
Effect of Fasting and Refeeding on Juvenile Leopard Mandarin Fish Siniperca scherzeri



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Rearing Condition
2.2. Experimental Design and Management
2.3. Analysis of Blood and Body Content
2.4. Growth Parameters and Statistical Analysis
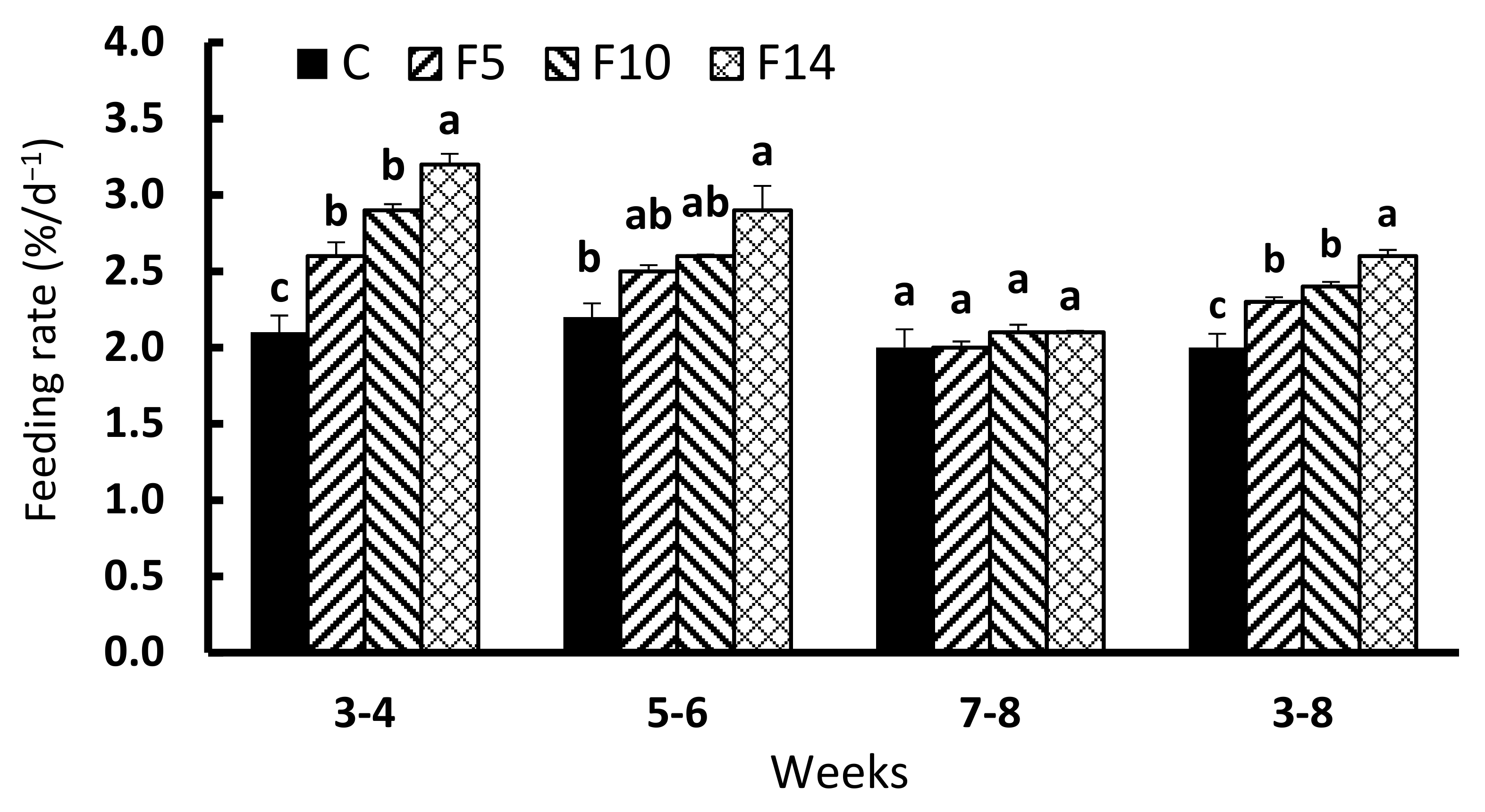
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilber, D.H.; Clarke, D.G. Biological Effects of Suspended Sediments: A Review of Suspended Sediment Impacts on Fish and Shellfish with Relation to Dredging Activities in Estuaries. N. Am. J. Fish. Manag. 2001, 21, 855–875. [Google Scholar] [CrossRef]
- Cho, S.H.; Lee, S.-M.; Park, B.H. Effect of feeding ratio on growth and body composition of juvenile olive flounder Paralichthys olivaceus fed extruded pellets during the summer season. Aquaculture 2006, 251, 78–84. [Google Scholar] [CrossRef]
- Oh, S.Y.; Noh, C.H.; Cho, S.Y. Effect of restricted feeding regimes on compensatory growth and body consumption of red sea bream, Pagrus major. J. World Aquac. Soc. 2007, 38, 443–449. [Google Scholar] [CrossRef]
- Urbinati, E.C.; Sarmiento, S.J.; Takahashi, L.S. Short-term cycles of feed deprivation and refeeding promote full compensatory growth in the Amazon fish matrinxa, Brycon amazonicus. Aquaculture 2014, 235, 273–283. [Google Scholar] [CrossRef]
- Oh, S.-Y.; Park, J. Effect of feed deprivation on compensatory growth in juvenile rock bream Oplegnathus fasciatus. Fish. Sci. 2019, 85, 813–819. [Google Scholar] [CrossRef]
- Assis, Y.P.A.S.; Porto, L.D.A.; Melo, N.; Palheta, G.D.A.; Luz, R.K.; Favero, G.C. Feed restriction as a feeding management strategy in Colossoma macropomum juveniles under recirculating aquaculture system (RAS). Aquaculture 2020, 529, 735689. [Google Scholar] [CrossRef]
- Ali, M.; Nicieza, A.; Wootton, R.J. Compensatory growth in fishes: A response to growth depression. Fish Fish. 2003, 4, 147–190. [Google Scholar] [CrossRef]
- Oh, S.Y.; Noh, C.H.; Kang, R.S. Compensatory growth and body consumption of juvenile black rockfish, Sebastes schlegeli following feed deprivation. Fish Sci. 2008, 74, 846–852. [Google Scholar] [CrossRef]
- Hayward, R.S.; Noltie, D.B.; Wang, N. Use of Compensatory Growth to Double Hybrid Sunfish Growth Rates. Trans. Am. Fish. Soc. 1997, 126, 316–322. [Google Scholar] [CrossRef]
- Jobling, M.; Meløy, O.; Dos Santos, J.; Christiansen, B. The compensatory growth response of the Atlantic cod: Effects of nutritional history. Aquac. Int. 1994, 2, 75–90. [Google Scholar] [CrossRef]
- de Macêdo, S.; de Almeida, O.C.; Lucena, J.E.C.; de Madeiro Torres, M.B.A.; de Alemeida Bicudo, J. Combined effects of dietary starch: Protein ratios and short cycles of fasting/refeeding on Nile tilapia growth and liver health. Aquac. Res. 2020, 52, 1139–1149. [Google Scholar] [CrossRef]
- Mattila, J.; Koskela, J.; Pirhonen, J. The effect of the length of repeated feed deprivation between single meals on compensatory growth of pikeperch Sander lucioperca. Aquaculture 2009, 296, 65–70. [Google Scholar] [CrossRef]
- Bolivar, R.B.; Jimenez, E.B.T.; Brown, C.L. Alternate-Day Feeding Strategy for Nile Tilapia Grow Out in the Philippines: Marginal Cost-Revenue Analyses. N. Am. J. Aquac. 2006, 68, 192–197. [Google Scholar] [CrossRef]
- Oh, S.-Y.; Kim, M.-S.; Kwon, J.Y.; Maran, B.A.V. Effects of feed restriction to enhance the profitable farming of blackhead seabream Acanthopagrus schlegelii schlegelii in sea cages. Ocean Sci. J. 2013, 48, 263–268. [Google Scholar] [CrossRef]
- Morshedi, V.; Kochanian, P.; Bahmani, M.; Yazdani, M.; Pourali, H.; Ashouri, G.; Pasha-Zanoosi, H. Cyclical short-term starvation and refeeding provokes compensatory growth in sub-yearling Siberian sturgeon, Acipenser baerii Brandt, 1869. Anim. Feed Sci. Technol. 2017, 232, 207–214. [Google Scholar] [CrossRef]
- Roa, F.G.B.; Silva, S.S.; Hoshiba, M.A.; Silva, L.K.S.; de Barros, A.F.; de Abreu, J.S. Production performance of tambaqui juveniles subjected to short feed-deprivation and refeeding cycles. Bol. Inst. Pesca 2019, 45, 1–9. [Google Scholar] [CrossRef]
- de Souza e Silva, W.; Hisano, H.; Mattioli, C.C.; Torres, I.F.A.; de Oliveira Paes-Leme, F.; Luz, R.K. Effects of cyclical short-term fasting and refeeding on juvenile Lophiosilurus alexandri, a carnivorous Neotropical catfish. Aquaculture 2019, 505, 12–17. [Google Scholar] [CrossRef]
- Zhou, C.W.; Yang, Q.; Cai, D.L. On the classification and distribution of the Sinipercinae fishes (Family Serranidae). Zool. Res. 1988, 9, 113–126. [Google Scholar]
- Zhang, L.; Wang, Y.J.; Hu, M.H.; Fan, Q.X.; Cheung, S.G.; Shin, P.K.S.; Li, H.; Cao, L. Effect of the timing of initial feeding on growth and survival of spotted mandarin fish Siniperca scherzeri larvae. J. Fish Biol. 2009, 75, 1158–1172. [Google Scholar] [CrossRef] [Green Version]
- Sankian, Z.; Khosravi, S.; Kim, Y.-O.; Lee, S.-M. Dietary protein requirement for juvenile mandarin fish, Siniperca scherzeri. J. World Aquac. Soc. 2018, 50, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.F.; Liu, J.K.; Huang, B.Y. The role of sense organs in the feeding behavior of Chinese perch. J. Fish Biol. 1998, 52, 1058–1067. [Google Scholar] [CrossRef]
- Su, S.Q.; Zhang, H.Q.; He, Z.Y.; Zhang, Z.X. A comparative study of the nutrients and amino acid composition of the muscle of Siniperca chuatsi and Siniperca scherzeri. J. SW Agricult. Univ. 2005, 27, 898–901. [Google Scholar]
- Bureau, D.P.; Hua, K.; Cho, C.Y. Effect of feeding level on growth and nutrient deposition in rainbow trout (Oncorhynchus mykiss Walbaum) growing from 150 to 600 g. Aquac. Res. 2006, 37, 1090–1098. [Google Scholar] [CrossRef]
- Yuan, Y.-C.; Yang, H.-J.; Gong, S.-Y.; Luo, Z.; Yuan, H.-W.; Chen, X.-K. Effects of feeding levels on growth performance, feed utilization, body composition and apparent digestibility coefficients of nutrients for juvenile Chinese sucker, Myxocyprinus asiaticus. Aquac. Res. 2009, 41, 1030–1042. [Google Scholar] [CrossRef]
- Lee, S.-M.; Cho, S.H.; Kim, D.-J. Effects of feeding frequency and dietary energy level on growth and body composition of juvenile flounder, Paralichthys olivaceus (Temminck & Schlegel). Aquac. Res. 2000, 31, 917–921. [Google Scholar] [CrossRef]
- Kim, Y.O.; Oh, S.Y.; Kim, T.W. Effects of the feeding rate on growth performance, body composition, and hematological properties of juvenile mandarin Siniperca scherzeri in recirculating aquaculture system. Sustainability 2021, 13, 8257. [Google Scholar] [CrossRef]
- Sankian, Z.; Khosravi, S.; Kim, Y.O.; Lee, S.M. Effect of dietary protein and lipid level on growth, feed utilization and muscle composition in golden mandarin fish, Siniperca scherzeri. Fish Aquat. Sci. 2017, 20, 7. [Google Scholar] [CrossRef]
- Sankian, Z.; Khosravi, S.; Kim, Y.O.; Lee, S.M. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, plasma biochemical indices, selected immune parameters and antioxidant enzyme activities of mandarin fish Siniperca scherzeri juveniles. Aquaculture 2018, 496, 79–87. [Google Scholar]
- Sankian, Z.; Khosravi, S.; Kim, Y.-O.; Lee, S.-M. Total replacement of dietary fish oil with alternative lipid sources in a practical diet for mandarin fish, Siniperca scherzeri, juveniles. Fish. Aquat. Sci. 2019, 22, 8. [Google Scholar] [CrossRef]
- Mo, A.J.; Sun, J.X.; Wang, Y.H.; Yang, K.; Yang, H.S.; Yuan, Y.C. Apparent digestibility of protein, energy and amino acids in nine protein sources at two content levels for mandarin fish, Siniperca chuatsi. Aquaculture 2018, 499, 42–50. [Google Scholar] [CrossRef]
- Kim, Y.O.; Oh, S.Y.; Lee, S.M. Influence of different feeding frequency on the growth and body composition of juvenile man-darin fish Siniperca scherzeri reared in a recirculating aquaculture system (RAS). Korean Soc. Fish Aquatic. Sci. 2020, 53, 538–543. [Google Scholar]
- Kim, Y.O.; Oh, S.Y.; Lee, W.S. Feeding ratio affects growth, body composition, and blood chemistry of mandarin (Siniperca scherzeri) in recirculating aquaculture system. Fish Aquat. Sci. 2021, 24, 219–227. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1995. [Google Scholar]
- Oh, S.-Y.; Kang, R.-S.; Myoung, J.-G.; Kim, C.-K.; Park, J.; Daniels, H.V. Effect of Ration Size Restriction on Compensatory Growth and Proximate Composition of Dark-banded Rockfish, Sebastes inermis. J. World Aquac. Soc. 2010, 41, 923–930. [Google Scholar] [CrossRef]
- Yokoyama, H.; Takashi, T.; Ishihi, Y.; Abo, K. Effects of restricted feeding on growth of red sea bream and sedimentation of aquaculture wastes. Aquaculture 2009, 286, 80–88. [Google Scholar] [CrossRef]
- Yilmaz, H.A.; Eroldogan, O.T. Combined effects of cycled starvation and feeding frequency on growth and oxygen consump-tion of gilthead sea bream, Sparus aurata. J. World Aquac. Soc. 2011, 42, 522–529. [Google Scholar] [CrossRef]
- Reigh, R.C.; Williams, M.B.; Jacob, B.J. Influence of repetitive periods of fasting and satiation feeding on growth and production characteristics of channel catfish, Ictalurus punctatus. Aquaculture 2006, 254, 506–516. [Google Scholar] [CrossRef]
- Mohseni, M.; Pourkazemi, M.; Hosseni, M.R.; Hassani, M.H.S.; Bai, S.C. Effects of the dietary protein levels and the protein to energy ratio in sub-yearling Persian sturgeon, Acipenser persicus (Borodin). Aquac. Res. 2011, 44, 378–387. [Google Scholar] [CrossRef]
- Jafari, N.; Falahatkar, B.; Sajjadi, M.M. Growth performance and plasma metabolites in juvenile Siberian sturgeon Acipenser baerii (Brandt, 1869) subjected to various feeding strategies at different sizes. Fish Physiol. Biochem. 2018, 44, 1363–1374. [Google Scholar] [CrossRef]
- Zhu, X.; Xie, S.; Lei, W.; Cui, Y.; Yang, Y.; Wootton, R. Compensatory growth in the Chinese longsnout catfish, Leiocassis longirostris following feed deprivation: Temporal patterns in growth, nutrient deposition, feed intake and body composition. Aquaculture 2005, 248, 307–314. [Google Scholar] [CrossRef]
- Xie, S.; Zhu, X.; Cui, Y.; Wootton, R.J.; Lei, W.; Yang, Y. Compensatory growth in gibel carp following feed deprivation: Temporal patterns in growth, nutrient deposition, feed intake and body composition. J. Fish Biol. 2001, 58, 999–1009. [Google Scholar] [CrossRef]
- Peres, H.; Satos, S.; Oliva-Teles, A. Lake of compensatory growth response in gilthead seabream, Sparus aurata juvenile following starvation and subsequent refeeding. Aquaculture 2011, 318, 384–388. [Google Scholar] [CrossRef]
- Tian, X.; Qin, J.G. A single phase of food deprivation provoked compensatory growth in barramundi, Lates calcarifer. Aquaculture 2003, 224, 169–179. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, Y.; Yang, Y.; Cai, F. Compensatory growth in hybrid tilapia, Oreochromis mossambicus × O. niloticus, reared in seawater. Aquaculture 2000, 189, 101–108. [Google Scholar] [CrossRef]
- Sevgili, H.; Hossu, B.; Emre, Y.; Kanyilmaz, M. Effect of various lengths of single-phase starvation on compensatory growth in rainbow trout under summer conditions (Oncorhynchus mykiss). Turk. J. Fish Aquat. Sci. 2013, 13, 465–477. [Google Scholar] [CrossRef]
- Henken, A.; Kleingeld, D.; Tijssen, P. The effect of feeding level on apparent digestibility of dietary dry matter, crude protein and gross energy in the African catfish Clarias gariepinus (Burchell, 1822). Aquaculture 1985, 51, 1–11. [Google Scholar] [CrossRef]
- Baloi, M.; Sterzelecki, F.; Sugai, J.; Passini, G.; Carvalho, C.; Cerqueira, V. Growth performance, body composition and metabolic response to feeding rates in juvenile Brazilian sardine Sardinella brasiliensis. Aquac. Nutr. 2017, 23, 1458–1466. [Google Scholar] [CrossRef]
- Russel, N.R.; Wotton, R.J. Appetite and growth compensation in the European minnow, Phoxinus (Cyprinidae), following short periods of food restriction. Environ. Biol. Fish 1992, 34, 277–285. [Google Scholar] [CrossRef]
- Qian, X.; Cui, Y.; Xiong, B.; Yang, Y. Compensatory growth, feed utilization and activity in gibel carp, following feed deprivation. J. Fish Biol. 2000, 56, 228–232. [Google Scholar] [CrossRef]
- Foss, A.; Imsland, A.K.; Vikingstad, E.; Stefansson, S.O.; Norberg, B.; Pedersen, S.; Sandvik, T.; Roth, B. Compensatory growth in Atlantic halibut: Effect of starvation and subsequent feeding on growth, maturation, feed utilization and flesh quality. Aquaculture 2009, 290, 304–310. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, Y.; Yang, Y.; Cai, F. Partial compensatory growth in hybrid tilapia Oreochromis mossambicus × O. niloticus following food deprivation. J. Appl. Ichthyol. 2005, 21, 389–393. [Google Scholar] [CrossRef]
- Blanquet, I.; Oliva-Teles, A. Effect of feed restriction on the growth performance of turbot (Scophthalmus maximus L.) juvenile under commercial rearing conditions. Aquac. Res. 2010, 41, 1255–1260. [Google Scholar]
- Känkänen, M.; Pirhonen, J. The effect of intermittent feeding on feed intake and compensatory growth of whitefish Coregonus lavaretus L. Aquaculture 2009, 288, 92–97. [Google Scholar] [CrossRef]
- Power, D.M.; Melo, J.; Santos, C.R.A. The effect of food deprivation and refeeding on the liver, thyroid hormones and trans-thyretin in sea bream. J. Fish Biol. 2000, 56, 374–387. [Google Scholar] [CrossRef]
- Peres, H.; Oliva-Teles, A. Protein and Energy Metabolism of European Seabass (Dicentrarchus labrax) Juveniles and Estimation of Maintenance Requirements. Fish Physiol. Biochem. 2005, 31, 23–31. [Google Scholar] [CrossRef]
- Jobling, M.; Johansen, S.J.S. The lipostat, hyperphagia and catch-up growth. Aquac. Res. 1999, 30, 473–478. [Google Scholar] [CrossRef]
- Johansen, S.J.S.; Ekli, M.; Stangnes, B.; Jobling, M. Weight gain and lipid deposition in Atlantic salmon, Salmo salar, during compensatory growth: Evidence for lipostatic regulation? Aquac. Res. 2001, 32, 963–974. [Google Scholar] [CrossRef]
- Gao, X.; Hong, L.; Liu, Z.; Guo, Z.; Wang, Y.; Lei, J. Body composition and compensatory growth in Nile tilapia Oreochromis niloticus under different feeding intervals. Chin. J. Oceanol. Limnol. 2015, 33, 945–956. [Google Scholar] [CrossRef]
- Tian, X.L.; Qin, R.G. Effects of previous ration restriction on compensatory growth in barramundi Lates calcarifer. Aquaculture 2004, 235, 273–283. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Barbieri, G.; Verani, J.R.; Hahn, N.S. Variação do fator de condição e do índice hepatossomático e suas relações com o ciclo reprodutivo em Rhinelepis aspera (Agassis, 1829) (Osteichthyes, Loricariidae) no rio Paranapanema, Porecatu, PR. Cienc. Cult. 1990, 42, 711–714. [Google Scholar]
- Cyrino, J.E.P.; Portz, L.; Martino, R.C. Protein and energy retention by juvenile largemouth bass Micropterus salmoides. Sci. Agric. 2000, 57, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Torstensen, B.E.; Tocher, D.R. The effects of fish oil replacement on lipid metabolism of fish. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G.M., Ng, W.K., Tocher, D.R., Eds.; Taylor & Francis: Abingdon, UK, 2011; pp. 405–437. [Google Scholar]
- McCue, M.D. Starvation physiology: Reviewing the different strategies animals use to survive a common challenge. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Jobling, M. Some observations on the effects of feeding frequency on the food intake and growth of plaice, Pleuronectes platessa L. J. Fish Biol. 1982, 20, 431–444. [Google Scholar] [CrossRef]
- Rehulka, J.; MinaRik, B.; Rehulkova, E. Red blood cell indices of rainbow trout Oncorhynchus mykiss (Walbaum) in aquaculture. Aquac. Res. 2004, 35, 529–546. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Khattab, Y.A.E.; Ahmad, M.H.; Shalaby, A.M.E. Compensatory growth, feed utilization, whole-body composition, and hematological changes in starved juvenile Nile tilapia (Oreochromis niloticus L.). J. Appl. Aquac. 2006, 18, 17–36. [Google Scholar] [CrossRef]
- Cho, Y.J.; Cho, S.H. Compensatory growth of olive flounder, Paralichthys olivaceus, fed extruded pellet with different feeding regimes. J. World Aquac. Soc. 2009, 40, 505–512. [Google Scholar] [CrossRef]
- Caruso, G.; Maricchiolo, G.; Micale, V.; Genovese, L.; Caruso, R.; Denaro, M.G. Physiological responses to starvation in the European eel (Anguilla anguilla): Effects on hematological, biochemical, non-specific immune parameters and skin structures. Fish Physiol. Biochem. 2010, 36, 71–83. [Google Scholar] [CrossRef]
- Caruso, G.; Denaro, M.G.; Caruso, R.; Genovese, L.; Mancari, F.; Maricchiolo, G. Short fasting and refeeding in red porgy (Pagrus pagrus, Linnaeus 1758): Response of some haematological, biochemical and non specific immune parameters. Mar. Environ. Res. 2012, 81, 18–25. [Google Scholar] [CrossRef]
- Falahatkar, B. The metabolic effects of feeding and fasting in beluga Huso huso. Mar. Environ. Res. 2012, 82, 69–75. [Google Scholar] [CrossRef]
- Chatzifotis, S.; Papadaki, M.; Despoti, S.; Roufidou, C.; Antonopoulou, E. Effect of starvation and re-feeding on reproductive indices, body weight, plasma metabolites and oxidative enzyme of sea bass (Dicentrachus labrax). Aquaculture 2011, 316, 53–59. [Google Scholar] [CrossRef]
- Favero, G.C.; Costa dos Santos, F.A.; Soares da Costa, G.J.; Cortezzi Pedras, P.P.; Lima Ferreira, A.; De Souza e Silva, W.; Soares Ferreira, N.; Neves, L.D.C.; Kennedy , R.L. Effects of short feed restriction cycles in Piaractus brachypomus juveniles. Aquaculture 2021, 536, 736465. [Google Scholar] [CrossRef]
- Heming, T.A.; Paleczny, E.J. Compositional changes in skin mucus and blood serum during starvation of trout. Aquaculture 1987, 66, 265–273. [Google Scholar] [CrossRef]
- Regost, C.; Arzel, J.; Cardinal, M.; Laroche, M.; Kaushik, S. Fat deposition and flesh quality in seawater reared, triploid brown trout (Salmo trutta) as affected by dietary fat levels and starvation. Aquaculture 2001, 193, 325–345. [Google Scholar] [CrossRef] [Green Version]
- Furné, M.; Morales, A.E.; Trenzado, C.E.; García-Gallego, M.; Hidalgo, M.C.; Domezain, A.; Rus, A.S. The metabolic effects of prolonged starvation and refeeding in sturgeon and rainbow trout. J. Comp. Physiol. B 2011, 182, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.C.; Gimbo, R.; Montoya, L.N.F.; Zanuzzo, F.S.; Urbinati, E.C. Fasting and refeeding lead to more efficient growth in lean pacu (Piaractus mesopotamicus). Aquac. Res. 2018, 49, 359–366. [Google Scholar] [CrossRef]
- Godavarthy, P.; Kumari, Y.S.; Bikshapathy, E. Starvation induced cholesterogenesis in hepatic and extra hepatic tissues of climbing Perch, Anabas testudineus (Bloch). Saudi J. Biol. Sci. 2012, 19, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | CFi (%) 1 | CFf (%) 2 | CVBLi (%) 3 | CVBLf (%) 4 | CVBWi (%) 5 | CVBWf (%) 6 |
|---|---|---|---|---|---|---|
| C | 1.07 ± 0.07 | 1.07 ± 0.03 | 13.6 ± 0.20 | 9.0 ± 0.4 | 18.7 ± 0.86 | 27.7 ± 1.13 |
| F5 | 1.05 ± 0.01 | 1.09 ± 0.03 | 13.4 ± 0.19 | 8.1 ± 1.1 | 13.7 ± 1.24 | 27.2 ± 1.02 |
| F10 | 1.06 ± 0.02 | 1.07 ± 0.03 | 13.7 ± 0.16 | 7.8 ± 0.9 | 15.3 ± 1.81 | 27.2 ± 0.81 |
| F14 | 1.07 ± 0.01 | 1.10 ± 0.03 | 13.6 ± 0.58 | 5.6 ± 0.7 | 16.0 ± 0.76 | 27.5 ± 0.52 |
| Periods | Variables | Treatment | |||
|---|---|---|---|---|---|
| C | F5 | F10 | F14 | ||
| Week 2 (n = 2) | Moisture (%) | 70.6 ± 0.9 b | 73.5 ± 0.6 ab | 74.8 ± 0.2 ab | 77.3 ± 1.2 a |
| Crude protein (%) | 19.9 ± 0.8 | 18.3 ± 0.3 | 20.0 ± 0.2 | 19.9 ± 0.9 | |
| Crude lipid (%) | 4.7 ± 0.05 a | 3.7 ± 0.05 b | 3.5 ± 0.05 b | 2.9 ± 0.15 c | |
| Ash (%) | 6.1 ± 0.1 | 6.0 ± 0.1 | 6.1 ± 0.1 | 6.1 ± 0.1 | |
| HSI (%) 1 | 2.5 ± 0.08 a | 1.7 ± 0.37 ab | 1.4 ± 0.14 b | 1.3 ± 0.06 b | |
| VSI (%) 2 | 7.7 ± 0.40 a | 5.0 ± 0.22 b | 3.9 ± 0.55 c | 3.5 ± 0.14 c | |
| Lipid/LBM 3 | 0.18 ± 0.01 a | 0.15 ± 0.01 b | 0.13 ± 0.01 bc | 0.10 ± 0.01 c | |
| Week 8 (n = 3) | Moisture (%) | 69.5 ± 0.4 | 71.8 ± 0.4 | 72.2 ± 0.4 | 72.0 ± 0.2 |
| Crude protein (%) | 19.5 ± 0.7 | 19.5 ± 0.3 | 19.8 ± 0.2 | 19.7 ± 0.5 | |
| Crude lipid (%) | 3.7 ± 0.2 | 3.6 ± 0.4 | 3.4 ± 0.3 | 3.6 ± 0.3 | |
| Ash (%) | 6.5 ± 0.2 | 6.0 ± 0.3 | 5.6 ± 0.2 | 5.9 ± 0.1 | |
| HSI (%) 1 | 1.3 ± 0.06 b | 1.6 ± 0.08 a | 1.7 ± 0.09 a | 1.7 ± 0.02 a | |
| VSI (%) 2 | 4.6 ± 0.06 | 4.5 ± 0.28 | 4.6 ± 0.19 | 4.8 ± 0.21 | |
| Lipid/LBM 3 | 0.14 ± 0.01 | 0.14 ± 0.02 | 0.13 ± 0.01 | 0.14 ± 0.01 | |
| Periods | C | F5 | F10 | F14 | |
|---|---|---|---|---|---|
| Week 2 (n = 2) | Hb (g dL−1) 1 | 7.8 ± 0.28 | 8.0 ± 0.36 | 7.7 ± 0.17 | 7.8 ± 0.15 |
| GLU (mg dL−1) 2 | 150.3 ± 12.7 b | 141.0 ± 14.2 b | 177.3 ± 14.3 ab | 232.9 ± 17.2 a | |
| TCHO (mg dL−1) 3 | 170.4 ± 3.55 a | 169.7 ± 4.07 a | 159.9 ± 3.25 ab | 150.0 ± 2.82 b | |
| GOT (U L−1) 4 | 395.1 ± 23.7 | 387.1 ± 14.2 | 375.5 ± 18.2 | 333.3 ± 10.6 | |
| GPT (U L−1) 5 | 129.2 ± 5.2 | 143.9 ± 6.6 | 150.1 ± 7.0 | 139.6 ± 6.0 | |
| HDLC (U L−1) 6 | 108.2 ± 1.60 | 109.1 ± 0.77 | 107.9 ± 1.51 | 108.1 ± 0.94 | |
| Week 8 (n = 3) | Hb (g dL−1) 1 | 7.3 ± 0.9 | 7.5 ± 0.6 | 6.6 ± 0.6 | 6.7 ± 0.5 |
| GLU (mg dL−1) 2 | 138.0 ± 10.1 | 141.0 ± 8.5 | 139.7 ± 6.36 | 138.3 ± 7.33 | |
| TCHO (mg dL−1) 3 | 143.3 ± 8.5 | 143.8 ± 3.3 | 137.0 ± 1.7 | 143.0 ± 8.3 | |
| GOT (U L−1) 4 | 226.8 ± 14.0 | 249.0 ± 15.1 | 274.5 ± 11.6 | 223.3 ± 9.5 | |
| GPT (U L−1) 5 | 71.0 ± 8.6 | 64.6 ± 5.4 | 53.5 ± 2.5 | 44.3 ± 2.7 | |
| HDLC (U L−1) 6 | 98.5 ± 5.1 | 99.8 ± 2.7 | 98.0 ± 0.9 | 98.3 ± 4.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-O.; Oh, S.-Y.; Kim, T. Effect of Fasting and Refeeding on Juvenile Leopard Mandarin Fish Siniperca scherzeri. Animals 2022, 12, 889. https://doi.org/10.3390/ani12070889
Kim Y-O, Oh S-Y, Kim T. Effect of Fasting and Refeeding on Juvenile Leopard Mandarin Fish Siniperca scherzeri. Animals. 2022; 12(7):889. https://doi.org/10.3390/ani12070889
Chicago/Turabian StyleKim, Yi-Oh, Sung-Yong Oh, and Taewon Kim. 2022. "Effect of Fasting and Refeeding on Juvenile Leopard Mandarin Fish Siniperca scherzeri" Animals 12, no. 7: 889. https://doi.org/10.3390/ani12070889
APA StyleKim, Y. -O., Oh, S. -Y., & Kim, T. (2022). Effect of Fasting and Refeeding on Juvenile Leopard Mandarin Fish Siniperca scherzeri. Animals, 12(7), 889. https://doi.org/10.3390/ani12070889