
Effects of Intravenous Antimicrobial Drugs on the Equine Fecal Microbiome

 , , and
, , and 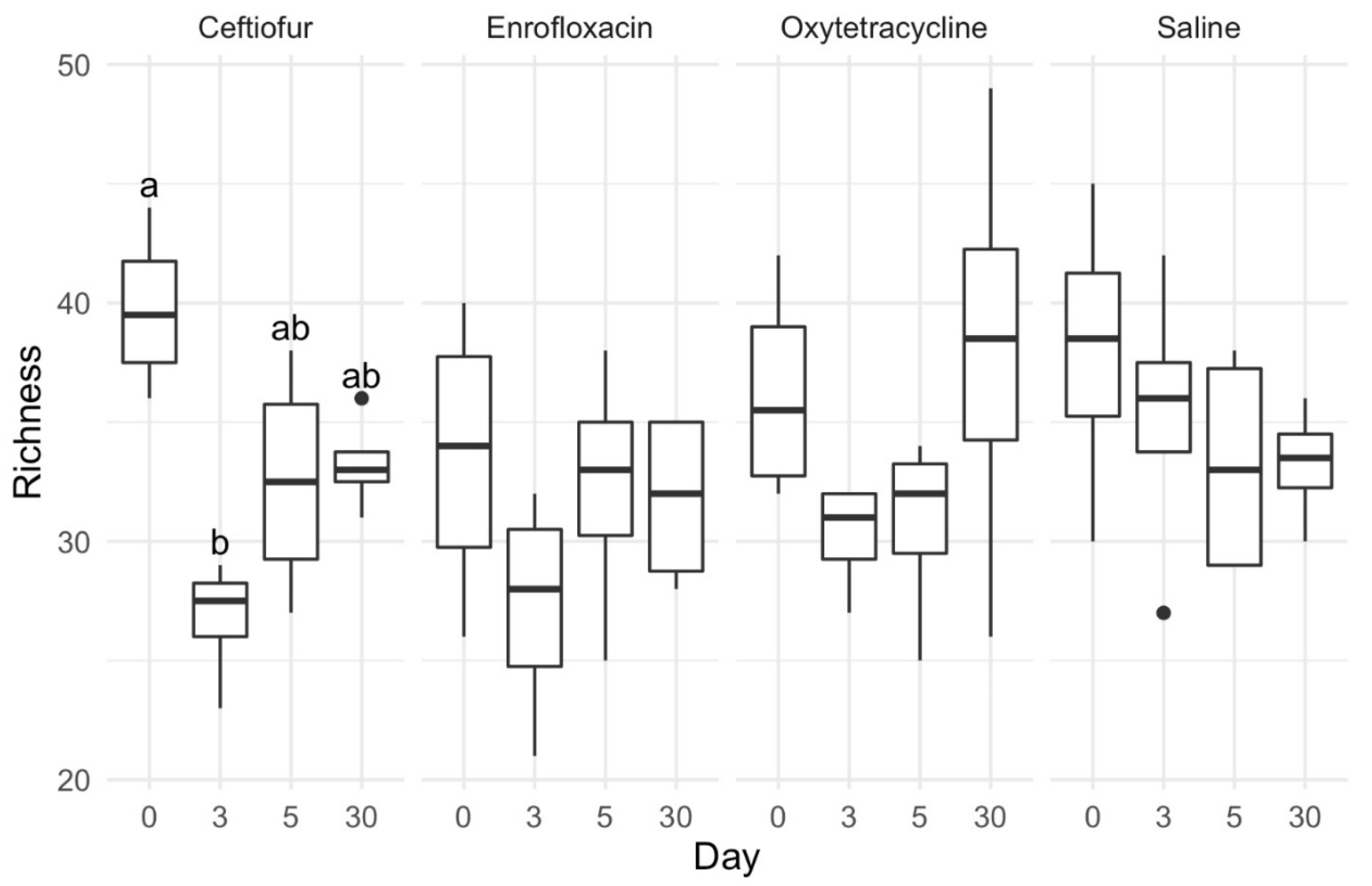{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experiment
2.3. DNA Extraction, PCR Amplification and Sequencing
3. Data Analysis
4. Results
4.1. Clinical Response to Antimicrobial Therapy
4.2. Overall Assessment of the Sequences
4.3. Alpha Diversity
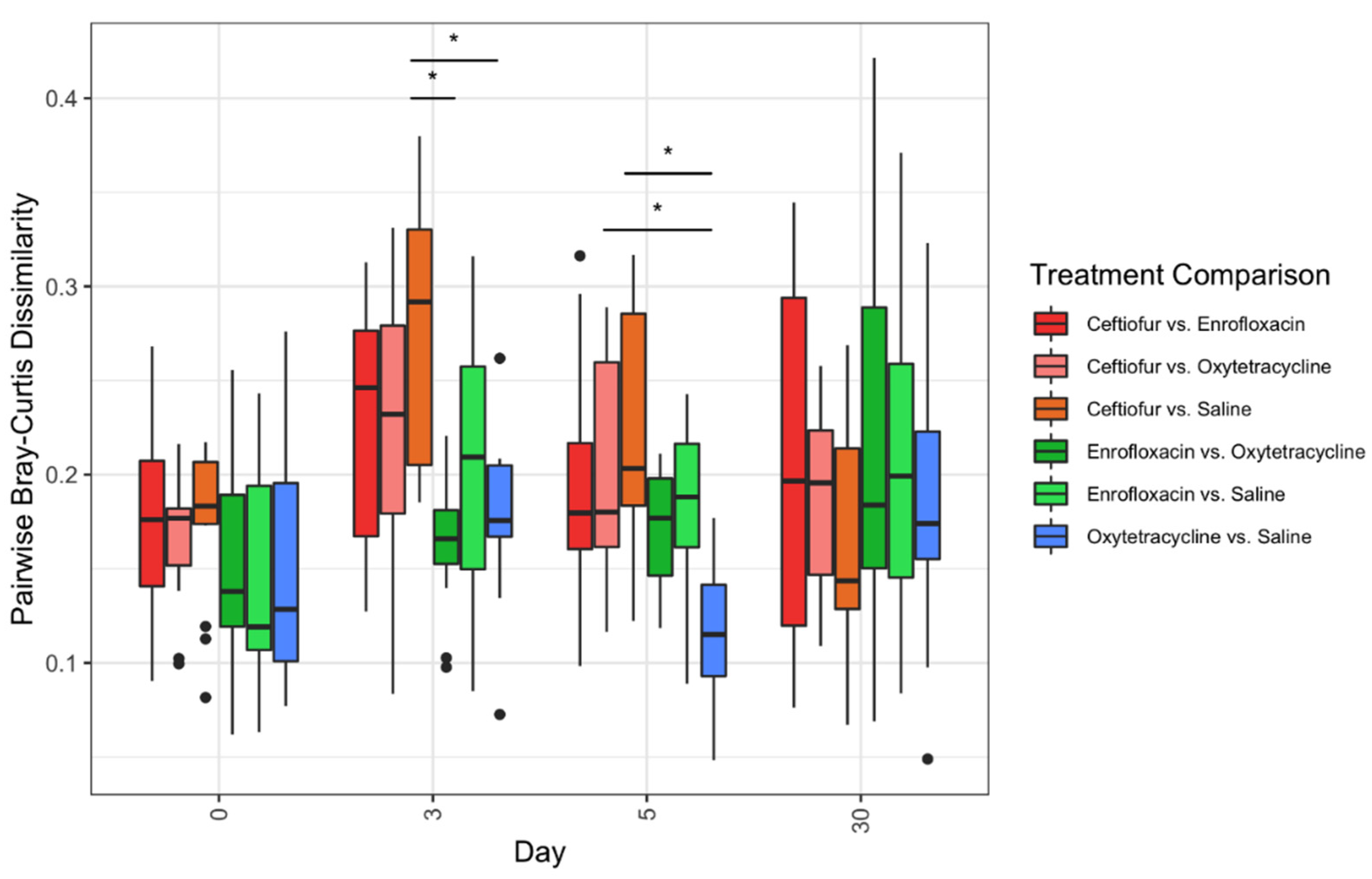
4.4. Beta Diversity
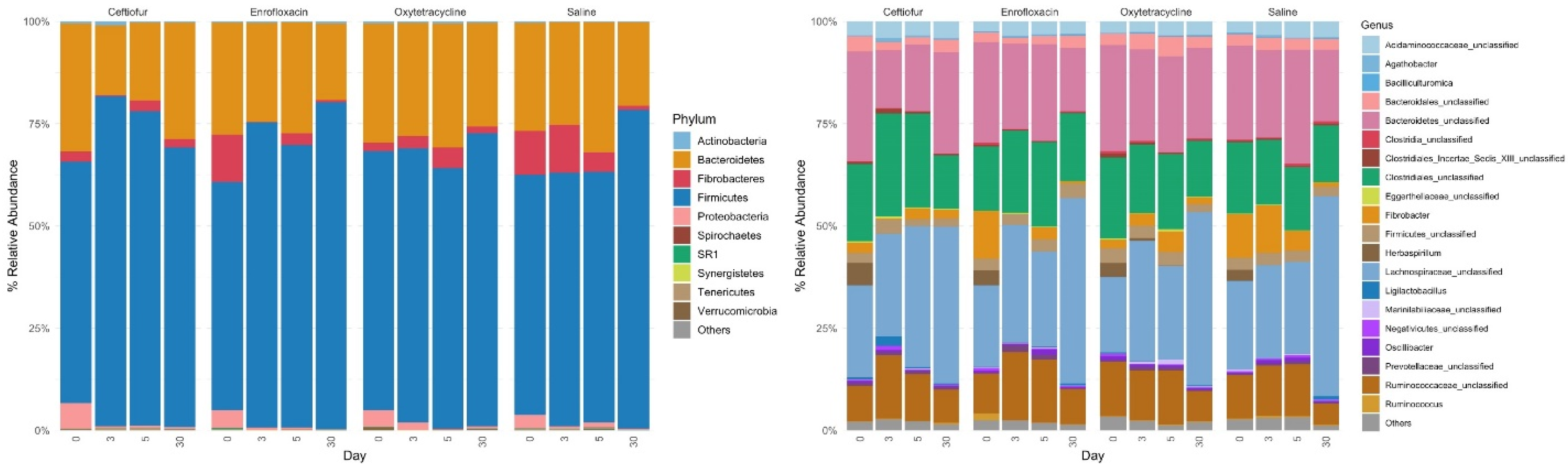
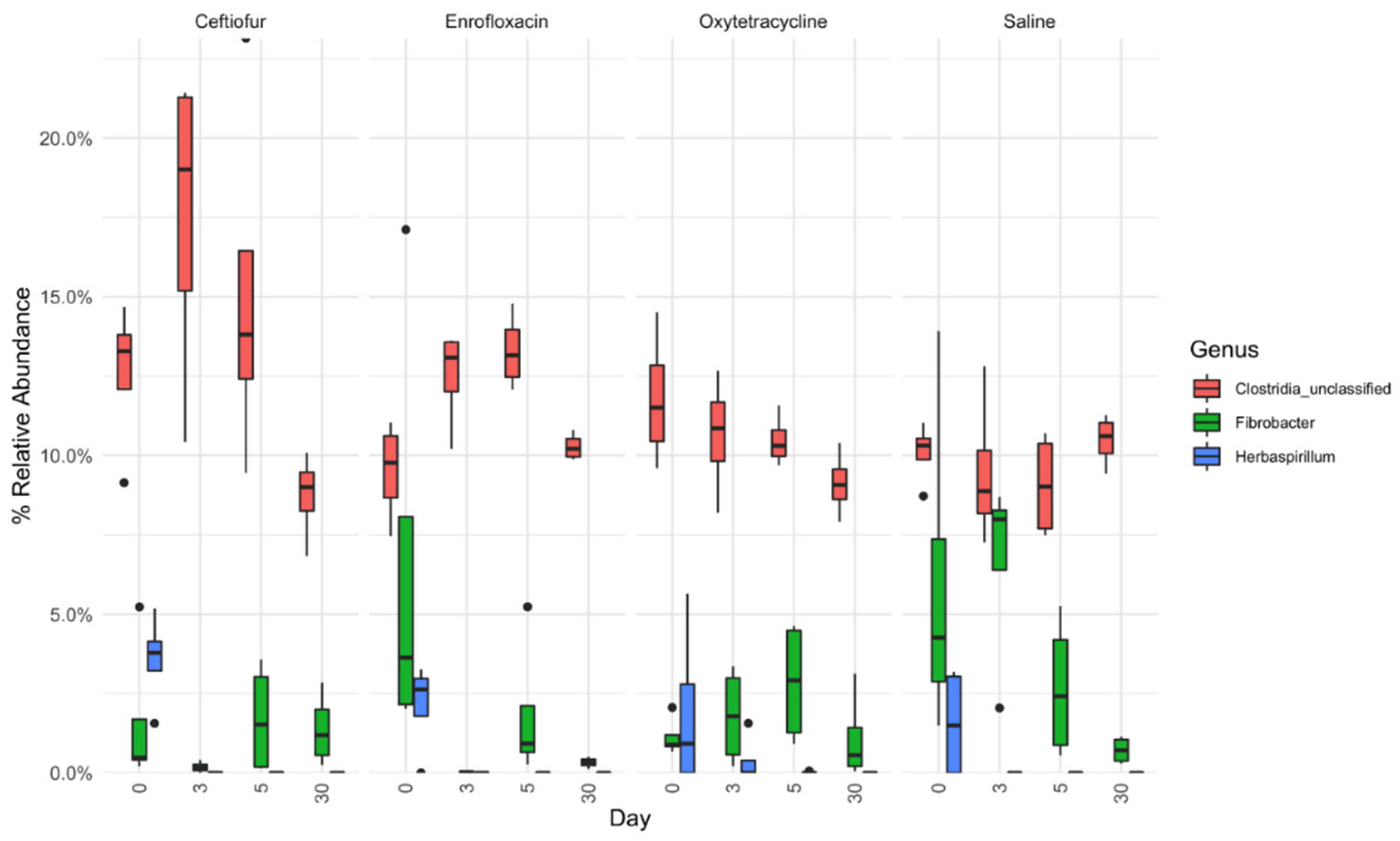
4.5. Relative Abundance
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Båverud, V. Clostridium difficile infections in animals with special reference to the horse. A review. Vet. Q. 2002, 24, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Båverud, V.; Franklin, A.; Gunnarsson, A.; Gustafsson, A.; Hellander-Edman, A. Clostridium difficile associated with acute colitis in mares when their foals are treated with erythromycin and rifampicin for rhodococcus equi pneumonia. Equine Vet. J. 1998, 30, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Båverud, V.; Gustafsson, A.; Franklin, A.; Lindholm, A.; Gunnarsson, A. Clostridium difficile associated with acute colitis in mature horses treated with antibiotics. Equine Vet. J. 1997, 29, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Barr, B.S.; Waldridge, B.M.; Morresey, P.R.; Reed, S.M.; Clark, C.; Belgrave, R.; Donecker, J.M.; Weigel, D.J. Antimicrobial-associated diarrhoea in three equine referral practices. Equine Vet. J. 2013, 45, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Stratton-Phelps, M.; Wilson, W.D.; Gardner, I.A. Risk of adverse effects in pneumonic foals treated with erythromycin versus other antibiotics: 143 cases (1986–1996). J. Am. Vet. Med. Assoc. 2000, 217, 68–73. [Google Scholar] [CrossRef]
- Abad, C.L.R.; Safdar, N. A review of clostridioides difficile infection and antibiotic-associated diarrhea. Gastroenterol. Clin. N. Am. 2021, 50, 323–340. [Google Scholar] [CrossRef]
- McGorum, B.C.; Pirie, R.S. Antimicrobial associated diarrhoea in the horse. Part 1: Overview, pathogenesis and risk factors. Equine Vet. Educ. 2010, 11, 369–373. [Google Scholar] [CrossRef]
- Ross, C.L.; Spinler, J.K.; Savidge, T.C. Structural and functional changes within the gut microbiota and susceptibility to clostridium difficile infection. Anaerobe 2016, 41, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Keeney, K.M.; Yurist-Doutsch, S.; Arrieta, M.-C.; Finlay, B.B. Effects of antibiotics on human microbiota and subsequent disease. Annu. Rev. Microbiol. 2014, 68, 217–235. [Google Scholar] [CrossRef]
- Kim, S.; Covington, A.; Pamer, E.G. The intestinal microbiota: Antibiotics, colonization resistance, and enteric pathogens. Immunol. Rev. 2017, 279, 90–105. [Google Scholar] [CrossRef]
- Dethlefsen, L.; Relman, D.A. Incomplete recovery and individualized responses of the human distal gut microbiota to Repeated antibiotic perturbation. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4554–4561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foditsch, C.; Pereira, R.V.V.; Siler, J.D.; Altier, C.; Warnick, L.D. Effects of treatment with enrofloxacin or tulathromycin on fecal microbiota composition and genetic function of dairy calves. PLoS ONE 2019, 14, e0219635. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.C.; Stämpfli, H.R.; Arroyo, L.G.; Allen-Vercoe, E.; Gomes, R.G.; Weese, J.S. Changes in the equine fecal microbiota associated with the use of systemic antimicrobial drugs. BMC Vet. Res. 2015, 11, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collinet, A.; Grimm, P.; Julliand, S.; Julliand, V. Multidimensional approach for investigating the effects of an antibiotic-probiotic combination on the equine hindgut ecosystem and microbial fibrolysis. Front. Microbiol. 2021, 12, 646294. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, R.; Arroyo, L.G.; Leclere, M.; Costa, M.C. Species-level gut microbiota analysis after antibiotic-induced dysbiosis in horses. Animals 2021, 11, 2859. [Google Scholar] [CrossRef] [PubMed]
- Arnold, C.; Pilla, R.; Chaffin, K.; Lidbury, J.; Steiner, J.; Suchodolski, J. Alterations in the fecal microbiome and metabolome of horses with antimicrobial-associated diarrhea compared to antibiotic-treated and non-treated healthy case controls. Animals 2021, 11, 1807. [Google Scholar] [CrossRef]
- Arnold, C.E.; Isaiah, A.; Pilla, R.; Lidbury, J.; Coverdale, J.S.; Callaway, T.R.; Lawhon, S.D.; Steiner, J.; Suchodolski, J.S. The cecal and fecal microbiomes and metabolomes of horses before and after metronidazole administration. PLoS ONE 2020, 15, e0232905. [Google Scholar] [CrossRef]
- Sun, Y.; Wolcott, R.D.; Dowd, S.E. Tag-encoded FLX amplicon pyrosequencing for the elucidation of microbial and functional gene diversity in any environment. Methods Mol. Biol. 2011, 733, 129–141. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, L.G.; Rossi, L.; Santos, B.P.; Gomez, D.E.; Surette, M.G.; Costa, M.C. Luminal and mucosal microbiota of the cecum and large colon of healthy and diarrheic horses. Animals 2020, 10, 1403. [Google Scholar] [CrossRef]
- Costa, M.C.; Arroyo, L.G.; Allen-Vercoe, E.; Stämpfli, H.R.; Kim, P.T.; Sturgeon, A.; Weese, J.S. Comparison of the fecal microbiota of healthy horses and horses with colitis by high throughput sequencing of the V3-V5 region of the 16S RRNA gene. PLoS ONE 2012, 7, e41484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclere, M.; Costa, M.C. Fecal microbiota in horses with asthma. J. Vet. Intern. Med. 2020, 34, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin de Bustamante, M.; Gomez, D.; MacNicol, J.; Hamor, R.; Plummer, C. The fecal bacterial microbiota in horses with equine recurrent uveitis. Animals 2021, 11, 745. [Google Scholar] [CrossRef] [PubMed]
- Martin de Bustamante, M.; Plummer, C.; MacNicol, J.; Gomez, D. Impact of ambient temperature sample storage on the equine fecal microbiota. Animals 2021, 11, 819. [Google Scholar] [CrossRef] [PubMed]
- Langner, K.; Blaue, D.; Schedlbauer, C.; Starzonek, J.; Julliand, V.; Vervuert, I. Changes in the faecal microbiota of horses and ponies during a two-year body weight gain programme. PLoS ONE 2020, 15, e0230015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, K.A.; Rogers, C.W.; Gee, E.K.; Kittelmann, S.; Bolwell, C.F.; Bermingham, E.N.; Biggs, P.J.; Thomas, D.G. Resilience of faecal microbiota in stabled thoroughbred horses following abrupt dietary transition between freshly cut pasture and three forage-based diets. Animals 2021, 11, 2611. [Google Scholar] [CrossRef]
- Kunz, I.G.Z.; Reed, K.J.; Metcalf, J.L.; Hassel, D.M.; Coleman, R.J.; Hess, T.M.; Coleman, S.J. Equine fecal microbiota changes associated with anthelmintic administration. J. Equine Vet. Sci. 2019, 77, 98–106. [Google Scholar] [CrossRef]
- Garber, A.; Hastie, P.; McGuinness, D.; Malarange, P.; Murray, J.-A. Abrupt dietary changes between grass and hay alter faecal microbiota of ponies. PLoS ONE 2020, 15, e0237869. [Google Scholar] [CrossRef]
- Arnold, C.E.; Pilla, R.; Chaffin, M.K.; Leatherwood, J.L.; Wickersham, T.A.; Callaway, T.R.; Lawhon, S.D.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. The effects of signalment, diet, geographic location, season, and colitis associated with antimicrobial use or salmonella infection on the fecal microbiome of horses. J. Vet. Intern. Med. 2021, 35, 2437–2448. [Google Scholar] [CrossRef]
- Siddiqui, M.T.; Cresci, G.A.M. The immunomodulatory functions of butyrate. J. Inflamm. Res. 2021, 14, 6025–6041. [Google Scholar] [CrossRef]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.; Proudman, C.J.; Duncan, S.H.; Flint, H.J.; Dyer, J.; Shirazi-Beechey, S.P. Alterations in microbiota and fermentation products in equine large intestine in response to dietary variation and intestinal disease. Br. J. Nutr. 2012, 107, 989–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milinovich, G.J.; Klieve, A.V.; Pollitt, C.C.; Trott, D.J. Microbial events in the hindgut during carbohydrate-induced equine laminitis. Vet. Clin. N. Am. Equine Pract. 2010, 26, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Milinovich, G.J.; Burrell, P.C.; Pollitt, C.C.; Klieve, A.V.; Blackall, L.L.; Ouwerkerk, D.; Woodland, E.; Trott, D.J. Microbial ecology of the equine hindgut during oligofructose-induced laminitis. ISME J. 2008, 2, 1089–1100. [Google Scholar] [CrossRef] [Green Version]
- Bailey, S.R.; Menzies-Gow, N.J.; Marr, C.M.; Elliott, J. The effects of vasoactive amines found in the equine hindgut on digital blood flow in the normal horse. Equine Vet. J. 2004, 36, 267–272. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Umesaki, Y.; Setoyama, H.; Matsumoto, S.; Imaoka, A.; Itoh, K. Differential roles of segmented filamentous bacteria and clostridia in development of the intestinal immune system. Infect. Immun. 1999, 67, 3504–3511. [Google Scholar] [CrossRef] [Green Version]
- Gomez, D.E.; Arroyo, L.G.; Costa, M.C.; Viel, L.; Weese, J.S. Characterization of the fecal bacterial microbiota of healthy and diarrheic dairy calves. J. Vet. Intern. Med. 2017, 31, 928–939. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The controversial role of human gut lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Uzal, F.A.; Arroyo, L.G.; Navarro, M.A.; Gomez, D.E.; Asín, J.; Henderson, E. Bacterial and viral enterocolitis in horses: A review. J. Vet. Diagn. Investig. 2021, 10406387211057468. [Google Scholar] [CrossRef]
- Baker, J.R.; Leyland, A. Diarrhoea in the horse associated with stress and tetracycline therapy. Vet. Rec. 1973, 93, 583–584. [Google Scholar] [CrossRef] [PubMed]
- Cook, W. Diarrhoea in the horse associated with stress and tetracycline therapy. Vet. Rec. 1973, 93, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Andersson, G.; Ekman, L.; Månsson, I.; Persson, S.; Rubarth, S.; Tufvesson, G. Lethal complications following administration of oxytetracycline in the horse. Nord. Vet. 1971, 23, 9–22. [Google Scholar]
- White, G.; Prior, S.D. Comparative effects of oral administration of trimethoprim/sulphadiazine or oxytetracycline on the faecal flora of horses. Vet. Rec. 1982, 111, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Bihan, M.; Methé, B.A. Analyses of the stability and core taxonomic memberships of the human microbiome. PLoS ONE 2013, 8, e63139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shreffler, J.; Huecker, M.R. Type I and Type II Errors and Statistical Power; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Kelly, B.J.; Gross, R.; Bittinger, K.; Sherrill-Mix, S.; Lewis, J.D.; Collman, R.G.; Bushman, F.D.; Li, H. Power and sample-size estimation for microbiome studies using pairwise distances and PERMANOVA. Bioinformatics 2015, 31, 2461–2468. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liepman, R.S.; Swink, J.M.; Habing, G.G.; Boyaka, P.N.; Caddey, B.; Costa, M.; Gomez, D.E.; Toribio, R.E. Effects of Intravenous Antimicrobial Drugs on the Equine Fecal Microbiome. Animals 2022, 12, 1013. https://doi.org/10.3390/ani12081013
Liepman RS, Swink JM, Habing GG, Boyaka PN, Caddey B, Costa M, Gomez DE, Toribio RE. Effects of Intravenous Antimicrobial Drugs on the Equine Fecal Microbiome. Animals. 2022; 12(8):1013. https://doi.org/10.3390/ani12081013
Chicago/Turabian StyleLiepman, Rachel S., Jacob M. Swink, Greg G. Habing, Prosper N. Boyaka, Benjamin Caddey, Marcio Costa, Diego E. Gomez, and Ramiro E. Toribio. 2022. "Effects of Intravenous Antimicrobial Drugs on the Equine Fecal Microbiome" Animals 12, no. 8: 1013. https://doi.org/10.3390/ani12081013
APA StyleLiepman, R. S., Swink, J. M., Habing, G. G., Boyaka, P. N., Caddey, B., Costa, M., Gomez, D. E., & Toribio, R. E. (2022). Effects of Intravenous Antimicrobial Drugs on the Equine Fecal Microbiome. Animals, 12(8), 1013. https://doi.org/10.3390/ani12081013