
An Investigation of Ovarian and Adrenal Hormone Activity in Post-Ovulatory Cheetahs (Acinonyx jubatus)

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Husbandry
2.2. Sample Collection and Preparation
2.3. Fecal Glucocorticoid Metabolite (fGM) Analysis
2.4. Fecal Estrogen Metabolite (fEM) Analysis
2.5. Fecal and Urinary Progestagen Metabolite Analysis
2.6. Statistical Analysis, Calculations and Definitions
3. Results
3.1. Ovulatory Condition and Time Period Comparisons
3.2. Fecal Hormone Metabolite Concentrations in Confirmed Pregnant Females
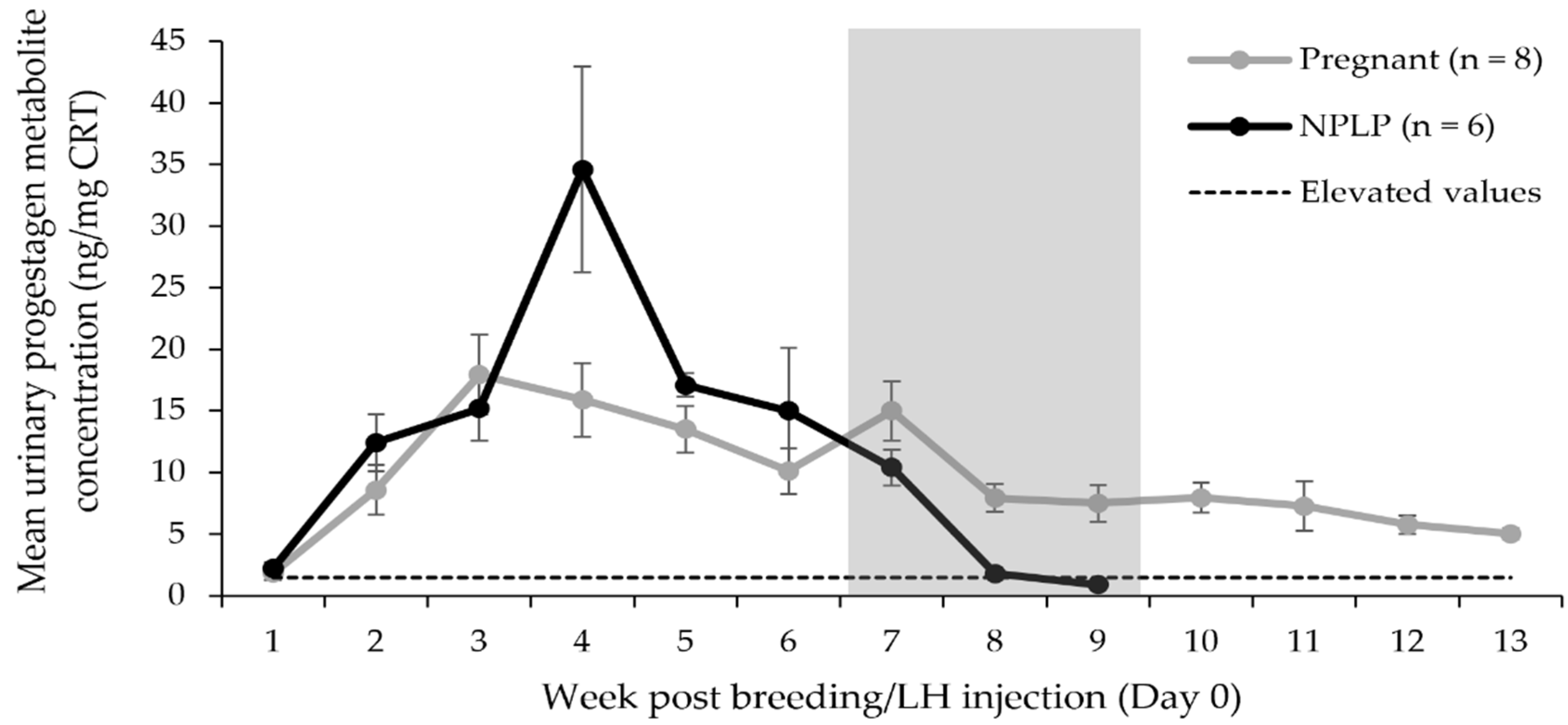
3.3. Urinary Progestagen Metabolite Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Durant, S.M.; Mitchell, N.; Groom, R.; Pettorelli, N.; Ipavec, A.; Jacobson, A.P.; Woodroffe, R.; Böhm, M.; Hunter, L.T.; Becker, M.S.; et al. The global decline of cheetah Acinonyx jubatus and what it means for conservation. Proc. Natl. Acad. Sci. USA 2017, 114, 528–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crosier, A.; Meeks, K.; Maloney, E.; Andrews, J. AZA Species Survival Plan® Yellow Program Population Analysis and Treeding and Transfer Plan for Cheetah (Acinonyx jubatus); AZA Population Management Center: Chicago, IL, USA, 2021. [Google Scholar]
- Dierenfeld, E.S. Nutrition of captive cheetahs: Food composition and blood parameters. Zoo Biol. 1993, 12, 143–150. [Google Scholar] [CrossRef]
- Vester, B.M.; Beloshapka, A.N.; Middelbos, I.S.; Burke, S.L. Evaluation of nutrient digestibility and fecal characteristics of exotic felids fed horse- or beef-based diets: Use of the domestic cat as a model for exotic felids. Zoo Biol. 2010, 29, 432–448. [Google Scholar] [CrossRef] [PubMed]
- Wielebnowski, N.; Brown, J.L. Behavioral correlates of physiological estrus in cheetahs. Zoo Biol. 1998, 17, 193–209. [Google Scholar] [CrossRef]
- Wielebnowski, N.C. Behavioral differences as predictors of breeding status in captive cheetahs. Zoo Biol. 1999, 18, 335–349. [Google Scholar] [CrossRef]
- Koester, D.C.; Freeman, E.W.; Brown, J.L.; Wildt, D.E.; Terrell, K.A.; Franklin, A.D.; Crosier, A.E. Motile sperm output by male cheetahs (Acinonyx jubatus) managed ex situ is influenced by public exposure and number of care-givers. PLoS ONE 2015, 10, e0135847. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Miranda, C.R.; Wells, S.A.; Golden, R.; Seidensticker, J. Vocalizations and other behavioral responses of male cheetahs (Acinonyx jubatus) during experimental separation and reunion trials. Zoo Biol. 1998, 17, 1–16. [Google Scholar] [CrossRef]
- Wielebnowski, N.C.; Ziegler, K.; Wildt, D.E.; Lukas, J.; Brown, J.L. Impact of social management on reproductive, adrenal and behavioral activity in the cheetah (Acinonyx jubatus). Anim. Conserv. 2002, 5, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Koester, D.C.; Freeman, E.W.; Wildt, D.E.; Terrell, K.A.; Franklin, A.D.; Meeks, K.; Crosier, A.E. Group management influences reproductive function of the male cheetah (Acinonyx jubatus). Reprod. Fertil. Dev. 2015; Epub ahead of print. [Google Scholar] [CrossRef]
- Munson, L.; Terio, K.A.; Worley, M.; Jago, M.; Bagot-Smith, A.; Marker, L. Extrinsic factors significantly affect patterns of disease in free-ranging and captive cheetah (Acinonyx jubatus) populations. J. Wildl. Dis. 2005, 41, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Munson, L. Diseases of captive cheetahs (Acinonyx jubatus): Results of the cheetah research council pathology survey, 1989–1992. Zoo Biol. 1993, 12, 105–124. [Google Scholar] [CrossRef]
- Terio, K.A.; Munson, L.; Moore, P.F. Characterization of the gastric immune response in cheetahs (Acinonyx jubatus) with Helicobacter-associated gastritis. Vet. Pathol. 2012, 49, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Franklin, A.D.; Crosier, A.E.; Vansandt, L.M.; Mattson, E.; Xiao, Z. Induction of cytokine production in cheetah (Acinonyx jubatus) peripheral blood mononuclear cells and validation of feline-specific cytokine assays for analysis of cheetah serum. J. Zoo Wildl. Med. 2015, 46, 306–313. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.J.; Wildt, D.E.; Goldman, D.; Merril, C.R.; Bush, M. The cheetah is depauperate in genetic variation. Science 1983, 221, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Dobrynin, P.; Liu, S.; Tamazian, G.; Xiong, Z.; Yurchenko, A.A.; Krasheninnikova, K.; Kliver, S.; Schmidt-Kuntzel, A.; Koepfli, K.-P.; Johnson, W.; et al. Genomic legacy of the African cheetah, Acinonyx jubatus. Genome Biol. 2015, 16, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.L.; Graham, L.H.; Wielebnowski, N.; Swanson, W.F.; Wildt, D.E.; Howard, J.G. Understanding the basic reproductive biology of wild felids by monitoring of faecal steroids. In The Fourth International Symposium on Canine and Feline Reproduction: Advances in Reproduction in Dogs, Cats and Exotic Carnivores; Concannon, P.W., England, G.C.W., Farstad, W., Linde-Forsberg, C., Verstegen, J.P., Doberska, C., Eds.; Journal of Reproduction and Fertility: Middlesex, UK, 2001; pp. 71–82. [Google Scholar]
- Brown, J.L.; Wildt, D.E. Assessing reproductive status in wild felids by non-invasive faecal steroid monitoring. Int. Zoo Yearb. 1997, 35, 173–191. [Google Scholar] [CrossRef]
- Crosier, A.E.; Comizzoli, P.; Baker, T.; Davidson, A.; Munson, L.; Howard, J.G.; Marker, L.L.; Wildt, D.E. Increasing age influences uterine integrity, but not ovarian function or oocyte quality, in the cheetah (Acinonyx jubatus). Biol. Reprod. 2011, 85, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Howard, J.G.; Munson, L.; McAloose, D.; Kriete, M.; Bush, M.; Wildt, D.E. Comparative evaluation of seminal, vaginal, and rectal bacterial flora in the cheetah and domestic cat. Zoo Biol. 1993, 12, 81–96. [Google Scholar] [CrossRef]
- Howard, J.G.; Roth, T.L.; Byers, A.P.; Swanson, W.F.; Wildt, D.E. Sensitivity to exogenous gonadotropins for ovulation induction and laparoscopic artificial insemination in the cheetah and clouded leopard. Biol. Reprod. 1997, 56, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Terrell, K.A.; Wildt, D.E.; Anthony, N.M.; Bavister, B.D.; Leibo, S.P.; Penfold, L.M.; Marker, L.L.; Crosier, A.E. Different patterns of metabolic cryo-damage in domestic cat (Felis catus) and cheetah (Acinonyx jubatus) spermatozoa. Cryobiology 2012, 64, 110–117. [Google Scholar] [CrossRef]
- Wildt, D.E.; Grisham, J. Basic research and the cheetah SSP program. Zoo Biol. 1993, 12, 3–4. [Google Scholar] [CrossRef]
- Koester, D.C.; Wildt, D.E.; Maly, M.; Comizzoli, P.; Crosier, A.E. Non-invasive identification of protein biomarkers for early pregnancy diagnosis in the cheetah (Acinonyx jubatus). PLoS ONE 2017, 12, e0188575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byron, M.J.; Koester, D.C.; Edwards, K.L.; Mozdziak, P.E.; Farin, C.E.; Crosier, A.E. Immunoglobulin J chain as a non-invasive indicator of pregnancy in the cheetah (Acinonyx jubatus). PLoS ONE 2020, 15, e0225354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maly, M.A.; Edwards, K.L.; Farin, C.E.; Koester, D.C.; Crosier, A.E. Assessing puberty in ex situ male cheetahs (Acinonyx jubatus) via fecal hormone metabolites and body weights. Gen. Comp. Endocrinol. 2018, 268, 22–33. [Google Scholar] [CrossRef]
- Maly, M.A.; Edwards, K.L.; Koester, D.C.; Farin, C.E.; Crosier, A.E. Assessing puberty in female cheetahs (Acinonyx jubatus) via faecal hormone metabolites and body weight. Reprod. Fertil. Dev. 2021, 33, 841–854. [Google Scholar] [CrossRef]
- Crosier, A.E.; Lamy, J.; Bapodra, P.; Rapp, S.; Maly, M.; Junge, R.; Haefele, H.; Ahistus, J.; Santiestevan, J.; Comizzoli, P. First birth of cheetah cubs from in vitro fertilization and embryo transfer. Animals 2020, 10, 1811. [Google Scholar] [CrossRef]
- Wildt, D.E.; Brown, J.L.; Bush, M.; Barone, M.A.; Cooper, K.A.; Grisham, J.; Howard, J.G. Reproductive status of the cheetah (Acinonyx jubatus) in North American zoos: The benefits of physiological surveys for strategic planning. Zoo Biol. 1993, 12, 45–80. [Google Scholar] [CrossRef]
- Crosier, A.E.; Pukazhenthi, B.S.; Henghali, J.N.; Howard, J.G.; Dickman, A.J.; Marker, L.; Wildt, D.E. Cryopreservation of spermatozoa from wild-born Namibian cheetahs (Acinonyx jubatus) and influence of glycerol on cryosurvival. Cryobiology 2006, 52, 169–181. [Google Scholar] [CrossRef]
- Crosier, A.E.; Henghali, J.N.; Howard, J.; Pukazhenthi, B.S.; Terrell, K.A.; Marker, L.L.; Wildt, D.E. Improved quality of cryopreserved cheetah (Acinonyx jubatus) spermatozoa after centrifugation through Accudenz. J. Androl. 2009, 30, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Howard, J.G.; Donoghue, A.M.; Barone, M.A.; Goodrowe, K.L.; Blumer, E.S.; Snodgrass, K.; Starnes, D.; Tucker, M.; Bush, M.; Wildt, D.E. Successful induction of ovarian activity and laparoscopic intrauterine artificial insemination in the cheetah (Acinonyx jubatus). J. Zoo Wildl. Med. 1992, 23, 288–300. [Google Scholar]
- Crosier, A.E.; Comizzoli, P.; Koester, D.C.; Wildt, D.E. Circumventing the natural, frequent estrogen waves of the female cheetah (Acinonyx jubatus) using oral progestin (Altrenogest). Reprod. Fertil. Dev. 2016, 29, 1486–1498. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, A.M.; Howard, J.G.; Byers, A.P.; Goodrowe, K.L.; Bush, M.; Blumer, E.; Lukas, J.; Stover, J.; Snodgrass, K.; Wildt, D.E. Correlation of sperm viability with gamete interaction and fertilization in vitro in cheetah (Acinonyx jubatus). Biol. Reprod. 1992, 46, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Roth, T.L.; Swanson, W.F.; Blumer, E.; Wildt, D.E. Enhancing zona penetration by spermatozoa from a teratospermic species, the cheetah (Acinonyx jubatus). J. Exp. Zool. 1995, 271, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Wildt, D.E.; Wielebnowski, N.; Goodrowe, K.L.; Wells, S.; Graham, L.H.; Howard, J.G. Reproductive activity in captive female cheetahs (Acinonyx jubatus) assessed by faecal steroids. J. Reprod. Fertil. 1996, 106, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Thatcher, M.-J.D.; Shille, V.M.; Fliss, M.F.; Bazer, F.W.; Sisum, W.; Randal, S. Characterization of feline conceptus proteins during pregnancy. Biol. Reprod. 1991, 44, 108–120. [Google Scholar] [CrossRef]
- Young, K.M.; Walker, S.L.; Lanthier, C.; Waddell, W.T.; Monfort, S.L.; Brown, J.L. Noninvasive monitoring of adrenocortical activity in carnivores by fecal glucocorticold analyses. Gen. Comp. Endocrinol. 2004, 137, 148–165. [Google Scholar] [CrossRef]
- Koester, D.C.; Wildt, D.E.; Brown, J.L.; Meeks, K.; Crosier, A.E. Public exposure and number of conspecifics have no influence on ovarian and adrenal activity in the cheetah (Acinonyx jubatus). Gen. Comp. Endocrinol. 2017, 243, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Adachi, I.; Kusuda, S.; Kawai, H.; Ohazama, M.; Taniguchi, A.; Kondo, N.; Yoshihara, M.; Okuda, R.; Ishikawa, T.; Kanda, I.; et al. Fecal progestagens to detect and monitor pregnancy in captive female cheetahs (Acinonyx jubatus). J. Reprod. Dev. 2011, 57, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Wells, A.; Terio, K.A.; Ziccardi, M.H.; Munson, L. The stress response to environmental change in captive cheetahs (Acinonyx jubatus). J. Zoo Wildl. Med. 2004, 35, 8–14. [Google Scholar] [CrossRef]
- Tilbrook, A.J.; Turner, A.I.; Clarke, I.J. Effects of stress on reproduction in non-rodent mammals: The role of glucocorticoids and sex differences. Rev. Reprod. 2000, 5, 105–113. [Google Scholar] [CrossRef]
- Blank, M.H.; Adania, C.H.; Swanson, W.F.; de Souza Ramos Angrimani, D.; Nichi, M.; Alcindo de Barros Vaz Guimarães, M.; Barnabe, R.C. Comparative fecal steroid profile during pregnancy, parturition, and lactation between natural fertilization and embryo transfer in ocelots (Leopardus pardalis). Theriogenology 2022, 182, 26–34. [Google Scholar] [CrossRef] [PubMed]
- MacDougall-Shackleton, S.A.; Bonier, F.; Romero, L.M.; Moore, I.T. Glucocorticoids and “stress” are not synonymous. Integr. Org. Biol. 2019, 1, obz017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelican, K.M.; Wildt, D.E.; Pukazhenthi, B.; Howard, J. Ovarian control for assisted reproduction in the domestic cat and wild felids. Theriogenology 2006, 66, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.H.; Brown, J.L. Cortisol metabolism in the domestic cat and implications for developing a non-invasive measure of adrenocorticol activity in non-domestic felids. Zoo Biol. 1996, 15, 71–82. [Google Scholar] [CrossRef]
- Brown, J.L.; Wasser, S.K.; Wildt, D.E.; Graham, L.H. Comparative aspects of steroid hormone metabolism and ovarian activity in felids, measured non-invasively in feces. Biol. Reprod. 1994, 51, 776–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, C.J.; Stabenfeldt, G.H.; Cragun, J.R.; Addiego, L.A.; Overstreet, J.W.; Lasley, B.L. Relationship of serum estradiol and progesterone concentrations to the excretion profiles of their major urinary metabolites as measured by enzyme immunoassay and radioimmunoassay. Clin. Chem. 1991, 37, 838–844. [Google Scholar] [CrossRef]
- Amendolagine, L.; Schoffner, T.; Koscielny, L.; Schook, M.; Copeland, D.; Casteel, J.; Duff, B.; Koester, D. In-house monitoring of steroid hormone metabolites in urine informs breeding management of a giant anteater (Myrmechophaga tridactyla). Zoo Biol. 2018, 37, 40–45. [Google Scholar] [CrossRef]
- Verhage, H.G.; Beamer, N.B.; Brenner, R.M. Plasma levels of estradiol and progesterone in the cat during polyestrus, pregnancy and pseudopregnancy. Biol. Reprod. 1976, 14, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, K.; Inada, S.; Seki, K.; Sasaki, A.; Hama, N.; Kusunoki, H. Long-term monitoring of fecal steroid hormones in female snow leopards (Panthera uncia) during pregnancy or pseudopregnancy. PLoS ONE 2011, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Putman, S.B.; Brown, J.L.; Franklin, A.D.; Schneider, E.C.; Boisseau, N.P.; Asa, C.S.; Pukazhenthi, B.S. Characterization of ovarian steroid patterns in female African lions (Panthera leo), and the effects of contraception on reproductive function. PLoS ONE 2015, 10, e0140373. [Google Scholar] [CrossRef]
- Allen, W.R. Maternal recognition and maintenance of pregnancy in the mare. Anim. Reprod. 2005, 2, 209–223. [Google Scholar]
- Ginther, O.J.; Knopf, L.; Kastelic, J.P. Ovarian follicular dynamics in heifers during early pregnancy. Biol. Reprod. 1989, 41, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Edwards, P.D.; Boonstra, R. Glucocorticoids and CBG during pregnancy in mammals: Diversity, pattern, and function. Gen. Comp. Endocrinol. 2018, 259, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Korgun, E.T.; Ozmen, A.; Unek, G.; Mendilcioglu, I. The effects of glucocorticoids on fetal and placental development. In Glucocorticoids-New Recognition of Our Familiar Friend; London, UK. 2012. Available online: https://books.google.com.hk/books?hl=zh-CN&lr=&id=F_qdDwAAQBAJ&oi=fnd&pg=PA305&dq=The+effects+of+glucocorticoids+on+fetal+and+placental+development.&ots=xAJSrS8Usg&sig=TpYGZEMEoSyvTjUhp-otJ3Sl3as&redir_esc=y&hl=zh-CN&sourceid=cndr#v=onepage&q=The%20effects%20of%20glucocorticoids%20on%20fetal%20and%20placental%20development.&f=false (accessed on 18 March 2022).
- Dettmer, A.M.; Rosenberg, K.L.; Suomi, S.J.; Meyer, J.S.; Novak, M.A. Associations between parity, hair hormone profiles during pregnancy and lactation, and infant development in rhesus monkeys (Macaca mulatta). PLoS ONE 2015, 10, e0131692. [Google Scholar] [CrossRef]
- Hinde, K.; Skibiel, A.L.; Foster, A.B.; Del Rosso, L.; Mendoza, S.P.; Capitanio, J.P. Cortisol in mother’s milk across lactation reflects maternal life history and predicts infant temperament. Behav. Ecol. 2015, 26, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Milani, C.; Rota, A.; Olsson, U.; Paganotto, A.; Holst, B.S. Serum concentration of mineralocorticoids, glucocorticoids, and sex steroids in peripartum bitches. Domest. Anim. Endocrinol. 2021, 74, 106558. [Google Scholar] [CrossRef]
- Petrullo, L.; Lu, A. Natural variation in fetal cortisol exposure is associated with neonatal body mass in captive vervet monkeys (Chlorocebus aethiops). Am. J. Primatol. 2019, 81, e22943. [Google Scholar] [CrossRef]
- Alekseeva, G.S.; Loshchagina, J.A.; Erofeeva, M.N.; Naidenko, S.V. Stressed by maternity: Changes of cortisol level in lactating domestic cats. Animals 2020, 10, 903. [Google Scholar] [CrossRef]
- Carrera, S.C.; Sen, S.; Heistermann, M.; Lu, A.; Beehner, J.C. Low rank and primiparity increase fecal glucocorticoid metabolites across gestation in wild geladas. Gen. Comp. Endocrinol. 2020, 293, 113494. [Google Scholar] [CrossRef]
- Ryan, C.P.; Anderson, W.G.; Berkvens, C.N.; Hare, J.F. Maternal gestational cortisol and testosterone are associated with trade-offs in offspring sex and number in a free-living rodent (Urocitellus richardsonii). PLoS ONE 2014, 9, e111052. [Google Scholar]
- Pratt, N.; Lisk, R. Effects of social stress during early pregnancy on litter size and sex ratio in the golden hamster (Mesocricetus auratus). Reproduction 1989, 87, 763–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götz, A.A.; Wolf, M.; Stefanski, V. Psychosocial maternal stress during pregnancy: Effects on reproduction for F0 and F1 generation laboratory rats. Physiol. Behav. 2008, 93, 1055–1060. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Females | Ovulation Events |
|---|---|---|
| Pregnant | Feces: 8; Urine: 4 | Feces: 15; Urine: 8 |
| NPLP natural | Feces: 5; Urine: 2 | Feces: 7; Urine: 4 |
| NPLP AI | Feces: 4; Urine: 1 | Feces: 5; Urine: 2 |
| Parameter | Pregnant (n = 15) | NPLP Natural (n = 7) | NPLP AI (n = 5) | Published Value 1 [Source] |
|---|---|---|---|---|
| Fecal progestagen metabolites: | ||||
| Time to elevation (days) | 5.9 ± 0.48 | 5.57 ± 0.43 | 5.8 ± 1.28 | 3–4 [40] |
| Length of ovulation event (days) | 92.60 ± 0.40 | 60.29 ± 4.75 | 55.20 ± 2.73 | Preg: 94.2 ± 0.5 [36] Preg: 92.8 ± 0.4 [40] |
| Pre-breeding/LH injection baseline (μg/g) | 1.58 ± 0.38 | 1.10 ± 0.36 | 1.28 ± 0.74 | 0.92 ± 0.2 [25] |
| Mean of elevated values (μg/g) | 38.32 ± 3.58 | 30.74 ± 4.63 | 36.99 ± 9.22 | 40–70 [40] Preg: 28.7 ± 6.5 [25] NPLP: 43.9 ± 12.7 [25] |
| Peak (μg/g) | 120.78 ± 18.86 | 97.14 ± 16.57 | 110.12 ± 30.82 | |
| Peak post-breeding/LH injection (day) | 39.60 ± 4.33 | 31.14 ± 2.52 | 29.8 ± 4.73 | |
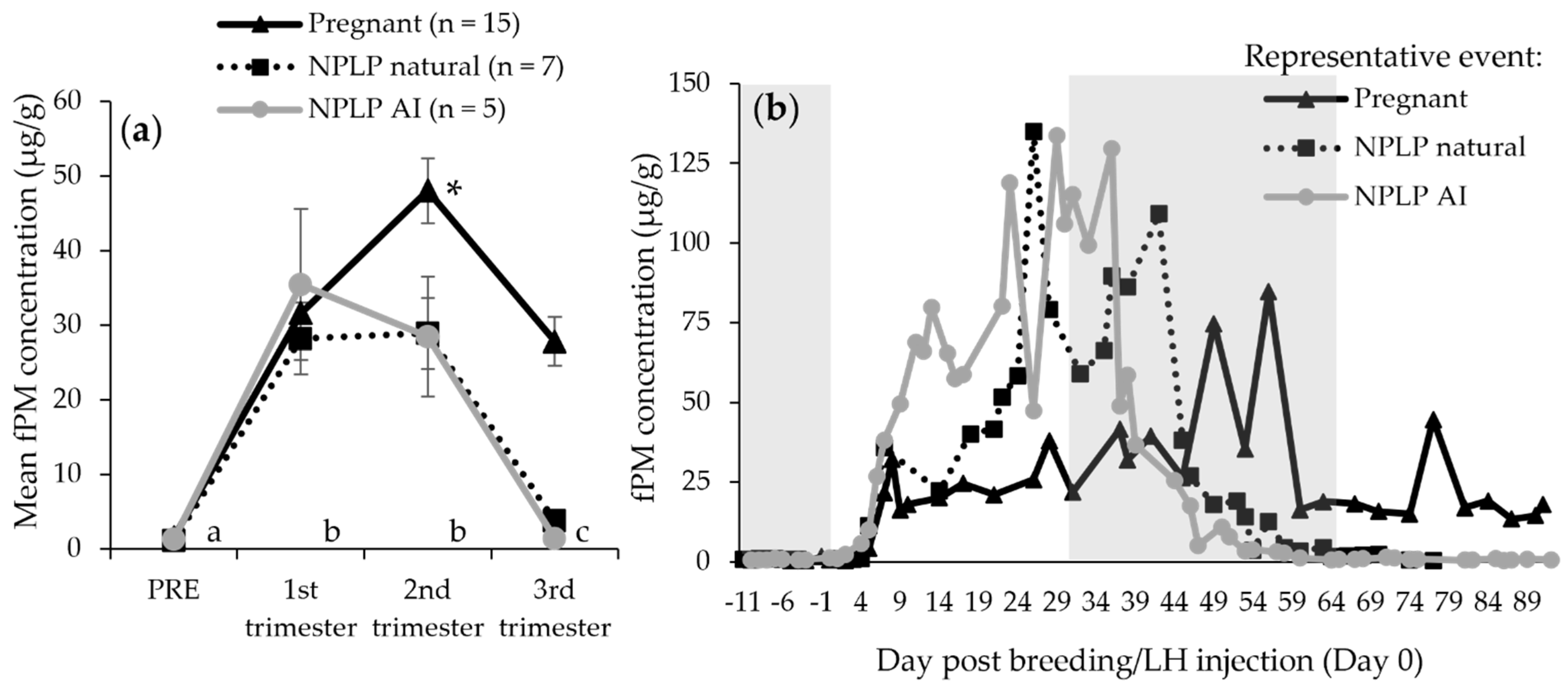
| First trimester mean (μg/g) | 31.63 ± 3.61 | 28.12 ± 4.80 | 35.43 ± 10.13 | |
| Second trimester mean (μg/g) | 48.05 ± 4.37 | 28.91 ± 4.79 | 28.45 ± 8.02 | |
| Third trimester mean (μg/g) | 27.85 ± 3.30 | 3.775 ± 1.37 | 1.43 ± 0.47 | |
| Fecal estrogen metabolites: | ||||
| Pre-breeding/LH injection– | ||||
| Mean (μg/g) | 0.34 ± 0.03 | 0.31 ± 0.04 | 0.41 ± 0.16 | 0.26 ± 0.02 [33] 0.29 ± 0.01 [39] 0.33 ± 0.07 [25] |
| Baseline (μg/g) | 0.26 ± 0.02 | 0.26 ± 0.05 | 0.35 ± 0.14 | 0.21 ± 0.02 [33] 0.22 ± 0.01 [39] |
| Peak frequency (%) | 10.06 ± 2.22 | 14.08 ± 2.00 | 14.75 ± 3.40 | |
| Mean peak amplitude (μg/g) | 0.75 ± 0.35 2 | 0.22 ± 0.04 | 0.27 ± 0.16 | |
| Post-breeding/LH injection– | ||||
| Mean (μg/g) | 0.36 ± 0.02 | 0.36 ± 0.04 | 0.32 ± 0.06 | Preg: 0.32 ± 0.06 [25] NPLP: 0.28 ± 0.05 [25] |
| Baseline (μg/g) | 0.24 ± 0.01 | 0.28 ± 0.04 | 0.20 ± 0.02 | |
| Peak frequency (%) | 21.53 ± 2.03 | 15.00 ± 1.43 | 17.01 ± 2.28 | |
| Mean peak amplitude (μg/g) | 0.34 ± 0.03 | 0.29 ± 0.04 | 0.33 ± 0.10 | |
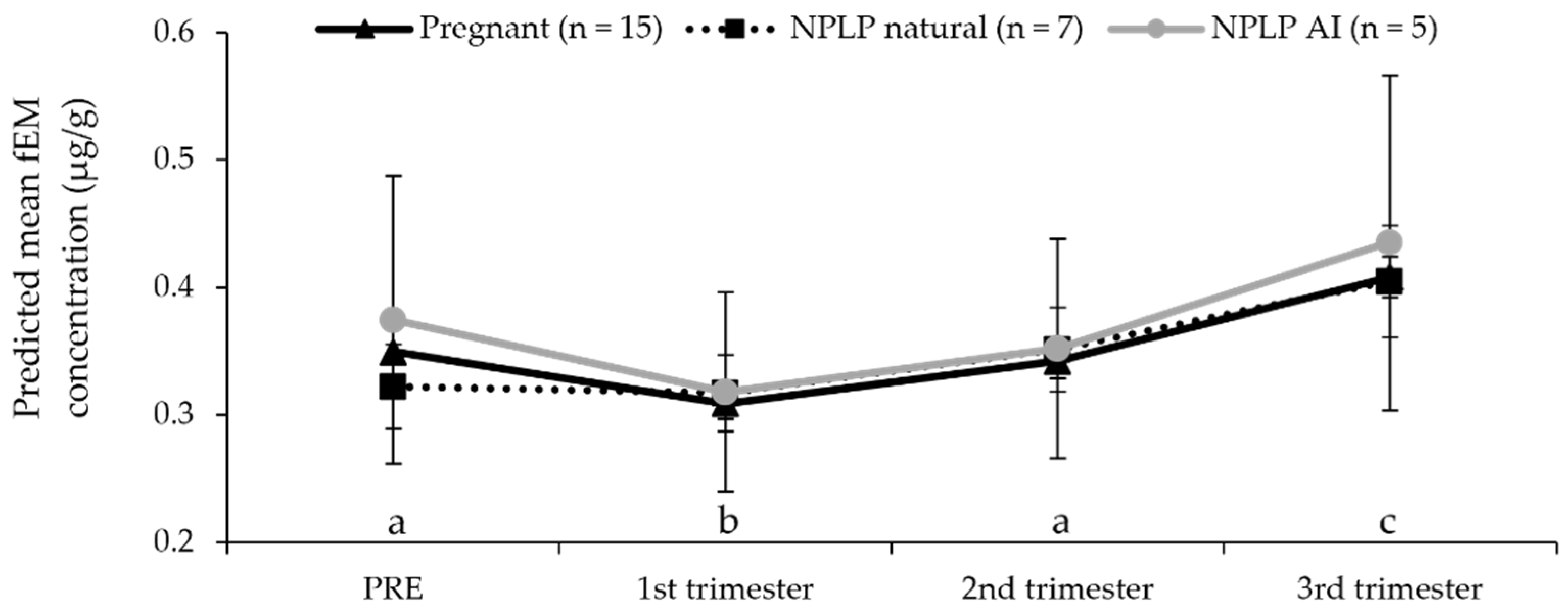
| First trimester mean (μg/g) | 0.29 ± 0.02 | 0.33 ± 0.04 | 0.36 ± 0.07 | |
| Second trimester mean (μg/g) | 0.37 ± 0.02 | 0.35 ± 0.04 | 0.29 ± 0.06 | |
| Third trimester mean (μg/g) | 0.44 ± 0.05 | 0.45 ± 0.05 | 0.32 ± 0.06 | |
| Fecal glucocorticoid metabolites: | ||||
| Pre-breeding/LH injection– | ||||
| Mean (μg/g) | 0.42 ± 0.05 | 0.48 ± 0.07 | 0.65 ± 0.07 | 0.98 ± 0.13 [33] 0.62 ± 0.07 [39] |
| Baseline (μg/g) | 0.25 ± 0.04 | 0.29 ± 0.02 | 0.40 ± 0.05 | 0.60 ± 0.12 [33] 0.32 ± 0.04 [39] |
| Peak frequency (%) | 12.25 ± 5.02 | 8.22 ± 3.78 | 14.10 ± 4.67 | |
| Mean peak amplitude (μg/g) | 0.47 ± 0.20 | 0.84 ± 0.40 | 1.63 ± 0.15 | |
| Post-breeding/LH injection– | ||||
| Mean (μg/g) | 0.53 ± 0.06 | 0.35 ± 0.05 | 0.61 ± 0.17 | |
| Baseline (μg/g) | 0.28 ± 0.03 | 0.26 ± 0.04 | 0.43 ± 0.19 | |
| Peak frequency (%) | 14.42 ± 2.99 | 4.60 ± 2.06 | 15.44 ± 4.19 | |
| Mean peak amplitude (μg/g) | 1.25 ± 0.20 | 0.48 ± 0.18 | 1.82 ± 0.71 | |
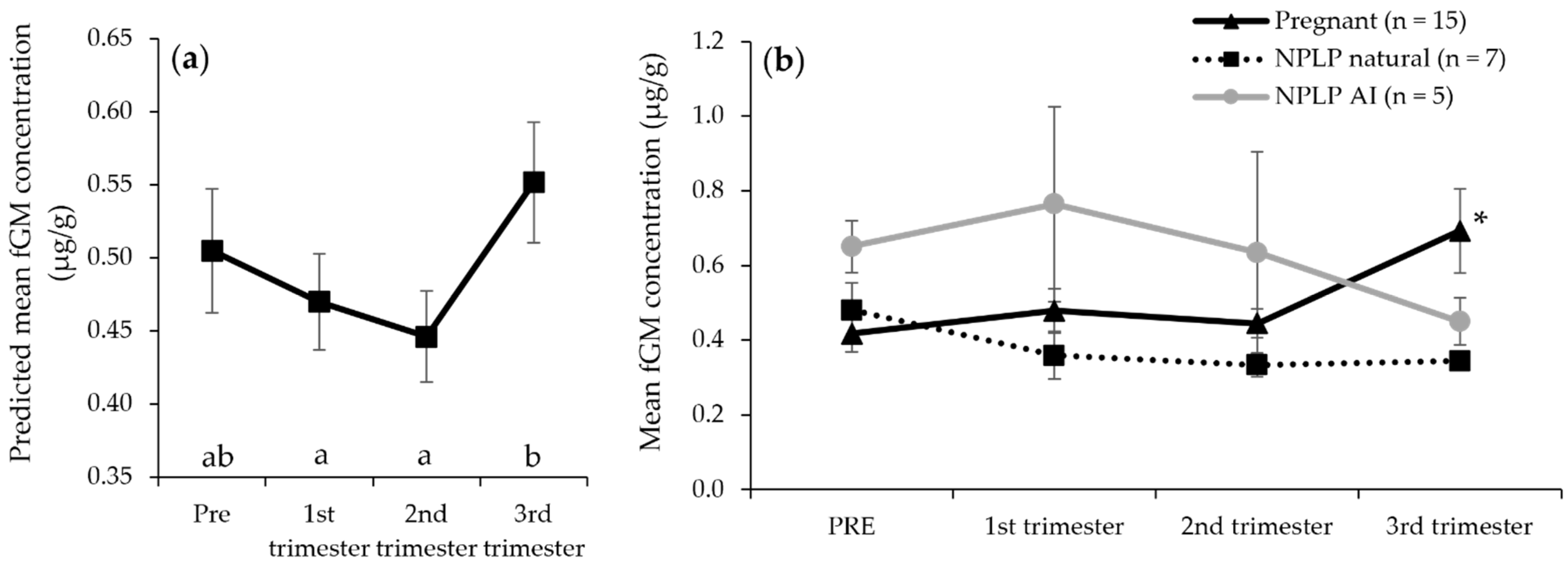
| First trimester mean (μg/g) | 0.48 ± 0.06 | 0.36 ± 0.06 | 0.77 ± 0.26 | |
| Second trimester mean (μg/g) | 0.45 ± 0.04 | 0.33 ± 0.03 | 0.64 ± 0.27 | |
| Third trimester mean (μg/g) | 0.69 ± 0.11 | 0.34 ± 0.02 | 0.45 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koester, D.C.; Maly, M.A.; Putman, S.; Edwards, K.L.; Meeks, K.; Crosier, A.E. An Investigation of Ovarian and Adrenal Hormone Activity in Post-Ovulatory Cheetahs (Acinonyx jubatus). Animals 2022, 12, 809. https://doi.org/10.3390/ani12070809
Koester DC, Maly MA, Putman S, Edwards KL, Meeks K, Crosier AE. An Investigation of Ovarian and Adrenal Hormone Activity in Post-Ovulatory Cheetahs (Acinonyx jubatus). Animals. 2022; 12(7):809. https://doi.org/10.3390/ani12070809
Chicago/Turabian StyleKoester, Diana C., Morgan A. Maly, Sarah Putman, Katie L. Edwards, Karen Meeks, and Adrienne E. Crosier. 2022. "An Investigation of Ovarian and Adrenal Hormone Activity in Post-Ovulatory Cheetahs (Acinonyx jubatus)" Animals 12, no. 7: 809. https://doi.org/10.3390/ani12070809
APA StyleKoester, D. C., Maly, M. A., Putman, S., Edwards, K. L., Meeks, K., & Crosier, A. E. (2022). An Investigation of Ovarian and Adrenal Hormone Activity in Post-Ovulatory Cheetahs (Acinonyx jubatus). Animals, 12(7), 809. https://doi.org/10.3390/ani12070809