
Effect of CXCL17 on Subcutaneous Preadipocytes Proliferation in Goats

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Animals and Samples Collection
2.2. Total RNA Extraction and Reverse Transcription
2.3. Cloning of Coding Region of Goat CXCL17
2.4. Bioinformatics Analysis of Goat CXCL17 Coding Region
2.5. Induced Differentiation of Subcutaneous Preadipocytes
2.6. Real-Time Fluorescence Quantitative PCR (qPCR)
2.7. Construction of Overexpression Plasmid and Synthesis of siRNA
2.8. Transfection
2.9. MTT Assay
2.10. Statistical Analysis
3. Results
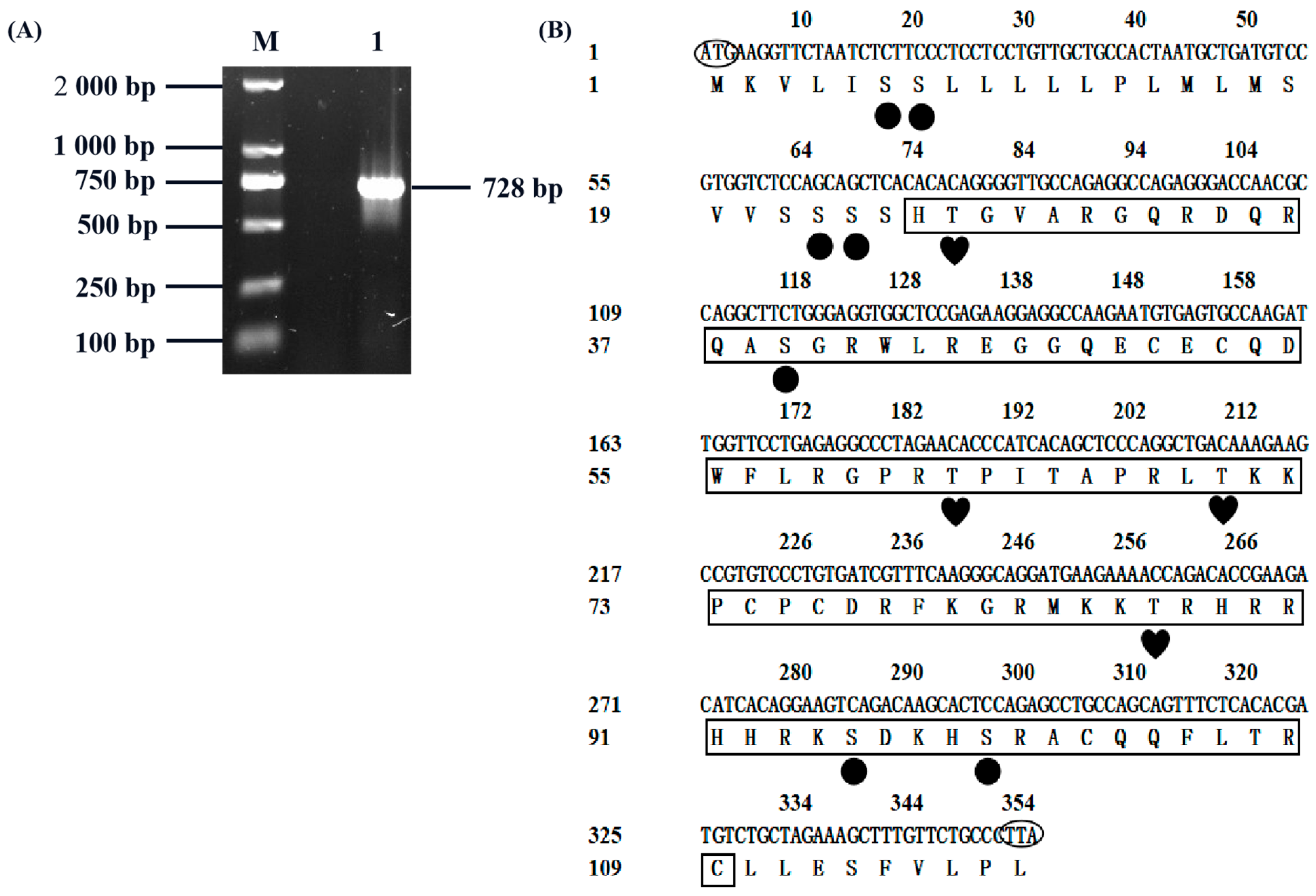
3.1. Goat CXCL17 Gene Cloning
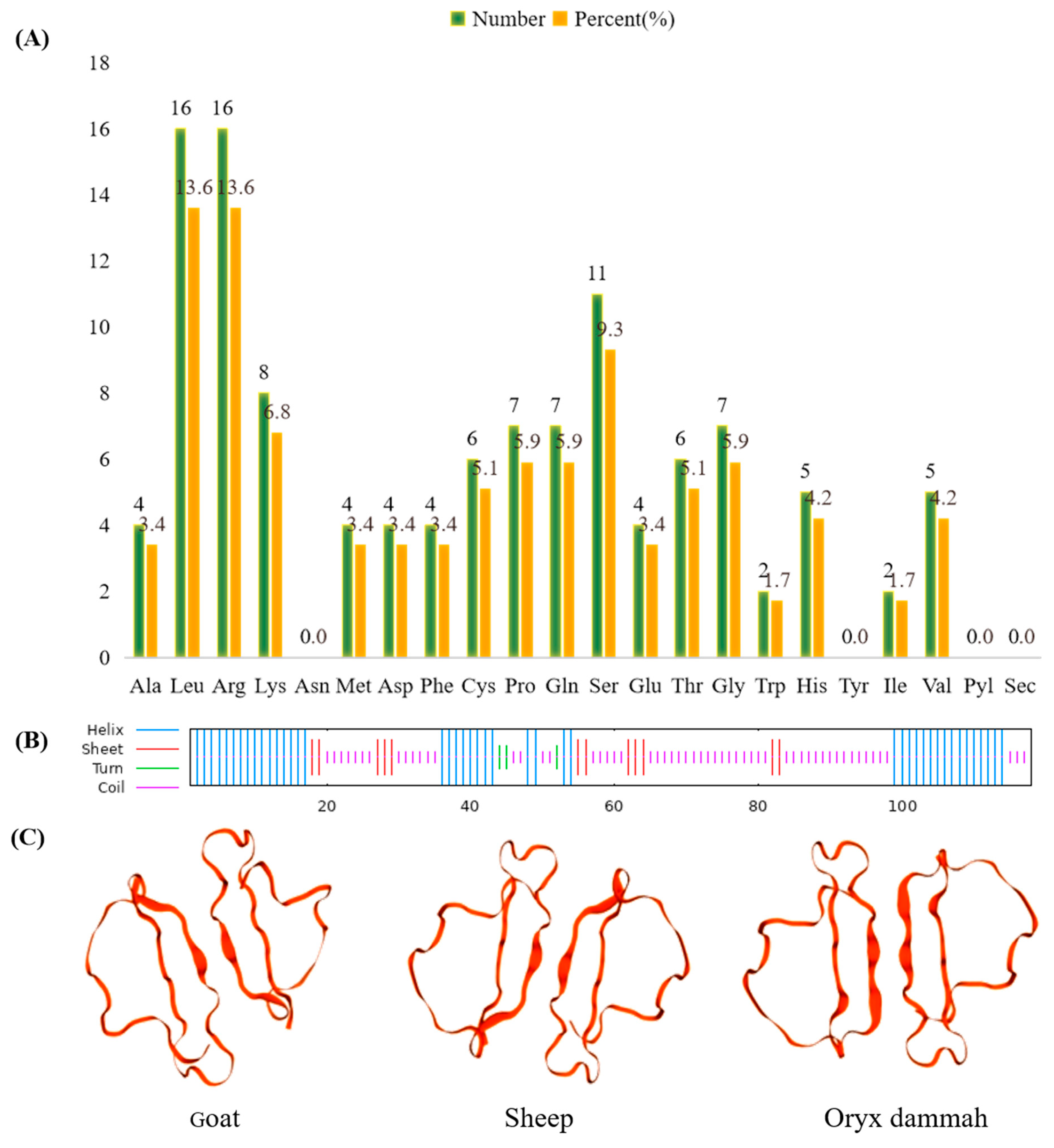
3.2. Protein Structure and Amino Acid Composition Analysis of Goat CXCL17
3.3. Subcellular Localization, Signal Peptide Prediction, Protein Phosphorylation Site and Transmembrane Domain, Amino Acid Sequence Homology and Phylogenetic Tree Analysis
3.4. Analysis of Tissue and Temporal Expression Profile of CXCL17 Gene in Goats
3.5. CXCL17 Expression Plasmid Construction
3.6. Transfected Plasmid with Different Dose Showed Different Value-Added Effect
3.7. Knockdown of CXCL17 Inhibits Goat Subcutaneous Preadipocytes Proliferation
3.8. Analysis of CXCL17 Interaction Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yoon, H.; Shaw, J.L.; Haigis, M.C.; Greka, A. Lipid metabolism in sickness and in health: Emerging regulators of lipotoxicity. Mol. Cell 2021, 81, 3708–3730. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.A.; Su, W.; Chapman, N.M.; Chi, H. Lipid metabolism in T cell signaling and function. Nat. Chem. Biol. 2022, 18, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Oteng, A.-B.; Kersten, S. Mechanisms of Action of trans Fatty Acids. Adv. Nutr. Int. Rev. J. 2020, 11, 697–708. [Google Scholar] [CrossRef]
- Li, X.; Fu, X.; Yang, G.; Du, M. Review: Enhancing intramuscular fat development via targeting fibro-adipogenic progenitor cells in meat animals. Animal 2020, 14, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Kou, J. Study on the Developmental Difference of Subcutaneous Adipose Tissue in Different Parts of Duck. Master’s Thesis, Sichuan Agricultural University, Chengdu, China, 2013. [Google Scholar]
- Anderson, F.; Pannier, L.; Pethick, D.; Gardner, G. Intramuscular fat in lamb muscle and the impact of selection for improved carcass lean meat yield. Animal 2015, 9, 1081–1090. [Google Scholar] [CrossRef]
- Liu, R.; Liu, X.; Bai, X.; Xiao, C.; Dong, Y. A Study of the Regulatory Mechanism of the CB1/PPARγ2/PLIN1/HSL Pathway for Fat Metabolism in Cattle. Front. Genet. 2021, 12, 631187. [Google Scholar] [CrossRef]
- Denisov, S.S. CXCL17: The Black Sheep in the Chemokine Flock. Front. Immunol. 2021, 12, 712897. [Google Scholar] [CrossRef]
- Guo, Y.J.; Ou, Z.L.; Shao, Z.M. CXCL17: A novel member of the CXC chemokine family. Chem. Life 2013, 33, 5. [Google Scholar] [CrossRef]
- Lee, W.-Y.; Wang, C.-J.; Lin, T.-Y.; Hsiao, C.-L.; Luo, C.-W. CXCL17, an orphan chemokine, acts as a novel angiogenic and anti-inflammatory factor. Am. J. Physiol. Metab. 2013, 304, E32–E40. [Google Scholar] [CrossRef]
- Xiao, S.; Xie, W.; Zhou, L. Mucosal chemokine CXCL17: What is known and not known. Scand. J. Immunol. 2020, 93, e12965. [Google Scholar] [CrossRef]
- Oka, T.; Sugaya, M.; Takahashi, N.; Takahashi, T.; Shibata, S.; Miyagaki, T.; Asano, Y.; Sato, S. CXCL17 Attenuates Imiquimod-Induced Psoriasis-like Skin Inflammation by Recruiting Myeloid-Derived Suppressor Cells and Regulatory T Cells. J. Immunol. 2017, 198, 3897–3908. [Google Scholar] [CrossRef]
- Hsu, Y.-L.; Yen, M.-C.; Chang, W.-A.; Tsai, P.-H.; Pan, Y.-C.; Liao, S.-H.; Kuo, P.-L. CXCL17-derived CD11b+Gr-1+ myeloid-derived suppressor cells contribute to lung metastasis of breast cancer through platelet-derived growth factor-BB. Breast Cancer Res. 2019, 21, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, H.; Zhen, Z.; Ma, X.; Yu, W.; Zeng, H.; Li, L. CXCL17 promotes cell metastasis and inhibits autophagy via the LKB1-AMPK pathway in hepatocellular carcinoma. Gene 2018, 690, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, L.; Hammarström, M.-L.; Lindmark, G.; Hammarström, S.; Sitohy, B. Ectopic expression of the chemokine CXCL17 in colon cancer cells. Br. J. Cancer 2016, 114, 697–703. [Google Scholar] [CrossRef]
- Yu, J.L.; Ruan, J.Y.; Li, S.H.; He, B.J.; Zhu, P.Y.; Xue, X.Y.; Zou, R.M.; Lin, K.Z. Expression of chemokine CXCL17 and its clinical relevance in gastric carcinoma. Chin. J. Pathophysiol. 2016, 32, 1022–1026. [Google Scholar]
- Liu, W.; Xie, X.; Wu, J. Mechanism of lung adenocarcinoma spine metastasis induced by CXCL17. Cell Oncol. 2020, 43, 311–320. [Google Scholar] [CrossRef]
- Hiraoka, N.; Yamazaki–Itoh, R.; Ino, Y.; Mizuguchi, Y.; Yamada, T.; Hirohashi, S.; Kanai, Y. CXCL17 and ICAM2 Are Associated with a Potential Anti-Tumor Immune Response in Early Intraepithelial Stages of Human Pancreatic Carcinogenesis. Gastroenterology 2011, 140, 310–321.e4. [Google Scholar] [CrossRef]
- Matsui, A.; Yokoo, H.; Negishi, Y.; Endo-Takahashi, Y.; Chun, N.A.L.; Kadouchi, I.; Suzuki, R.; Maruyama, K.; Aramaki, Y.; Semba, K.; et al. CXCL17 Expression by Tumor Cells Recruits CD11b+Gr1highF4/80− Cells and Promotes Tumor Progression. PLoS ONE 2012, 7, e44080. [Google Scholar] [CrossRef]
- Burkhardt, A.M.; Tai, K.P.; Flores-Guiterrez, J.P.; Vilches-Cisneros, N.; Kamdar, K.; Barbosa-Quintana, O.; Valle-Rios, R.; Hevezi, P.A.; Zuñiga, J.; Selman, M.; et al. CXCL17 Is a Mucosal Chemokine Elevated in Idiopathic Pulmonary Fibrosis That Exhibits Broad Antimicrobial Activity. J. Immunol. 2012, 188, 6399–6406. [Google Scholar] [CrossRef]
- Shimada, S.; Makino, K.; Jinnin, M.; Sawamura, S.; Kawano, Y.; Ide, M.; Kajihara, I.; Makino, T.; Fukushima, S.; Ihn, H. CXCL17-mediated downregulation of type I collagen via MMP1 and miR-29 in skin fibroblasts possibly contributes to the fibrosis in systemic sclerosis. J. Dermatol. Sci. 2020, 100, 183–191. [Google Scholar] [CrossRef]
- Srivastava, R.; Hernández-Ruiz, M.; Khan, A.A.; Fouladi, M.A.; Kim, G.J.; Ly, V.T.; Yamada, T.; Lam, C.; Sarain, S.A.B.; Boldbaatar, U.; et al. CXCL17 Chemokine–Dependent Mobilization of CXCR8+CD8+ Effector Memory and Tissue-Resident Memory T Cells in the Vaginal Mucosa Is Associated with Protection against Genital Herpes. J. Immunol. 2018, 200, 2915–2926. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ruiz, M.; Zlotnik, A.; Llorente, L.; Hernandez-Molina, G. Markedly high salivary and lacrimal CXCL17 levels in primary Sjögren’s syndrome. Jt. Bone Spine 2018, 85, 379–380. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.; DelCurto-Wyffels, H.; Thomson, J.; Boles, J. Fat Deposition and Fat Effects on Meat Quality—A Review. Animals 2022, 12, 1550. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.L.; Li, Y.Y.; Lin, Y.Q.; Chen, D.; Sheng, X.; Zhao, N.; Liu, W. Cloning and expression characteristic analysis of goat ST13 gene. Chin. J. Biotech. 2022, 38, 2959–2973. [Google Scholar] [CrossRef]
- Sheng, X.Q.; Zhao, N.; Lin, Y.Q.; Chen, D.S.; Wang, R.L.; Li, Y.Y. Cloning and expression analysis of goat ZNF32. Biotechnol. Bull. 2022, 38, 300–311. [Google Scholar] [CrossRef]
- Lin, S.; Lin, Y.Q.; Zhu, J.J.; Xu, Q.; Zhao, Y.; Ji, Y.D.; Wang, Y. Effect of Wnt10b on the expression of preadipocytes differentiation related genes in goat. Acta Vet. Zootech. Sin. 2018, 49, 685–692. [Google Scholar]
- Chen, D.; Wang, R.; Sheng, X.; Zhao, N.; Lin, Y.; Wang, Y.; Zhu, J.; Li, Y. PDZK1-interacting protein 1(PDZK1IP1) promotes subcutaneous preadipocyte proliferation in goats. Anim. Biotechnol. 2022, 1–11. [Google Scholar] [CrossRef]
- Chi, Y.D.; Wang, Y.; Hu, M. Screening of Internal Reference Genes in Different Tissues and Organs of Goats. Genom. Appl. Biol. 2020, 39, 561–567. [Google Scholar] [CrossRef]
- Lin, L.-C.; Quon, T.; Engberg, S.; Mackenzie, A.E.; Tobin, A.B.; Milligan, G. G Protein-Coupled Receptor GPR35 Suppresses Lipid Accumulation in Hepatocytes. ACS Pharmacol. Transl. Sci. 2021, 4, 1835–1848. [Google Scholar] [CrossRef]
- Maravillas-Montero, J.L.; Burkhardt, A.M.; Hevezi, P.A.; Carnevale, C.D.; Smit, M.J.; Zlotnik, A. Cutting Edge: GPR35/CXCR8 Is the Receptor of the Mucosal Chemokine CXCL17. J. Immunol. 2015, 194, 29–33. [Google Scholar] [CrossRef]
- Hernández-Ruiz, M.; Othy, S.; Herrera, C.; Nguyen, H.-T.; Arrevillaga-Boni, G.; Catalan-Dibene, J.; Cahalan, M.D.; Zlotnik, A. Cxcl17 -/- mice develop exacerbated disease in a T cell-dependent autoimmune model. J. Leukoc. Biol. 2019, 105, 1027–1039. [Google Scholar] [CrossRef]
- Choreño-Parra, J.A.; Thirunavukkarasu, S.; Zúñiga, J.; Khader, S.A. The protective and pathogenic roles of CXCL17 in human health and disease: Potential in respiratory medicine. Cytokine Growth Factor Rev. 2020, 53, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Choreño-Parra, J.A.; Jiménez-Álvarez, L.A.; Ramírez-Martínez, G.; Sandoval-Vega, M.; Salinas-Lara, C.; Sánchez-Garibay, C.; Luna-Rivero, C.; Hernández-Montiel, E.M.; Fernández-López, L.A.; Cabrera-Cornejo, M.F.; et al. CXCL17 Is a Specific Diagnostic Biomarker for Severe Pandemic Influenza A(H1N1) That Predicts Poor Clinical Outcome. Front. Immunol. 2021, 12, 633297. [Google Scholar] [CrossRef]
- Zhang, K.; Liang, Y.; Feng, Y.; Wu, W.; Zhang, H.; He, J.; Hu, Q.; Zhao, J.; Xu, Y.; Liu, Z.; et al. Decreased epithelial and sputum miR-221-3p associates with airway eosinophilic inflammation and CXCL17 expression in asthma. Am. J. Physiol. Cell Mol. Physiol. 2018, 315, L253–L264. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Kojder, K.; Kapczuk, P.; Kupnicka, P.; Gawrońska-Szklarz, B.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. The Effect of Hypoxia on the Expression of CXC Chemokines and CXC Chemokine Receptors—A Review of Literature. Int. J. Mol. Sci. 2021, 22, 843. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Z. The Involvement of CXCL17-GPR35 in the Stepwise Process of Gastic Cancer Development and Its Related Finction Analysis. Master’s Thesis, China Medical University, Taiwan, China, 2021. [Google Scholar] [CrossRef]
- Zhang, Z.S. The Correlation between Chemokine CXCL17 and Proliferation and Proliferation of Gastric Cancer Cells. Master’s Thesis, Zhejiang University, Hangzhou, China, 2019. [Google Scholar] [CrossRef]
- Li, S.Z.; Xiao, S.Y.; Xue, Y.; Zhou, L.Y. Characterization of gastric microbiota in patients eradicated Helicobacter pylori and its relationship with CXCL17. Chin. J. Gastroenterol. Hepatol. 2021, 30, 883–892. [Google Scholar]
- Li, L.; Yan, J.; Xu, J.; Liu, C.-Q.; Zhen, Z.-J.; Chen, H.-W.; Ji, Y.; Wu, Z.-P.; Hu, J.-Y.; Zheng, L.; et al. CXCL17 Expression Predicts Poor Prognosis and Correlates with Adverse Immune Infiltration in Hepatocellular Carcinoma. PLoS ONE 2014, 9, e110064. [Google Scholar] [CrossRef]
- Kochumon, S.; Al Madhoun, A.; Al-Rashed, F.; Azim, R.; Al-Ozairi, E.; Al-Mulla, F.; Ahmad, R. Adipose tissue gene expression of CXCL10 and CXCL11 modulates inflammatory markers in obesity: Implications for metabolic inflammation and insulin resistance. Ther. Adv. Endocrinol. Metab. 2020, 11, 902. [Google Scholar] [CrossRef]
- Ma, F.F.; Cao, D.D.; Li, W.; Tang, R.Q.; Ouyang, S.R.; Wu, J.X. Expression and significance of chemokine CXCL1 in the adipose tissue of mice with obesity. J. Xinxiang Med. Univ. 2016, 33, 849–851. [Google Scholar]
- Pedersen, L.; Hojman, P. Muscle-to-organ cross talk mediated by myokines. Adipocyte 2012, 1, 164–167. [Google Scholar] [CrossRef]
- Hou, L.N.; Liu, J.; Li, Y.L.; Sun, Z.; Zhang, L.L.; Wang, Z.Q. Effect of C-X-C motif chemokine ligand 14 on adipocytes pyroptosis in high glucose environment. Med. Sci. J. Cent. South China 2022, 50, 7–12. [Google Scholar] [CrossRef]
- Pisabarro, M.T.; Leung, B.; Kwong, M.; Corpuz, R.; Frantz, G.D.; Chiang, N.; Vandlen, R.; Diehl, L.J.; Skelton, N.; Kim, H.S.; et al. Cutting Edge: Novel Human Dendritic Cell- and Monocyte-Attracting Chemokine-Like Protein Identified by Fold Recognition Methods. J. Immunol. 2006, 176, 2069–2073. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytical Contents | Analytical Software or Online Tools |
|---|---|
| Nucleotide sequence alignment | DNAMAN |
| Amino acid sequence translation | ORF Finder (NCBI) |
| Prediction of phosphorylation sites | NetPhos 3.1 |
| Prediction of signal peptide sites | SignalP-5.0 Server |
| Prediction of transmembrane domain | TMHMM |
| Subcellular localization analysis | PSORT Ⅱ |
| Prediction of secondary structure protein | SOPMA |
| Prediction of protein tertiary structure | SWISS-MODEL |
| Protein–protein interaction analysis | STRING |
| Phylogenetic tree construction | MEGA 5.05 |
| Conserved Domain analysis | Conserved Domain (NCBI) |
| Physical and chemical properties and primary structure analysis | Ex PASy-ProParam |
| Primer Names | Primer Sequence (5′→3′) * | Product Length (bp) | Tm (T/°C) | Purpose |
|---|---|---|---|---|
| CXCL17 | F: 5′ CCTGTTGCTGCCACTAATGC 3′ R: 5′ GTGATGTCTTCGGTGTCTGGT 3′ | 250 | 60 | qPCR |
| CXCL17 | F: 5′ CTAGAATTCTGATGAAGGTTCTAATCTCTTCCC 3′ 1 R: 5′ CGGGGTACCACTAAGGGCAGAACAAAGCTT 3′ | 357 | 60 | plasmid construction |
| UXT | F: 5′ GCAAGTGGATTTGGGCTGTAAC 3′ R: 5′ ATGGAGTCCTTGGTGAGGTTGT 3′ | 180 | 60 | qPCR |
| CCNE1 | F: 5′ CTCCCTGATTCCCACACCTG 3′ R: 5′ CATAAGATGCTTGTCCCTCA3′ | 193 | 60 | qPCR |
| PCNA | F: 5′ AGTGGAGAACTTGGAAATGGAA 3′ R: 5′ GAGACAGTGGAGTGGCTTTTGT 3′ | 154 | 60 | qPCR |
| CCND1 | F: 5′ TGAACTACCTGGACCGCT 3′ R: 5′ CAGGTTCCACTTGAGTTTGT 3′ | 212 | 60 | qPCR |
| CDK2 | F: 5′ GCCAGGAGTTACTTCTATGC 3′ R: 5′ TGGAAGAAAGGGTGAGCC 3′ | 180 | 60 | qPCR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, G.; Ma, X.; Wang, F.; Chen, D.; Lin, Y.; Wang, Y.; Liu, W.; Li, Y. Effect of CXCL17 on Subcutaneous Preadipocytes Proliferation in Goats. Animals 2023, 13, 1757. https://doi.org/10.3390/ani13111757
Lu G, Ma X, Wang F, Chen D, Lin Y, Wang Y, Liu W, Li Y. Effect of CXCL17 on Subcutaneous Preadipocytes Proliferation in Goats. Animals. 2023; 13(11):1757. https://doi.org/10.3390/ani13111757
Chicago/Turabian StyleLu, Guangyu, Xiaotong Ma, Fei Wang, Dingshuang Chen, Yaqiu Lin, Youli Wang, Wei Liu, and Yanyan Li. 2023. "Effect of CXCL17 on Subcutaneous Preadipocytes Proliferation in Goats" Animals 13, no. 11: 1757. https://doi.org/10.3390/ani13111757
APA StyleLu, G., Ma, X., Wang, F., Chen, D., Lin, Y., Wang, Y., Liu, W., & Li, Y. (2023). Effect of CXCL17 on Subcutaneous Preadipocytes Proliferation in Goats. Animals, 13(11), 1757. https://doi.org/10.3390/ani13111757