
Circadian Variation of Peripheral Blood Cells in Horses Maintained in Different Environmental and Management Conditions
 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Hematological Analysis
2.3. Statistical Analysis
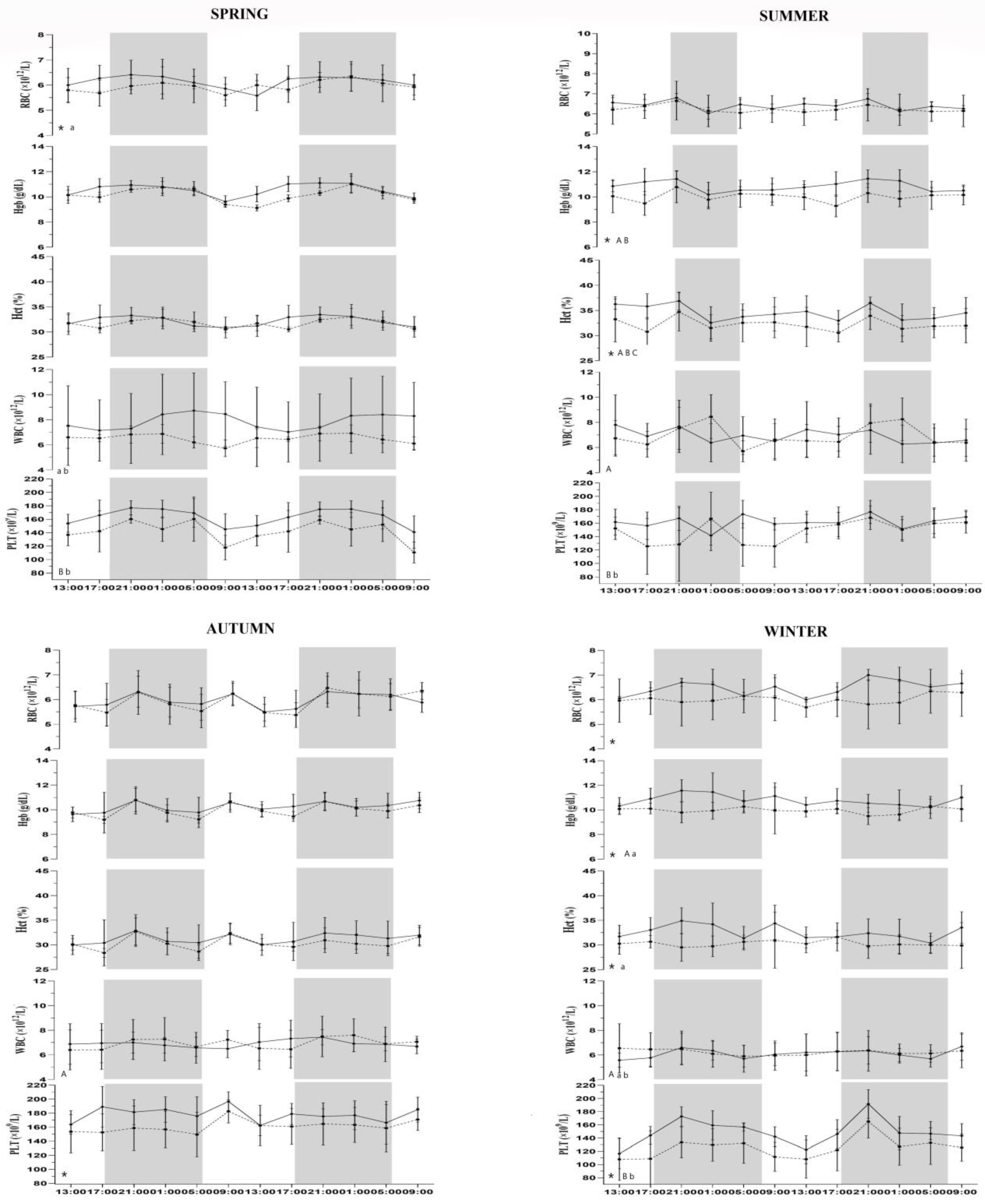
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huber, N.; Marasco, V.; Painer, J.; Vetter, S.G.; Göritz, F.; Kaczensky, P.; Walzer, C. Leukocyte Coping Capacity: An Integrative Parameter for Wildlife Welfare within Conservation Interventions. Front. Vet. Sci. 2019, 6, 105. [Google Scholar] [CrossRef] [Green Version]
- Sinha, R.; Kamboj, M.L.; Ranjan, A.; Devi, I. Effect of microclimatic variables on physiological and hematological parameters of crossbred cows in summer season. Indian J. Anim. Res. 2019, 53, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Nelson, R.J. Seasonal immune function and sickness responses. Trends Immunol. 2004, 25, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Piccione, G.; Caola, G.; Refinetti, R. Daily rhythms of blood pressure, heart rate, and body temperature in fed and fasted male dogs. J. Vet. Med. A 2005, 52, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Giannetto, C.; Aragona, F.; Arfuso, F.; Piccione, G.; De Caro, S.; Fazio, F. Diurnal variation in rectal and cutaneous temperatures of horses housed under diferent management conditions. Int. J. Biometeolol. 2022, 66, 1601–1611. [Google Scholar] [CrossRef]
- Piccione, G.; Fazio, F.; Giudice, E.; Grasso, F.; Morgante, M. Nycthemeral change of some haematological parameters in horses. J. Appl. Biomed. 2005, 3, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Dunlap, J.C.; Loros, J.J.; DeCoursey, P.J. Biological Timekeeping, Chronobiol; Sinauer Associates: Sunderland, UK, 2004; pp. 67–106. [Google Scholar]
- Giudice, E.; Messina, V.; Marafioti, S.; Monteverde, V.; Bazzano, M.; Piccione, G.; Guercio, A. Influence of Brucella infection on circadian rhythm of haematological parameters in dairy cows. Large Anim. Rev. 2012, 18, 75–77. [Google Scholar]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef]
- Satué, K.; Muñoz, A.; Montesinos, P. Seasonal variations in the erythrogram in pregnant Carthusian mares. In Proceedings of the 13th Conference of the ESVCP/ECVCP, 9TH Conference of AECCP, 12TH ACCP and ASVCP, Dublin, Ireland, 31 August–3 September 2011. [Google Scholar]
- Satué, K.; Hernández, A.; Muñoz, A. Physiological factors in the interpretation of equine hematological profile. In Hematology—Science and Practice; Lawrie, C., Ed.; InTech: Vienna, Austria, 2012. [Google Scholar]
- Satué, K.; Gardón, J.C.; Muňoz, A. Season of the year should be considered in the interpretation of hematology in carthusian. J. Hematol. Res. 2014, 1, 63–68. [Google Scholar] [CrossRef]
- Andriichuk, A.; Tkachenko, H. Seasonal variations of hematological indices in equines involved in recreational horse riding. Balt. Coast. Zone J. Ecol. Prot. Coastline 2015, 19, 11–22. [Google Scholar]
- Bazzano, M.; Giannetto, C.; Fazio, F.; Rizzo, M.; Giudice, E.; Piccione, G. Physiological adjustments of haematological profile during the last trimester of pregnancy and the early post-partum period in mares. Anim. Reprod. Sci. 2014, 149, 199–203. [Google Scholar] [CrossRef]
- Kumar, B.; Pachauri, S.P. Haematological profile of crossbred dairy cattle to monitor herd health status at medium elevation in Central Himalayas. Res. Vet. Sci. 2000, 69, 141–145. [Google Scholar] [CrossRef]
- Haus, E.; Smolensky, M.H. Biologic rhythms in the immune system. Chronobiol. Int. 1999, 16, 581–622. [Google Scholar] [CrossRef] [PubMed]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P.S. Circadian control of the immune system. Nat. Rev. Immunol. 2013, 13, 190–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Huang, W.; Ye, B.; Chen, C.; Huang, R.; Wu, F.; Wei, Q.; Zhang, W.; Hu, J. Changes of hematological and immunological parameters in COVID-19 patients. Int. J. Hematol. 2020, 112, 553–559. [Google Scholar] [CrossRef]
- Orozco-Solis, R.; Aguilar-Arnal, L. Circadian Regulation of Immunity Through Epigenetic Mechanisms. Front. Cell. Infect. Microbiol. 2020, 10, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downtown, P.; Early, J.O.; Gibbs, J.E. Circadian rhythm in adaptive immunity. Immunology 2019, 161, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Pritchett, D.; Reddy, A.B. Circadian Clocks in the Hematologic System. J. Biol. Rhythm. 2015, 30, 374–388. [Google Scholar] [CrossRef]
- Feriel, J.; Tchipeva, D.; Depasse, F. Effects of circadian variation, lifestyle and environment on hematological parameters: A narrative review. Int. J. Lab. Hematol. 2021, 43, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Pelegri, C.; Vilaplana, J.; Castellote, C.; Rabanal, M.; Franch, A.; Castell, M. Circadian rhythms in surface molecules of rat blood lymphocytes. Am. J. Physiol. Cell Physiol. 2003, 284, C67–C76. [Google Scholar] [CrossRef] [Green Version]
- Piccione, G.; Assenza, A.; Fazio, F.; Giudice, E.; Caola, G. Different periodicities of some haematological parameters in exercise- loaded athletic horses and sedentary horses. J. Equine Sci. 2001, 12, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Arfuso, F.; Giudice, E.; Di Pietro, S.; Piccione, G.; Giannetto, C. Modulation of serum protein electrophoretic pattern and leukocyte population in horses vaccinated against West Nile virus. Animals 2021, 11, 477. [Google Scholar] [CrossRef]
- Giannetto, C.; Giudice, E.; Piccione, G.; Castronovo, C.; Arfuso, F. Immune and Inflammatory Response in Horse Vaccinated Against Equine Herpesviruses 1 (EHV-1) and 4 (EHV-4) Assessed by Serum Protein Electrophoretic Pattern and Leukocyte Population. J. Equine Vet. Sci. 2022, 116, 104051. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, E.; Hopkins, R.J.; Blomgren, E.; Ventrop, M.; Brömssen, C.; Dahlborn, K. Daytime shelter use of individually kept horses during Swedish summer. J. Anim. Sci. 2015, 93, 802–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Refinetti, R.; Piccione, G. Intra- and inter-individual variability in the circadian rhythm of body temperature of rats, squirrels, dogs, and horses. J. Therm. Biol. 2005, 30, 139–146. [Google Scholar] [CrossRef]
- Nelson, W.; Tong, Y.L.; Lee, J.K.; Halberg, F. Methods for cosinor-rhythmometry. Chronobiologia 1979, 6, 305–323. [Google Scholar] [PubMed]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Clinical Biochemistry of Domestic Animals, 6th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Zakari, F.O.; Ayo, J.O.; Rekwot, P.I.; Kawu, M.U. Effects of age and season on haematological parameters of donkeys during the rainy and cold–dry seasons. Int. J. Biometeorol. 2015, 59, 1813–1824. [Google Scholar] [CrossRef]
- Gill, J.; Kownacka, M. Seasonal changes in erythrocyte, hemoglobin and leukocyte indexes in pregnant mares of thoroughbred horses. Bull. Pol. Acad. Sci. Tech. Sci. 1979, 27, 143–148. [Google Scholar]
- Haus, E. Chronobiology of circulating blood cells and platelets. In Biological Rhythms in Clinical and Laboratory Medicine; Touitou, Y., Haus, E., Eds.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 504–526. [Google Scholar]
- Mirzadeh, H.; Najafizadeh, A. Flow stress prediction at hot working conditions. Mater. Sci. Eng. A 2010, 527, 1160–1164. [Google Scholar] [CrossRef]
- Davi, G.; Patrono, C. Platelet activation and atherothrombosis. N. Engl. J. Med. 2007, 357, 2482–2494. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K. Thermoneutral zone and critical temperatures of horses. J. Therm. Biol. 1998, 23, 59–61. [Google Scholar] [CrossRef]
- Ono, T.; Inoue, Y.; Hisaeda, K.; Yamada, Y.; Hata, A.; Miyama, T.S.; Shibano, K.; Kitagawa, H.; Ohzawa, E.; Iwata, E. Effect of seasons and sex on the physical, hematological, and blood biochemical parameters of Noma horses. J. Equine Sci. 2021, 32, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Widenhorn, N.I.; Scaglione, M.C.; Tomatis, F.M.; Sartore, A.L. Endogenous haematological rhythm in dogs. Comp. Clin. Pathol. 2011, 20, 491–499. [Google Scholar] [CrossRef]
- Castanheira, M.; Rezende Paiva, S.; Louvandini, H.; Landim, A.; Soares Fiorvanti, M.C.; Paludo, G.R.; Dallago, B.S.; McManus, C. Multivariate analysis for characteristics of heat tolerance in horses in Brazil. Trop. Anim. Health Prod. 2010, 42, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Naidenko, S.V.; Klyuchnikova, P.S.; Kirilyuk, V.E.; Alekseeva, G.S. Effect of population density on number of leukocytes in domestic cats. Nat. Conserv. Res. 2020, 5, 89–96. [Google Scholar] [CrossRef]
- Shawaf, T.; Al Mubarak, A.; Eidi, H.; El-Bahr, S.M. Season’s Effects on Some Clinical, Hematological Parameters and Blood Cortisol Level in Sedated Arabian Horses with Xylazine. J. Equine Vet. Sci. 2020, 84, 102835. [Google Scholar] [CrossRef]
- Berger, J. Regulation of circadian rhythms. J. Appl. Biomed. 2004, 2, 131–140. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| BOX | PADDOCK | |
|---|---|---|
| SPRING | ||
| Ambient temperature (°C) | 16.40 ± 2.26 | 16.47 ± 0.39 |
| Relative humidity (%) | 61.00 ± 0 | 44.76 ± 0 |
| Ventilation (m/s) | 0.03 ± 0 a | 0.28 ± 0.08 a |
| Temperature–humidity index (THI) | 60.74 ± 2.47 | 60.50 ± 3.32 |
| SUMMER | ||
| Ambient temperature (°C) | 25 ± 0 | 23.50 ± 0 |
| Relative humidity (%) | 63 ± 0 | 61.50 ± 0.01 |
| Ventilation (m/s) | 0.4 ± 0.01 a | 1.38 ± 0.34 a |
| Temperature–humidity index (THI) | 73.08 ± 2.79 | 70.75 ± 4.09 |
| AUTUMN | ||
| Ambient temperature (°C) | 23.98 ± 0.96 | 23.15 ± 1.88 |
| Relative humidity (%) | 75.00 ± 6.57 | 73.83 ± 6.50 |
| Ventilation (m/s) | 0.00 ± 0.00 a | 0.52 ± 0.68 a |
| Temperature–humidity index (THI) | 72.76 ± 1.31 | 71.39 ± 1.60 |
| WINTER | ||
| Ambient temperature (°C) | 16.33 ± 1.74 | 15.00 ± 1.02 |
| Relative humidity (%) | 83.00 ± 9.49 | 82.50 ± 8.19 |
| Ventilation (m/s) | 0.00 ± 0.00 a | 0.17 ± 0.14 a |
| Temperature–humidity index (THI) | 61.06 ± 1.73 | 58.89 ± 1.90 |
| Box | Paddock | |||||||
|---|---|---|---|---|---|---|---|---|
| Mesor (1012/L) | Amplitude (%) | Acrophase (hh:mm) | Robustness (%) | Mesor (1012/L) | Amplitude (%) | Acrophase (hh:mm) | Robustness (%) | |
| SPRING | ||||||||
| LYMPHOCYTES | 3.24 ± 1.30 | 0.68 ± 0.32 | 6:15 ± 0.26 | 87.05 ± 13.21 | No Rhythmicity | |||
| MONOCYTES | No Rhythmicity | |||||||
| NEUTROPHILS | 4.13 ± 1.41 | 1.05 ± 0.62 | 06:27 ± 0.33 | 77.08 ± 10.31 | ||||
| EOSINOPHILS | No Rhythmicity | |||||||
| BASOPHILES | No Rhythmicity | |||||||
| SUMMER | ||||||||
| LYMPHOCYTES | 2.99 ± 0.08 | 0.59 ± 0.21 | 18:11 ± 0.02 | 92.1 ± 7.55 | No Rhythmicity | |||
| MONOCYTES | No Rhythmicity | |||||||
| NEUTROPHILS | 2.04 ± 0.12 | 0.43 ± 0.019 | 17:50 ± 0.52 | 93.9 ± 9.12 | ||||
| EOSINOPHILS | No Rhythmicity | |||||||
| BASOPHILES | No Rhythmicity | |||||||
| AUTUMN | ||||||||
| LYMPHOCYTES | No Rhythmicity | No Rhythmicity | ||||||
| MONOCYTES | ||||||||
| NEUTROPHILS | ||||||||
| EOSINOPHILS | ||||||||
| BASOPHILES | ||||||||
| WINTER | ||||||||
| LYMPHOCYTES | No Rhythmicity | 2.98 ± 0.96 | 0.51 ± 0.31 | 20.20 ± 0.05 | 84.17 ± 15.75 | |||
| MONOCYTES | No Rhythmicity | |||||||
| NEUTROPHILS | 2.99 ± 0.76 | 0.51 ± 0.43 | 19.27 ± 0.17 | 96.8 ± 17.43 | ||||
| EOSINOPHILS | No Rhythmicity | |||||||
| BASOPHILES | No Rhythmicity | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aragona, F.; Arfuso, F.; Fazio, F.; De Caro, S.; Giudice, E.; Monteverde, V.; Piccione, G.; Giannetto, C. Circadian Variation of Peripheral Blood Cells in Horses Maintained in Different Environmental and Management Conditions. Animals 2023, 13, 1865. https://doi.org/10.3390/ani13111865
Aragona F, Arfuso F, Fazio F, De Caro S, Giudice E, Monteverde V, Piccione G, Giannetto C. Circadian Variation of Peripheral Blood Cells in Horses Maintained in Different Environmental and Management Conditions. Animals. 2023; 13(11):1865. https://doi.org/10.3390/ani13111865
Chicago/Turabian StyleAragona, Francesca, Francesca Arfuso, Francesco Fazio, Salvatore De Caro, Elisabetta Giudice, Vincenzo Monteverde, Giuseppe Piccione, and Claudia Giannetto. 2023. "Circadian Variation of Peripheral Blood Cells in Horses Maintained in Different Environmental and Management Conditions" Animals 13, no. 11: 1865. https://doi.org/10.3390/ani13111865
APA StyleAragona, F., Arfuso, F., Fazio, F., De Caro, S., Giudice, E., Monteverde, V., Piccione, G., & Giannetto, C. (2023). Circadian Variation of Peripheral Blood Cells in Horses Maintained in Different Environmental and Management Conditions. Animals, 13(11), 1865. https://doi.org/10.3390/ani13111865