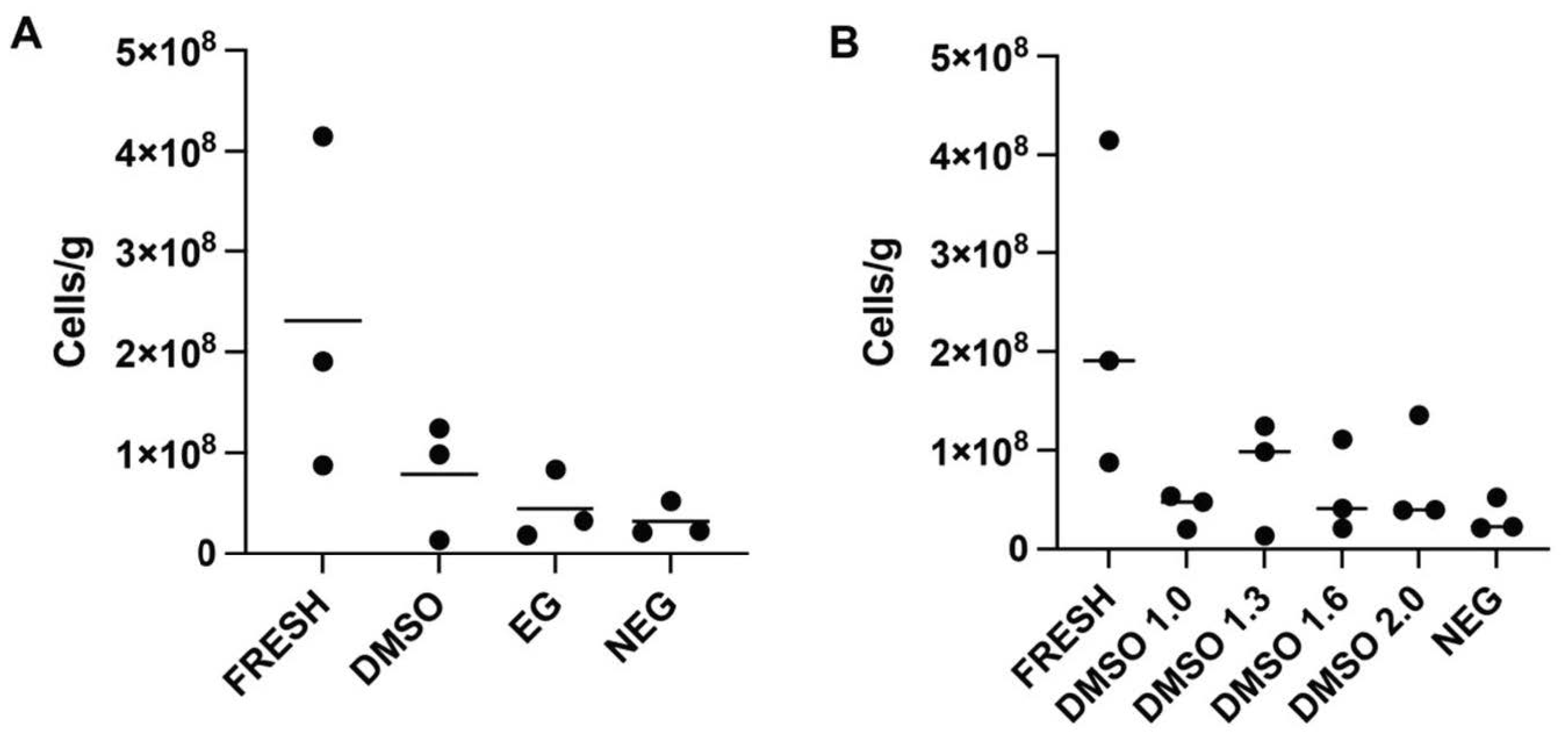
In experiment 1, the efficacy of two different permeating cryoprotectants was tested. Whole gonads from twelve (
n = 12) female
M. fluviatilis were divided across four treatment groups: fresh control, dimethyl sulfoxide (DMSO) treated, ethylene glycol (EG) treated, or negative control with each treatment assessed in triplicate. Ovaries in the fresh control group, were transferred into separate 1.2 mL CryoTubes with 1 mL trypsin-based digestion media prior to flow cytometry as described in
Section 2.4 and
Section 2.5. Ovaries undergoing cryopreservation were placed in a cryotube with 1ml of cryomedia containing a permeating cryoprotectant, either DMSO or EG, at a concentration of 1.3 M with 0.1 M trehalose (Merck, Darmstadt, Germany) and 1.5% BSA (Bovogen Biologicals Pty. Ltd., Keilor East, VIC, Australia) in a mixed salt extender previously described by [
27] (~300 mOsm, pH 7.8). Negative control samples contained all cryomedia components excluding the permeating cryoprotectant.
In experiment 2, DMSO was tested at a concentration of 1.0 M, 1.6 M and 2.0 M with each treatment assessed in triplicate (n = 9), with the same base media components as in experiment 1. Both experiments followed the same cooling and thawing method in which samples were equilibrated on ice for one hour and then cooled them at a rate of −1 °C/minute in a CoolCell LX cell freezing container (Corning, Glendale, AZ, USA) in a −80 °C freezer for at least 3 h before plunging them into liquid nitrogen where they were stored for at least 24 h before thawing.
Cryopreserved tissue was thawed in a 30 °C water bath for two minutes before rehydration in three changes of handling media (Eagles minimum essential media (EMEM, SigmaAldrich, St. Louis, MO, USA) supplemented with 5% FBS, ThermoFisher Scientific, Waltham, MA, USA; and 25 mM HEPES, ThermoFisher Scientific; pH 7.8) for 20 min per change.
2.4. Preparation of a Single Cell Suspension
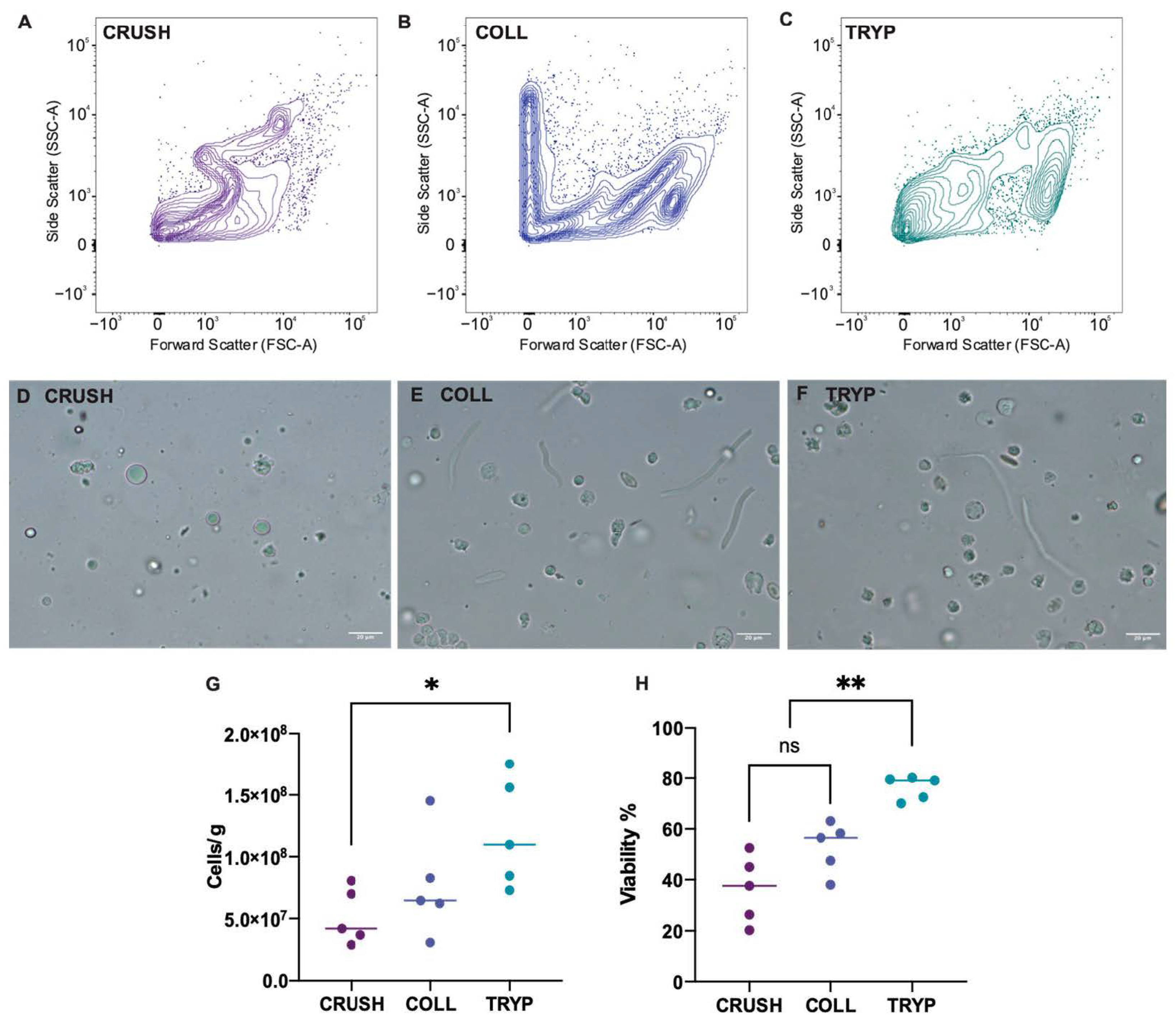
A single cell suspension was produced from fresh and frozen samples prior to flow cytometry. To optimize the cell suspension protocol, the ovary of five (n = 5) M. fluviatilis were split into three portions and assigned to one of the following cell suspension protocols—(i) mechanical crushing with a glass tissue grinder, (ii) digestion with 0.2% collagenase in PBS at 29 °C ± 1 °C for 2 h, or (iii) digestion in 0.25% trypsin with 1.13 mM EDTA in PBS for 30 min on ice and 1 h at room temperature. Digestion reactions using trypsin were stopped by addition of 2% serum. All samples were then filtered through 40 µm nylon mesh and cells were pelleted by centrifugation in a microfuge at 7000 rpm for 1 min at room temperature and resuspended in 1.13 mM EDTA in PBS for analysis by flow cytometry. The viability of cells 10 µm and larger was determined using the flow cytometer, and the total number of cells recovered per gram of tissue was determined using a haemocytometer.
2.5. Flow Cytometry
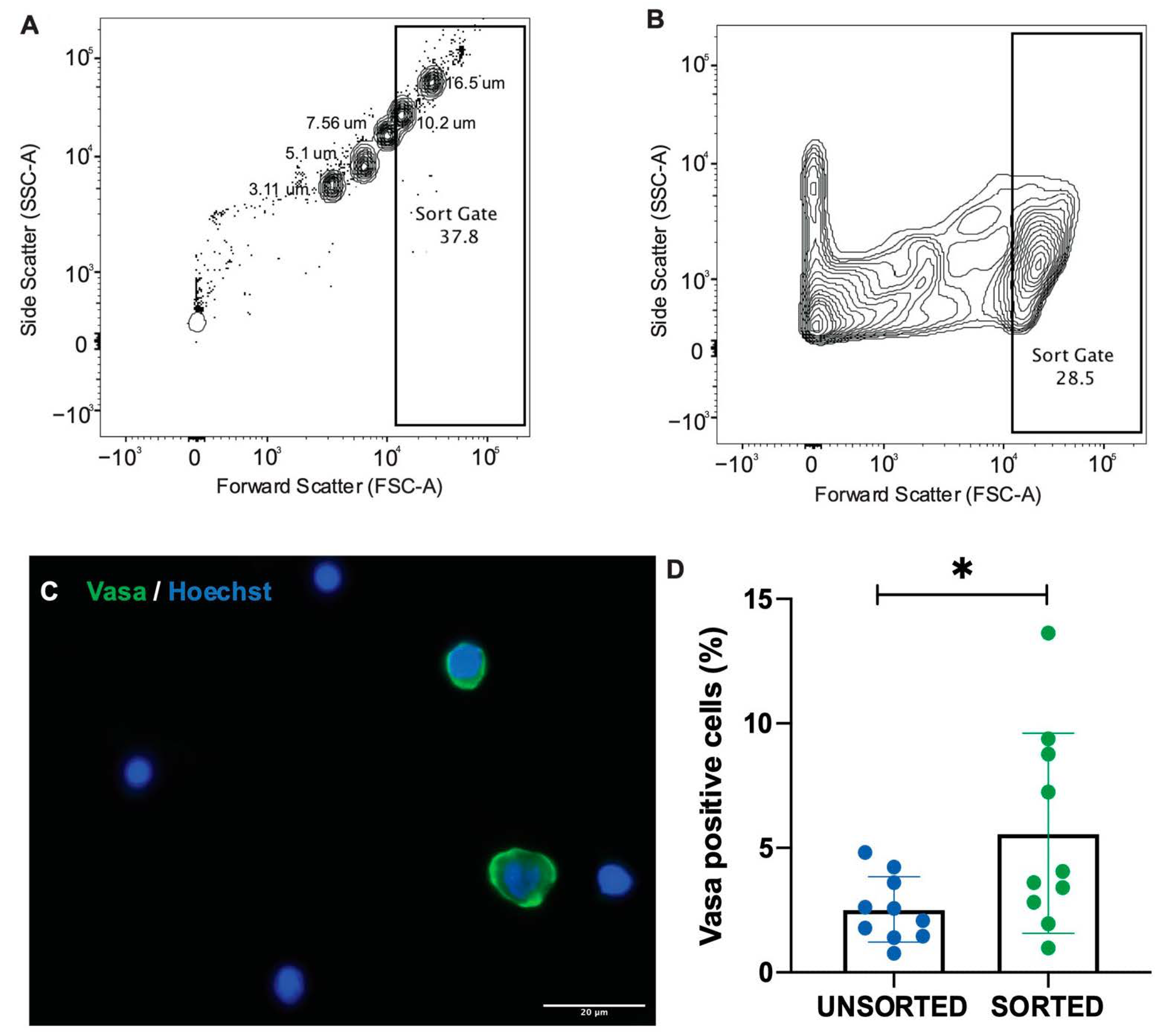
A set of size-specific beads (16.5 µm, 10.2 µm, 7.56 µm, 5.11 µm, 3.3 µm, Spherotech, Lake Forest, IL, USA) were run on the FACS Aria Fusion flow cytometer (BD Biosciences, Macquarie Park, NSW, Australia) prior to sample analysis to provide an approximate scale to determine the size of cells in suspension. Oogonia are the primary target when cryopreserving ovarian tissue. As such, it was important to determine the number of these cells able to be isolated from the single cell suspension using flow cytometry. To do this, ovaries from five (n = 5) M. fluviatilis were pooled into a single cell suspension and split into ten 500 uL samples, a 100 µL aliquot was taken to analyse the unsorted cells and the remaining sample was run through a flow cytometer. A gate was set to sort events approximately 10 µm and higher, capturing the approximate range of oogonia sizes as determined by histology. Live cells in the sorted and unsorted samples were imaged and measured to determine the range of sizes captured by the gate. The cells were then fixed in 2% formalin and dried onto SuperFrost plus slides overnight at 37 °C. Slides were stained with anti-Vasa antibody and Hoechst and imaged. Ten fields were selected randomly per sample and the number of Vasa-positive and Vasa-negative cells were counted to determine the average proportion of Vasa positive, oogonia in each sample analysed.
2.6. Viability Analysis
Viability analysis was performed for all samples from
Section 2.3 and
Section 2.4. At time of analysis, single cell suspensions were split across four 5 mL sample tubes in preparation for flow cytometry. Viability was determined using two vitality stains, Propidium Iodide (PI), a membrane impermeable dye that tags non-viable cells with membrane damage, and a membrane permeable counter-stain SYBR-14. One sample was kept as an unstained control, and single dye controls were prepared for SYBR14 and PI, with the PI control sample flash frozen in liquid nitrogen three times to ensure a high number of dead cells for analysis. Samples were incubated with SYBR14 (5 µL/mL) for 5 min in the dark, followed by a second 5 min incubation with PI (5 µL/mL). The flow cytometer was set to capture at least 50,000 events for each sample and flow cytometry output was analysed in FlowJo
TM Software for Mac [
28]. The viability of each sample was determined by the proportion of SYBR14 positive only events within all events captured by the sorting gate.
2.7. Statistical Analysis
Data analysis was performed in Prism 8 for MacOS (version 8.4.2) with a p-value of 0.05 considered statistically significant; all data are reported as mean ± standard deviation unless otherwise specified. Cell suspension viability, suspension cell recovery and cryopreservation viability data were all found to meet the assumptions of normality and variance using the Shapiro-Wilk test and the Brown-Forsythe test, respectively; data were then analysed by one-way ANOVA with Tukeys post hoc test. Comparison of the proportion of Vasa-positive cells in unsorted and sorted ovarian cell suspensions met the assumption of normality using the Shapiro-Wilk test and were analysed using an un-paired, two-tailed t-test.
4. Discussion
We have described protocols for successful cryopreservation of ovarian tissue in the Murray River rainbowfish (
Melanotaenia fluviatilis) and the isolation and viability analysis of ovarian cell suspensions using flow cytometry. Ovarian tissue from this species cryopreserved in the permeating cryoprotectant DMSO at concentrations between 1.0 to 2.0 M were all found to have viabilities comparable to fresh controls. This is consistent with our previous findings in testicular tissue in this species [
22]. Furthermore, this is the first report of ovarian tissue cryopreservation within the order Atheriniformes and the family Melanotaeniidae, providing a baseline method to begin the investigation of similar use of this biobanking method in other species within these groups.
Previous studies have used both slow cooling and vitrification methods to cryopreserve fish ovarian tissue. In transgenic rainbow trout, oogonia producing green fluorescence were isolated by flow cytometry and were found to have a viability of 72.9% when tissue was cryopreserved in 1.0 M DMSO, 0.1 M trehalose and 10% (
vol/
vol) egg yolk, using a slow cooling method [
29]. In carp, slow cooling resulted in approximately 65% viability in oogonia post thaw, using a cryomedia containing 1.5 M DMSO [
14]. Vitrification of ovarian tissue in salmonids [
16] and the cyprinid honmoroko [
19] resulted in oogonia viabilities of 37.98–40% and ~50%, respectively. While both slow cooling and vitrification have been used previously, slow cooling is preferable when working with threatened species as it is more flexible for in-field applications, requiring minimal equipment to begin the initial freezing process.
Post-thaw viability assessment is usually performed using the trypan blue exclusion assay as seen in the aforementioned studies in rainbow trout, carp, salmonid and cyprinid honmoroko. Here, we used flow cytometry as a tool to isolate cell populations from our fresh and cryopreserved ovarian cell suspensions and analyse their viability. Flow cytometry uses a fluidics and laser system to individually interrogate each cell in a suspension, gathering information that can help identify unique cell populations and sort specific cell types out of suspension. The analysis and sorting of fish testicular cells using flow cytometry has been reported in numerous species including starry goby (
Asterropteryx semipunctata) [
30] Pacific blue fin tuna (
Thunnus orientalis) [
31], Japanese char (
Salvelinus leucomaenis) and masu salmon (
Oncorhynchus masou) [
32]. We had also reported the use of flow cytometry to analyse testicular cell suspension in
M. fluviatilis [
22]. However, few studies have used this method to analyse ovarian cell suspensions in fish, with those that have reporting only in transgenic species [
29]. Here, we have analysed fish ovarian cell suspensions in a non-transgenic, native species. To do this we first had to produce a sufficient number of single ovarian cells in suspension for analysis. Similar studies in male fish have produced cell suspensions by crushing testicular tissue with a glass tissue grinder [
22,
30]. Tissue grinders are a time-efficient method to produce cell suspensions, however initial assessment of this method in the
M. fluviatilis ovary indicated that grinding alone was not sufficient to separate oogonia into suspension. Our histological analysis of the ovary of this species indicated that oogonia exist in closely bound nests within the germinal epithelium. To disrupt these tight bonds, we tested two digestion methods and found that ovarian cell suspensions produced using 0.25% trypsin with 1.13 mM EDTA in PBS yielded the highest overall recovery of cells per grams of tissue digested, and the highest cell viability. This has defined a successful method to produce a single cell suspension of ovarian cells in this species. These can be used in future downstream applications such as ovarian cell injection into a surrogate or in vitro cell culture.
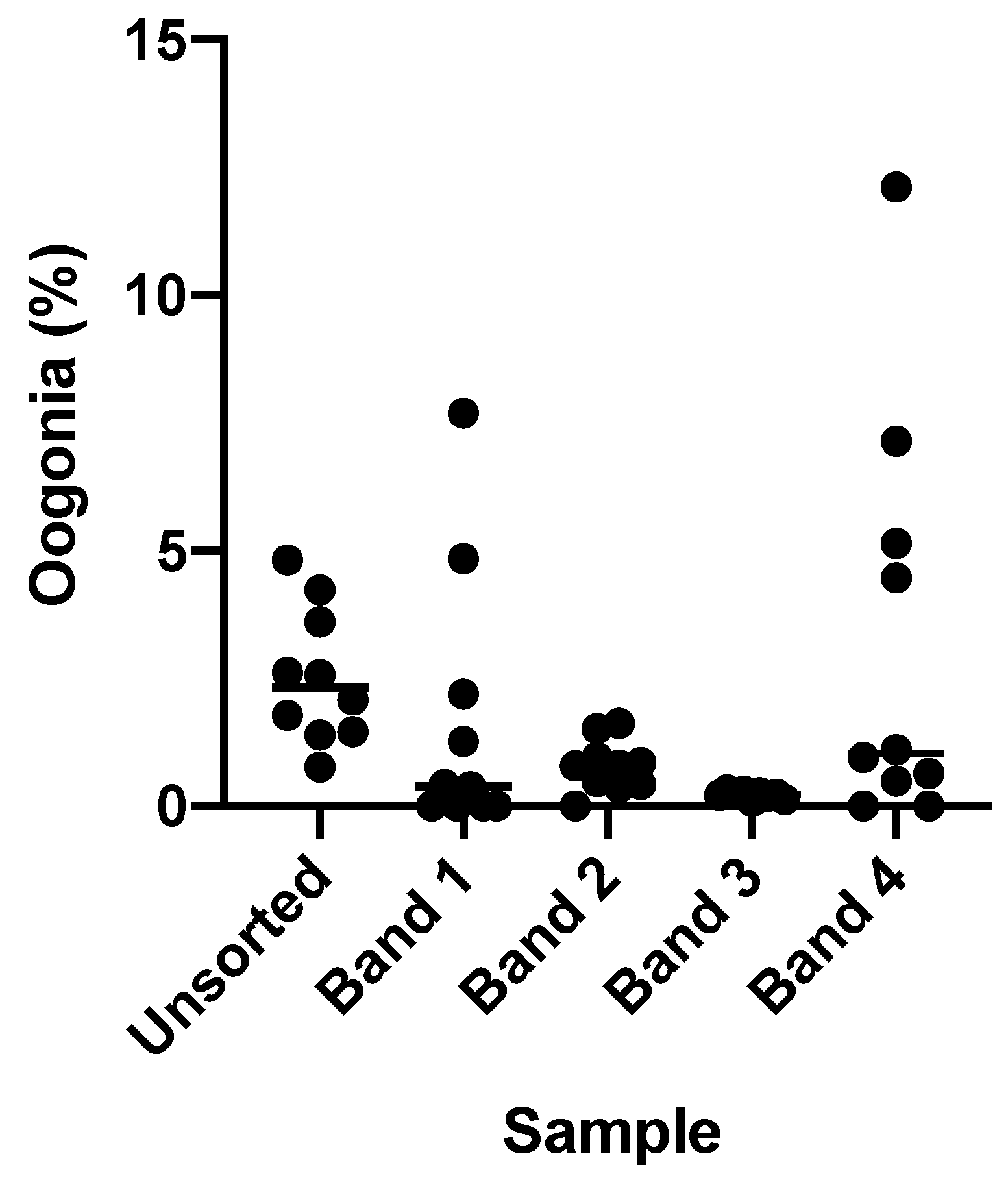
Flow cytometry is a fast and objective means of determining the viability of select population of cells. As oogonia were our primary focus in ovarian tissue cryopreservation, we wanted to determine if flow cytometry could be used to isolate oogonia from a single cell suspension. Other studies have often focused on gradient-based separation methods, with studies in smaller fish species such as zebrafish requiring ovaries from four or five individuals to produce a large enough cell volume to analyse [
33]. Methods designed to isolate specific cell populations, such as flow cytometry, may be better suited to isolate oogonia from small amounts of gonadal tissue. This is particularly important when considering conservation applications where access to tissue is often very limited. In this study we found that unsorted samples of ovarian cells in suspension contained approximately 2.53 ± 1.31% oogonia, which was consistent with histological estimations of the total proportion of oogonia in the ovary of 2.29 ± 0.81%. After sorting ovarian cell suspensions using the flow cytometer, the number of oogonia increased to 5.85 ± 4.01%, almost doubling the number of oogonia available compared to unsorted controls.
The advantage of gonad cryopreservation in comparison to gamete cryopreservation methods is that it can be performed using fish at any life stage, regardless of their sexual maturity and the natural spawning period of the fish. This is particularly important when considering in-field applications for cryopreservation methods, as techniques restricted to one phase of the reproductive cycle will significantly limit the number of animals that can be sampled. However, this can also result in variations in the number of oogonial cells available. The number of oogonia available in the ovary of our selected species was very low in comparison to previous studies, which have used immature or non-spawning individuals with a naturally higher proportion of oogonia present in the ovary. The fish used in this study were all of approximately reproductive age, with oocytes at various stages of oogenesis, and thereby contained a naturally low proportion of oogonial cells. It is therefore difficult to compare directly the recovery rate of this sorting method to previous studies. However, in our own pilot studies in this species, gradient centrifugation using a 20/40/60% AllGrad (CooperSurgical, Malov, Denmark) gradient did not result in any cell bands with an increased proportion of oogonia compared to an unsorted cell sample (
Figure A2). Producing a sufficiently enriched sample of oogonia is important for the success of downstream applications such as germ cell transplantation, and this will be more challenging in species were total oogonia counts are low. Here, we have shown the usefulness and efficiency of flow cytometry to isolate and enhance the number of oogonia from ovarian cell suspensions in species with low numbers of oogonia in the ovary.
While the cryopreservation of fish ovarian tissue has been more successful in comparison to the cryopreservation of more differentiated germ cell types such as mature oocytes and even whole embryos, oogonia isolated by any method from the ovary must be able to differentiate into viable, fertilisable oocytes in order to be able to produce new individuals. Transplantation of fish ovarian cells into sterile surrogates using germ cell transplantation techniques has resulted in the production of oocytes in two species of fish; pejerrey (
Odontesthes bonariensis) [
34] and rainbow trout (
Oncorhynchus mykiss) [
35]. Interspecies transplantation of oogonia and primordial germ cells has shown that oogenesis appears to be sensitive to species-related differences between the donor oogonia and the surrogate ovarian environment, often resulting in transplant failure [
4]. The production of oocytes relies on maternal depositing of germplasm and, later, vitellogenin to produce functional eggs [
36]. A better understanding of the species-specific requirements of oogenesis will allow us to select better surrogate species for transplantation. The interspecies transplantation of oogonia maybe one method to identify the critical requirements needed for oogenesis to occur in a surrogate environment. Identifying the stage at which cell transplantation fails, will enable investigation of why this failure has occurred and help to determine ways in which to correct it or identify surrogates that may be more suitable. For example, if oogonia do not colonise the gonad at all, this may suggest important differences in cell signaling pathways between species. Alternatively, if oogonia are able to colonise the gonad but fail to enter the gonadotrophin-dependent secondary growth phase, endocrinological differences may be the cause of failure. In this way, the interspecies transplantation of oogonia is not just an assisted reproductive technique that may be useful in conservation or aquaculture, but a tool to study the critical processes of oogenesis in fish species. Surrogate selection and species relatedness will be particularly important in the context of conservation. Studies in frogs have shown closely related species often face similar environmental pressures and threats [
37]. If the same is true of fish species, closely related surrogates may not always be available if these close relatives are also facing their own conservation concerns. A better understanding of the mechanisms behind oogenesis and species related conflicts that arise after germ cell transplantation, will inform better surrogate selection in future transplantation studies in endangered fish species.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





